环状RNA特征与功能及在水产中的研究进展
2021-11-04闫润堃刘晓林孟智超孟沛艺
李 斌 ,闫润堃,刘晓林,孟智超,李 悦,孟沛艺,黄 勇
(河南科技大学动物科技学院,河南 洛阳 471023)
环状RNA(Circluar RNAs,circRNAs)是一类不具有5′和3′末端头尾的闭合环状非编码RNA分子,广泛存在于真核细胞中[1-2]。早在1976年,在马铃薯(Solanumtuberosum)块茎类病毒中首次发现一些RNA病毒能抗磷酸二酯酶的降解,并鉴定出这些RNA分子是由一个以上外显子以共价闭环结构形式存在[3]。随后研究人员通过电子显微镜能观察到真核细胞中此RNA为环形结[4]。鉴于circRNAs结构的特殊性和当时科技条件对RNA分子世界认识的限制,circRNAs曾一度被认为是因RNA剪切拼接错误而形成且几乎无生物功能的副产物,因而未引起当时研究者的广泛重视[5]。直到1991年,Nigro等首次证实在人类细胞中鉴定出了circRNAs,其重要的生理功能才逐渐被人们所认识[6]。近年来,随着生物信息分析技术的快速发展和RNA高通量测序技术的进步,越来越多的circRNAs在真核生物中被发现,使其成为继发现微小RNA(MicroRNA,miRNA),Piwi蛋白相作用RNA(Piwi-interactiing RNA,piRNA)和长链非编码(Long non-coding RNA,lncRNA)之后RNA领域又一研究的热点[7-8]。本文综述了circRNAs的生物学特征、形成机制、主要功能及其在水产中的最新研究进展,为深入研究circRNAs调控机制提供一定的理论基础。
1 circRNAs的生物学特征
目前研究发现,circRNAs 在生物体中所具有的重要功能与它的生物学特征有关。主要特征有:1)细胞定位:大部分circRNAs来源于编码基因的外显子并位于细胞质中,少数的circRNAs来源于内含子和外显子-内含子并位于细胞核中[9];2)在生物体中广泛存在:circRNAs从病毒、真菌到高等脊椎动物以及植物等多种生物组织中都存在(图1)[10];3)高稳定性:由于circRNA是通过5′与3′端首尾相连共价闭合的环状分子,所以能抵抗RNA外切酶(Ribonuclease R,RNase R)的降解,比对应线性mRNA稳定得多[11];4)较高保守性:大部分circRNAs的一些序列在进化上是保守的,不仅在哺乳动物中具有保守性,甚至在进化关系较远的果蝇也存在保守性[12];5)半衰期较长:研究表明,circRNAs比对应的线性mRNA 转录本更具稳定性,circRNAs的半衰期超过48 h,而mRNA的半衰期约10 h[13-14];6)特异性表达:circRNA在不同细胞类型和不同组织中的发育阶段有特异性表达,尤其是在病理和非病理组织中存在明显的表达差异[15],例如由DOCK1基因产生的circRNA在乳腺癌细胞系MCF-7中高水平表达,而在肺癌细胞系A549中呈现低水平表达[16];7)差异性表达:有研究表明约1/8 的基因同时含有线性mRNAs和circRNAs,而且在有些组织中,细胞内circRNAs的含量比对应的线性mRNA更加丰富,其表达水平比线性mRNA 高出10倍之多[17-19]。

2 circRNAs的形成
经典的生物真核mRNA生成机制是mRNA前体(Precursor mRNA,pre-mRNA)剪切掉内含子并连接外显子,通过连接酶的作用从而形成具有5′帽子和3′尾巴的线性RNA转录本;而circRNAs的形成方式不同于线性RNA的剪切,它主要是由pre-mRNA以反向剪接方式产生,形成共价闭合环状RNA分子,其产生过程通常涉及到本身宿主基因的转录和剪切,并与线性剪切机制相互竞争。根据circRNAs在基因组中的来源及其构成序列的碱基来源,可分为7 种类型:1)仅来源于外显子的环状RNAs(Exon circular RNAs,EcRNAs);2)来源于外显子-内含子的环状RNAs(Exon-intron circular RNAs,EIciRNAs);3)仅来源于内含子的环状RNA(Intron circular RNAs,ciRNAs);4)来源于融合基因的环状RNA(Fusion-circRNAs,f-circRNAs):基因间重排,产生异常融合而形成环状RNAs;5)来源于反义链转录的环状RNAs(Antisense circular RNAs):6)来源于基因间序列的环状RNAs(Intergenic circular RNA);7)TricRNAs成环(tRNA intronic circRNAs):由pre-tRNAs 剪切后形成的tricRNAs[21-23]。
目前公认的生物体内circRNAs形成主要是通过反向剪切来完成,有4种产生机制[24-26]。1)套索驱动成环机制(Lariat-driven circularization):主要通过外显子跳跃,由附近上游外显子的3′端形成剪接供体(Splice donor)和靠近附近下游外显子的5′端形成剪接受体(Splice acceptor)以共价键结合的方式,然后再在剪接酶的作用下清除内含子,形成ecircRNAs(图2A)。2)内含子配对成环机制(Intron pairing-driven circularization):不通过外显子跳跃,而是由上下游外显子两侧的2个内含子上的碱基反向互补配对,如ALU 重复序列,通过配对成环,然后除去内含子进而形成ecircRNAs;这种配对形成的环化机制也可以保留内含子形成EIciRNA,这种形成机制还需要进一步探索(图2B)。3)RNA结合蛋白(RNA-binding protein,RBP)环化机制(RBP driven-circularization):该蛋白可以和两侧翼内含子序列结合,能在内含子之间建立桥梁而碱基产生配对,保证侧翼内含子序列相互结合,从而促进circRNAs 或者EIciRNAs的生成(图2C)。4)内含子套索环化机制(Circular intron RNA):经由剪接体形成的内含子套索产生,要求5′剪接位点富含7个碱基的GU 序列和3′分支位点富含11个碱基的C序列,以避免脱分支酶对其降解,从而保证ciRNAs生成(图2D)。目前关于circRNA形成机制的方式尚有争议,后续的研究工作需要进一步揭示其具体产生机制。

3 circRNAs的功能
circRNAs广泛存在于真核生物中,具有circRNA序列保守性,表达呈现一定的组织特异性和发育阶段性,这表明 circRNA是一种功能分子。随着对circRNA研究的逐步深入,目前相对比较明确和成熟的观点是circRNA具有吸附 miRNA、吸附蛋白、蛋白质翻译和调控基因转录等功能。
3.1 作为miRNA的海绵体
circRNAs序列上具有miRNAs结合位点,能作为miRNA的“海绵吸附体”或竞争性的内源RNA(Competitive endogenous RNA,ceRNA)与miRNAs结合,增强其所调控靶基因的表达[27-28](图3A)。早期的研究报道了一种存在于人和鼠脑组织中的circRNA,被称之为小脑变性相关蛋白1反义转录物(Antisense tothe cerebellar degeneration-related protein 1 transcript,CDR1as)。该circRNA包含有74个能与miR-7结合的位点,能吸附miR-7,充当“超级海绵吸附体”。当过表达CDR1as时,能与miR-7结合,进而促进miR-7靶基因的表达;而当CDR1as的表达被抑制时,则会降低miR-7靶基因的表达水平[29]。此外,另一个有代表性的研究是circSRY(Circular sex-determining region Y),该环状RNA分子来源于小鼠睾丸中Y 染色体中的性别决定区基因。研究发现,circSRY中含有16个miR-138保守结合位点,通过“海绵吸附”作用来抑制miR-138的活性,进而调控miR-138所作用的靶基因[30]。目前越来越多的证据表明circRNAs 在真核生物中作为miRNA 海绵体吸附机制是circRNAs发挥调控功能中的一个普遍现象[31-32]。
3.2 作为蛋白质的海绵体
一些circRNAs可以通过其环状结构上的特定靶点与蛋白质结合,从而形成蛋白质/circRNAs复合体,发挥生理作用(图3B)。例如circRNAs可与RBP相互结合,调控circRNAs 的形成[33]。盲肌蛋白(Muscleblind,Mbl)是一种RBP,可结合其宿主基因第2外显子,当过表达Mbl时,则会促进Mbl基因生成环状分子circMbl,并同时抑制Mbl基因通过常规的剪接方式生成对应线性mRNA转录本;而新生成的circMbl反过来又可以与过量的Mbl结合,进行平衡circMbl和Mbl的生成量,使各自表达趋于稳定[34]。circFoxo3可以与阻滞细胞循环的周期蛋白依赖性激酶抑制剂(Cyclin-denpendent kinase inhibitor1,p21)和细胞周期蛋白依赖激酶2(Cyclin-dependent kinase 2,CDK2)结合,形成circFoxo3-p21-CDK2三原复合物,从而影响CDK2的生物学功能,导致细胞不能正常进入细胞周期,而发生细胞的周期停滞或者细胞凋亡[35-36]。此外,其他研究证实,circPABPN1能与(Human antigen R,HuR)蛋白竞争性的结合,并抑制PABPN1基因的翻译[37]。此外,circANRIL可与pre-rRNA和(Pescadillo homolgue 1,PES1)蛋白进行竞争性的结合,抑制核酸外切酶对pre-rRNA的进一步加工,阻止核糖体的成熟[38]。
3.3 编码蛋白或者多肽
一般认为circRNAs因具有环状结构、无5′和3′端特征,曾被认为无编码蛋白或者多肽能力,通常定义为非编码RNA。1986年,有学者在HDV(Hepatitis delta virus)中发现circRNA分子可以编码病毒蛋白,而且与肝炎疾病的发生相关[39]。合成有内部核糖体进入位点序列(Internal ribosome entry site,IRES)的circRNAs可以招募核糖体来执行蛋白翻译(图3C),而不含IRES的circRNAs 则不能开启翻译机制[40]。Li J等研究发现circRNAs序列中含有的绿色荧光蛋白(Green fluorescent protein,GFP)的开放阅读框(Open reading frame,ORF)能在大肠杆菌体内表达,并产生GFP[41]。另外一项研究也表明,在circRNAs中人工合成缺乏的IRES,可通过插入多个标记有 Flag 编码序列的方式,能保证circRNAs以滚环扩增复制的形式进行翻译[42]。Chen X等首次建立了汇总可编码蛋白的circRNAs数据库(circRNADb),并对潜在可能编码circRNAs中的IRES和ORF进行了注释,并随后用质谱仪验证了他们编码蛋白的表达[43]。来自小鼠和人成肌细胞的环状分子circZNF609 ORF在成环时会产生一个与线性mRNA转录本相同的起始密码子和终止密码子,并不依赖于mRNA结构的方式来编码蛋白[44]。最近的研究表明,富含有N6-腺苷酸甲基化(m6A)的circRNAs序列,可以通过招募eIF4G2和 YTHDF3促进不依赖于5′和3′结构端翻译的开启机制而进行编码蛋白[45]。研究发现,circFBXW7能翻译21 kDa并含有185氨基酸的蛋白质FBXW7,该蛋白能与USP28竞争性的结合来调控原癌基因c-Myc 蛋白的稳定性,来进一步抑制恶性胶质瘤细胞的增殖与发生[46]。Zhang M等研究亦证实了circPINT能通过IRES机制编码出含有87aa的全新多肽(PINT87aa)[47]。后续对这些能编码蛋白的circRNAs需要开展功能验证。
3.4 调控宿主基因的表达
研究表明,circRNAs与线性 mRNA在序列上具有高度的一致性,能通过互补序列与U1小核糖核蛋白(U1 small nuclear ribonucleoproteins,U1snRNP)和RNA聚合酶II(RNA polymerase II,Pol II)结合形成复合体顺式调控宿主基因的表达(图3D)。ciankrd52(Ciankyrin repeat domain 52)是来源于锚蛋白重复结构域52的circRNA,通过特异性的与U1snRNP和Pol II结合来调控宿主基因ankrd52表达,增强了ankrd52转录效率。当特异性敲出ciankrd52 后,能明显降低ankrd52的转录效率,说明circRNA具有顺式调控宿主基因表达功能[48]。此外,研究还发现来源于细胞中的circEIF3J,同样能与U1snRNP相互作用,对宿主基因EIF3J的表达进行调控[35]。Li F等证实,来源于E3泛素蛋白连接酶的环状RNA分子cirITCH与E3泛素蛋白连接酶的3′UTR上同样都具有miRNAs的结合位点,对miR-7、miR-17和miR-214进行海绵吸附。进一步分析表明,cirITCH能与这些miRNAs相互作用,增强E3泛素蛋白连接酶的转录水平[49]。
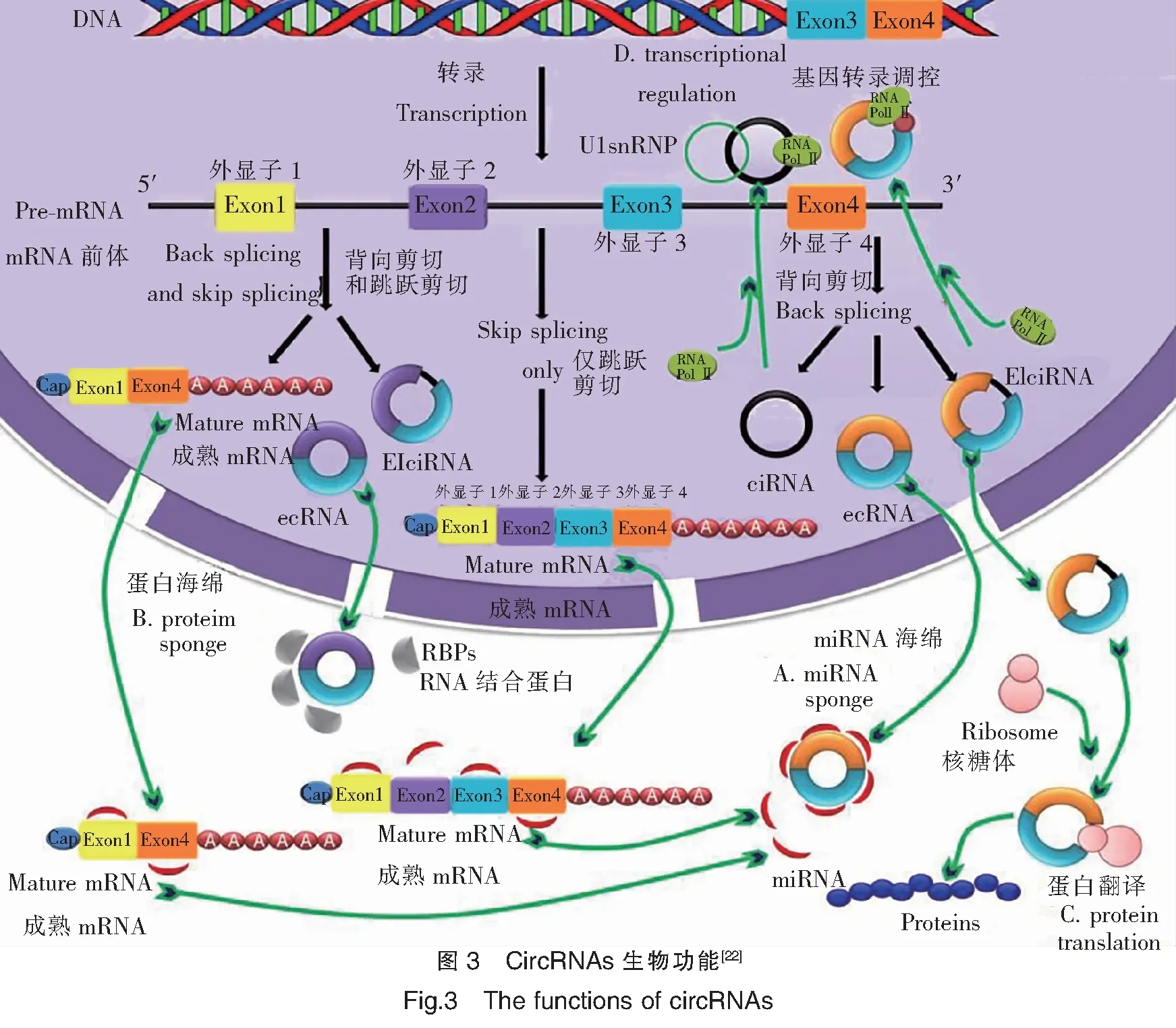
注:A.充当miRNA的分子海绵体; B.充当蛋白的分子海绵;C.蛋白翻译; D.基因转录调控。
4 水产circRNAs研究进展
当前circRNAs 的研究大多数集中在与人类疾病相关方面的医学领域,尤其是在癌症研究报道非常多[26,50-51],而在水产领域相关研究较少。随着对circRNAs深入研究,一些鱼类circRNAs也逐渐被发现与鉴定。Nitsche A等利用转录组测序技术,首次报道了腔棘鱼肌肉可能存在的circRNAs[52]。Shen Y等对斑马鱼(Daniorerio)进行了高通量组织测序,总共鉴定出了3 868个circRNAs,其中发现有1 122个circRNAs在人类、大鼠和腔棘鱼中高度保守,表明这些保守的circRNAs在脊椎动物中可能执行相同的生理功能[53]。进一步的靶点分析显示有29个circRNAs能作为斑马鱼miR-2193的海绵吸附体。随后有学者也对斑马鱼的五个重要组织包括血液、脑、心脏、鳃和肌肉进行了circRNAs鉴定,又新发现3 428个circRNAs,部分circRNAs通过(Quantitative Real-time PCR,qRT-PCR)得到实验验证,发现这些circRNAs都来源于已知注释的蛋白和lncRNAs[54]。Liu H等对斑马鱼胚胎不同发育时相进行了circRNAs鉴定,共发现了1 029 circRNAs,并在囊胚期、原肠胚、体节期和消化管形成期的不同发育阶段都有不同表达水平[55],并对其中随机选取的23个circRNAs证实了它们的表达水平。研究结果暗示着有一些circRNAs可能与调控斑马鱼胚胎发育的关键节点有关。最近,Sun L Y等通过转录组测序在斑马鱼中鉴定了3 431与神经视网膜相关的circRNAs[56]。当前,斑马鱼是已被科学界所熟知的重要模式生物,这些发现的circRNAs能为建立斑马鱼与人类疾病模型提供重要研究价值。
此外,在其他鱼类中也有circRNAs的研究报道。Xu S等在大黄鱼(Larimichthyscrocea)中发现了975个circRNAs,随机选择2个circRNAs得到了验证[57]。靶点预测发现,在鉴定的975个circRNAs中,全部存在有5个以上的miRNAs结合位点;其中的22个circRNAs,发现具有10个以上的miR-430和let-7家族成员结合位点,这些靶点的功能分析还需要进一步实验证实。最近,Hu X等在鲫鱼(Carassiusauratus)肾脏组织中鉴定出了37个circRNAs,靶点预测表明,这些circRNAs作为海绵体能吸附520个miRNAs[58]。而且,通过circRNA-miRNA-mRNA间的互作网络分析显示,miRNAs至少能和1个circRNA和1个mRNA组成调控网络共同发挥作用。circRNAs也与鱼类病毒感染相关联[59]。他们对来自感染GCRV(Grass carp reovirus)第1、3、5、7天草鱼(Ctenopharyngodonidella)肾脏组织和未感染的正常组织进行了cricRNAs测序鉴定,总共鉴定了5 052个新的circRNAs,其中有41个cricRNAs在感染病毒前和感染后的草鱼肾脏组织中呈现差异表达。进一步功能分析显示,这些cricRNAs的宿主基因与金属离子结合,蛋白质泛素化,酶活性和核苷酸结合等生理活动相关。靶点预测显示,有72个miRNAs能与这些差异表达的circRNAs相结合。该研究结果为将来在草鱼中深入研究circRNAs与宿主基因和病原体之间互作机理提供了参考。Fan B等首次从无乳链球菌感染的罗非鱼(Oreochromisniloticus)脑组织中鉴定了11 263个circRNAs[60]。而且在这些鉴定的环状分子中,99% circRNAs都存在有miRNAs的结合位点。进一步差异表达分析显示,有369上调和468下调的circRNAs,对其来源的宿主基因进行KEGG通路分析,结果表明这些基因都与免疫响应有关。
当前在鱼类中鉴定的circRNAs,主要集中在对miRNAs海绵作用和网络调控通路分析,尚无circRNAs功能研究。如何实现其miRNA的海绵机制,未来需要更进一步的功能研究。
5 展望
相比其他的非编码RNA,circRNAs作为RNA世界的一颗新星,其调控作用也日显重要,已成为当前生物与医学领域的研究热点。近年来的研究发现,circRNAs与疾病的发生有着密切的相关性,尤其是与癌症相关的疾病,因此,可以作为将来疾病监测、检测与治疗的一种新型生物标记或者应用靶标分子。目前,虽然已有极少部分的circRNAs功能得到阐释,但仍然有大多数circRNAs功能未知,相关功能研究依然处于初始阶段。随着新一代的高通量测序和生物信息学分析技术快速发展以及circRNAs研究工具和数据库的不断完善,不仅会有大量的circRNAs被发现,而且circRNAs作为重要调控因子的面纱也将被逐步揭开。在水产研究方面,目前的研究还是停留在鱼类以及病原体感染后组织circRNAs的发掘、验证和组织表达谱分析,对于circRNA 的功能研究还仅集中在circRNAs宿主基因的功能注释或miRNA靶点结合网络预测分析,关键的circRNA功能研究还鲜有报道。因此未来还需进一步深入明确circRNAs在鱼类基因中的调控作用,加深与鱼类经济性状和抗病基因的circRNAs筛选、鉴定与功能研究。结果有助于阐明复杂调控网络机制,更好地为将来水产经济动物分子育种和品种改良提供理论研究基础。
