特提斯海变迁对全球动物分布的影响
2021-10-22李枢强赵喆侯仲娥
李枢强,赵喆,侯仲娥
(中国科学院 动物研究所, 动物进化与系统学重点实验室, 北京 100101)
1893年,奥地利地质学家爱德华·修斯(Eduard Suess)根据在阿尔卑斯山和喜马拉雅山等内陆山区分布的海洋动物化石,提出在现今的欧亚大陆南缘曾长期存在过一片现已消失了的古海洋——“特提斯海”(Tethys Sea),以古希腊海洋之神的妻子命名[1].特提斯海主要存在于中生代(2.5亿~6 500万年前),是现今印度洋和大西洋的前身.那时地处欧亚大陆西南端的阿尔卑斯山、伊朗高原和阿拉伯半岛尚未形成,非洲北部淹没在水下,在大西洋和印度洋之间是宽阔的海道,而不是如今狭长的地中海.特提斯海有漫长的消亡史,现存的地中海、黑海、咸海和里海都是特提斯海消失后的遗迹[2].
近年来,解析全球动物多样性分布格局的形成原因和规律成为动物学家和生态保护学家关注的焦点,因为这对预测未来全球动物多样性变化、保护和管理受威胁的物种十分重要.特提斯海曾经覆盖了很大的区域,是很多动物的栖息地.时至今日,全球近1/3的生物多样性热点地区仍分布在特提斯海曾覆盖过的地方,例如地中海盆地、伊朗-安纳托利亚地区、中亚山区、喜马拉雅山区、中国西南山区和印度-西太平洋群岛等[3].无论是植物还是动物,在这些地区都展现出了极高的物种多样性.
20世纪90年代以前,与特提斯海相关的动物多样性研究多为对该区域的化石和现生特有物种的描述[4].大量的研究显示,特提斯海复杂的地质历史对该区域内的动物多样性分布有显著影响[5].例如,化石记录和现存珊瑚礁鱼类的分布格局表明,物种多样性中心从5 000万年前的特提斯海西部(现在的地中海盆地)转移到了现在的印度-西太平洋地区[6-7];化石研究还表明,软体动物的多样性中心在渐新世时期分布在地中海区域,而到了中新世时期则转移到了印度-西太平洋地区[8].进入21世纪,古地理和古环境的重建促使现代动物学家开始逐渐关注动物多样性演化与地质历史之间的潜在联系[9].近10年,高通量测序技术的普及和计算机算力的提升,促进了分子生物学从全球尺度上开展更加精细、准确的动物多样性格局研究[10],以及在地质历史背景下开展大尺度的动物多样性时空演化过程研究[11-12].目前,古生物学和分子系统学研究都证实了发生在特提斯海区域的一系列地质构造事件及其海陆变迁史极大地影响了动物的起源、扩散、分化、存留和灭绝,塑造了特提斯海区域内丰富的物种多样性,影响了现今全球动物多样性的分布和组成.
本文首先给出了最新的特提斯海区域古地理重建模型,然后结合特提斯海区域动物多样性演化研究的最新结果,系统论述了特提斯海演化和变迁对全球动物多样性格局的影响,包括特提斯海演化通过改变栖息地之间的物理联系对动物多样性空间分布的直接影响,以及特提斯海海陆变迁通过引起区域地理和气候环境的变化对动物演化和多样化的间接影响.
1 特提斯海的地理重建及演化历史
20世纪80年代掀起了针对特提斯海的地质学研究浪潮,随后特提斯海的演化历史被逐步复原,并产生了一系列古地理和古环境地图[13-14].这些地图覆盖了约3亿年的时间跨度,并且每一张都有详细的注解.近些年,针对于青藏高原和副特提斯海(Paratethys Sea)等区域,地质学家们提出了更加详细的构造运动模型[15-16].
随着理论的不断完善,目前认为在中生代主要有2个特提斯海先后占据了劳亚(Laurasia)和冈瓦纳(Gondwana)古大陆之间的区域[2,17].第1个是在二叠纪到三叠纪时期(3亿~2亿年前),当所有大陆板块汇聚形成盘古大陆(Pangaea)时,在现在的地中海区域形成了一个向东开放的泛大陆海湾,被称为古特提斯海(Paleo-Tethys Sea)(图1a).但一条大陆物质带(辛梅利亚大陆“Cimmeria”,包含现在土耳其、伊朗、阿富汗、中国西藏、中南半岛和马来西亚等部分陆块)在石炭纪晚期到二叠纪早期开始从冈瓦纳古陆北部分离并向北移动,最终在侏罗纪早期(约1.8亿年前)与劳亚古陆南缘相撞,导致古特提斯海消失(图1a-b).古特提斯海的证据保存在海洋沉积物中,这些沉积物现在并入了从土耳其北部到外高加索、伊朗北部和阿富汗、中国西藏北部以及中南半岛的山脉中.在中生代早期,随着辛梅利亚大陆的北移,新特提斯海(Neo-Tethys Sea)开始形成,通常简称为特提斯海,也称古地中海(图1b).
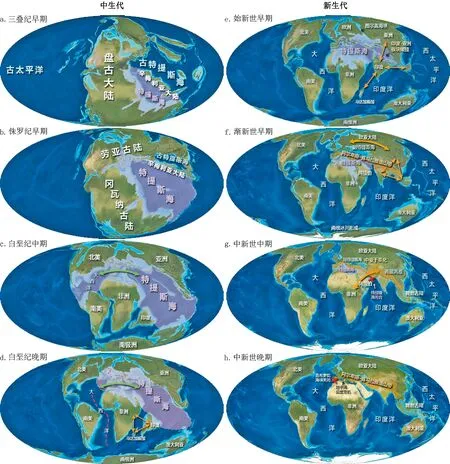
绿色(海洋)和橙色(陆地)双箭头符号表示扩散通道;红色虚线和叉号表示海洋地理屏障.一些重要的地质事件在地图相应的位置上进行了标注.古地理复原图主要参考Scotese的古地图项目[14].图1 中生代(a-d)和新生代(e-h)特提斯海的演化变迁史Fig.1 Mesozoic (a-d) and Cenozoic (e-h) evolution history of the Tethys Sea
伴随着盘古大陆的分离、解体和重组,特提斯海在大约2亿年的时间里经历了从扩张、分裂、收缩直到最后消亡的完整过程[2,14].侏罗纪时期,盘古大陆的分裂导致了特提斯海的逐渐开放,成为中生代时期的主要海洋通道.白垩纪中期(图1c),冈瓦纳古陆和劳亚古陆的解体导致特提斯海分裂,大西洋开始逐渐形成[17-18].冈瓦纳古陆解体后形成的非洲、印度和澳大利亚等陆地板块,以不同速度向北移动,使特提斯海逐渐变窄.板块的运动轨迹大体上可以理解成以现在的地中海西部为轴进行左旋,印度板块移动较快.在白垩纪(图1c-d),海平面的升高[19]导致全球40%的陆地区域被淹没.该时期特提斯海显著拓宽并与向北流动的海道合并,这在特提斯热带动物群和温带北部动物群混合的化石证据中得到证实[2].进入新生代(图1e-f),非洲、印度和阿拉伯板块与欧亚板块间的碰撞、俯冲和挤压变形等构造运动变得活跃,形成了阿尔卑斯-喜马拉雅造山带[20],导致特提斯海从现在的西藏和帕米尔地区向西逐渐退出[21-22].在始新世和渐新世的交界期,造山带将特提斯海分隔,造山带以北巨大的内陆浅海被称为副特提斯海(图1f).副特提斯海曾通过图尔盖海峡与北冰洋连接,通过丹麦-波兰海沟与北海相连.渐新世时期,特提斯海发生了剧烈的海退,欧洲中部和西部的陆地面积增加[16].中新世时期(图1g),非洲-阿拉伯板块与欧亚板块的碰撞导致了位于大西洋和印度洋之间的特提斯海的闭合[13,23].中新世晚期(596万~533万年前,图1h),由于直布罗陀海峡关闭,地中海盆地的大部分海水因蒸发而干涸(地中海盐度危机),特提斯海消失[24-25].进入上新世(约533万年前),由于全球海平面的上升,大西洋的海水迅速重新填满了地中海盆地(赞克尔期洪水),形成了现在的地中海[26].
2 特提斯海演化对动物多样性空间分布的影响
现今全球动物多样性的空间分布格局是长期演化的结果,并且主要是由扩散和隔离这2种过程决定的.扩散使物种能够定植到更广阔的区域,导致种群扩张,促进遗传多样化,提高物种的存活率;隔离则减少了物种的遗传连续性,促进异域新物种的形成.盘古大陆大约在侏罗纪早期开始分裂解体,导致特提斯海逐渐开放并成为全球的主要海洋通道.白垩纪时期,北美和欧洲的分离以及冈瓦纳古陆的解体导致了特提斯海的分裂.新生代以来,板块的聚合和碰撞加速了特提斯海的消亡.这些地质运动极大地影响了全球动物的扩散和隔离,驱动新物种形成,导致不同动物类群形成了相同或相似的多样性空间分布模式.
2.1 特提斯海的开放和分裂
盘古大陆从中生代早期开始持续分裂,并在白垩纪晚期因冈瓦纳古陆的解体达到峰值(图1d),期间特提斯海也逐渐开放并开始分裂.该时期大陆和海洋的变化主导了当时全球的动物多样性格局,同时也对之后全球动物的长期演化轨迹产生了深远影响.例如,中生代化石研究表明,大陆的裂解促进了全球海洋无脊椎动物多样性的增加,出现在中生代晚期的物种多样性峰值与大陆和海洋的分裂状态相一致[27];大陆裂解的顺序和时间决定了中生代恐龙动物群的分化和分布格局[28].大陆的裂解和特提斯海的分裂促进了现生陆生和淡水动物祖先的隔离分化,形成了目前现生分类群的主要分支单元(目或科).例如,两栖动物起源于盘古大陆,随后劳亚古陆和冈瓦纳古陆的分离和破碎化促进了新蛙亚目冠群的分化,形成了许多目前的科级分类单元[29],同时也主导了现今两栖动物全球分布格局的形成[30].类似的多样化模式也出现在现生淡水鱼类[31]、淡水端足类[32]、现代鸟类[33]、蜘蛛目[34],以及鞘翅目昆虫[35]等动物类群中.Sanmartín的团队曾分别对北半球和南半球陆生生物的一般地理分布模式进行研究,结果显示不同时期各大陆动物群之间的关系模式基本符合各大陆的裂解次序,并发现与植物相比,陆生动物的扩散更容易受到海洋的限制,这说明过去的板块运动和海洋变化对现今的陆生动物多样性分布的影响也比植物更为显著[36-37].
直到始新世结束之前,特提斯海为发育中的大西洋和印度-西太平洋的海洋动物提供了一条交流通道[38](图1c-e, 绿色箭头).两地重叠的化石记录反映了这种连通性[39-40].化石证据还表明,全球海洋动物多样性的中心在始新世时期就位于特提斯海的西部区域(现东大西洋和地中海地区)[6],该区域也通常被认为是一个重要的动物多样性起源区[7],且允许动物的双向扩散.特提斯海的连通作用影响了现生海洋动物的高级分类阶元(科或属)的全球多样性组成和分布.例如,对于蒙特波卡(Monte Bolca)的始新世海洋鱼类化石的研究表明,这里鱼类的科级多样性组成与现今大西洋和印度-西太平洋非常相似[41].几乎所有珊瑚礁鱼类的现生科在始新世都是广布的[42].腹足类也展现出了相似的分布格局,在地中海、中亚和印度-西太平洋等地区存在共有的化石[8].海洋有鳞目也曾通过特提斯海遍布全球,并在白垩纪晚期分化成3个主要分支[43].在西藏和新疆地区发现的底栖双壳类和介形类动物化石的年代正好对应特提斯海在白垩纪晚期和古新世扩张至远东地区的事件[44-45].尽管特提斯海在白垩纪时期对海洋鱼类而言是交流的通道,但大西洋的扩张对于海洋底栖和淡水动物来说形成了一道难以逾越的屏障(图1d, 红色虚线).例如,底栖的心形海胆[46]和介形类动物[47]化石在南美洲和非洲呈间断分布.一些现生的甲壳类动物在大西洋两岸的孔隙地下水中孑遗分布,表明它们起源于特提斯海,大西洋的扩张促使它们隔离分化[48].
2.2 特提斯海的收缩和闭合
白垩纪时期,冈瓦纳古陆解体并陆续形成了非洲、印度和澳大利亚等陆地板块.这些板块的北向移动导致了特提斯海海域面积的逐渐缩小并形成了多个临时“陆桥”,为陆生和淡水动物群在各板块间进行扩散和交流提供了机会,直接影响了现今各板块的动物多样性组成.印度-马达加斯加板块大约在1.3亿年前脱离南极板块向北移动[49],约在5 500万年前开始与亚洲板块碰撞并一直持续至今[50](图1c-g).尽管印度板块在与亚洲板块接触之前经历了很长一段孤立时期,但动物地理学研究表明在整个白垩纪晚期,印度板块与邻近的陆块始终保持着生物交流[51].在印度板块北移的过程中,一些能够跨越弱海洋屏障的陆生及淡水动物类群,能够通过印度板块扩散到邻近的大陆(图1d-f, 橙色箭头).例如,蛙类[52]和双翅目昆虫[53]通过印度板块在非洲、马达加斯加和亚洲之间进行扩散.有研究显示印度板块与亚洲板块的生物交换在始新世中期开始加速,并在中新世中期达到峰值,表明印度和亚洲在此期间形成了一个持续的扩散通道[54].一些研究还显示印度板块的北向移动促进了印度和东南亚动物群之间的交流,包括一些蛙类[55]、爬行动物[56]和无脊椎动物类群[57].阿拉伯板块虽然位移较小,但也起到了与印度板块相似的作用.中新世时期,亚洲和非洲的动物群可以通过阿拉伯板块相互扩散[52-53](图1g, 橙色箭头).化石研究也证明在渐新世和中新世交界时期,非洲-阿拉伯和欧亚大陆的哺乳动物开始产生交换[58].此外,渐新世时期,副特提斯海从图尔盖海峡退出[16],为欧洲和亚洲的陆生动物群开辟了迁移通道[36,59](图1f, 橙色箭头).
特提斯海的收缩阻碍了大西洋和印度-西太平洋水生动物的基因交流.很多水生软体动物在始新世中晚期开始分化成大西洋和印度-西太平洋分支[38,60].欧亚地区的鳉鱼在渐新世分化成特提斯海西部(地中海盆地)和东部(阿拉伯盆地)2个主要分支[61].此外,很多特提斯海东西两侧的珊瑚礁鱼类在渐新世-中新世早期出现分化[9,62].在甲壳动物[63]和苔藓虫[64]中也是如此.中新世中期,由于非洲-阿拉伯板块与欧亚板块的碰撞,特提斯海在1 800万~1 200万年前闭合[13,23](图1g).特提斯海的闭合彻底阻止了大西洋和印度-西太平洋海洋动物在赤道附近的扩散.例如,地中海和巴基斯坦腹足类动物化石的相似性从中新世早期开始显著下降,见证了地理隔离的产生[8].一些现生海洋动物类群的地理分化时间和格局也清晰地反映了特提斯海的闭合事件.例如,一些深海腹足类[65]和珊瑚礁鱼类[7,62].
特提斯海的收缩和闭合不仅迫使大西洋和印度-西太平洋的水生类群产生持久的隔离分化,还导致了全球海洋生物的多样性中心从中新世早期的地中海区域逐渐转移到了现在的印度-西太平洋地区.中新世早期,东南亚群岛由于澳大利亚-新几内亚板块和巽他古陆(Sundaland,大部分沉于海下,陆地的部分包括苏门答腊岛、爪哇岛、婆罗洲和附近岛屿)的碰撞而开始形成[66](图1g).新增浅水生境促进了造礁珊瑚的大量增殖,以及其他海洋动物的快速分化[67-68].特提斯海闭合后,大西洋和印度-西太平洋热带动物群只能通过非洲南边海域进行交流[42].在更新世早期由于非洲南边海域形成了一个冷水屏障,交流被进一步限制,但一些鱼类[69]和海龟[70]仍能在间冰期穿过好望角进行双向的扩散并发生辐射演化.
3 特提斯海变迁与现今动物多样性格局形成的潜在联系
地质构造运动能够直接引起区域地理、气候和生境的剧烈变化,对生物的多样化起到促进或限制作用,影响现今的生物多样性组成和分布.进入新生代,伴随着板块运动,欧亚地区的地貌和环境发生了剧烈变化.渐新世时期,阿尔卑斯-喜马拉雅造山带的形成,加速了特提斯海的退出,导致在中亚地区形成了一个包含半咸水到淡水生境的全新生态系统(图1f).中新世,随着海域面积的进一步萎缩,中亚地区最终形成了一片巨大的干旱区[16,21,71](图1g).中新世晚期,直布罗陀海峡的隆升和全球海平面的波动控制了流入地中海的海水流量,导致特提斯海发生盐度危机并一度蒸发殆尽[24-25](图1h).特提斯海的退出导致一些海洋动物形成了孑遗类群,著名的例子是桨足纲动物,它们的分布被限制在特提斯海边缘的海洋洞穴中[72];海陆变迁导致了海洋动物的灭绝,如副特提斯海盆地中的软体动物[73],以及盐度危机时期的地中海鱼类[74];而新出现的陆地、山脉、淡水和洞穴生境则同时为陆生和水生动物提供了新的空白生态位,促进了它们的扩张和快速分化.例如,特提斯海的退出促进了地栖蜘蛛在欧亚地区的迅速扩张和多样化[12,59],同时也促使钩虾从海洋转移到淡水生境并在欧亚大陆的河流中扩张和分化[11].下面总结归纳了一些具有代表性的研究,用以说明新生代以来特提斯海区域的海陆环境剧变与现今动物多样性格局形成的潜在联系.
3.1 阿尔卑斯-喜马拉雅造山带的形成
欧亚大陆目前的地貌特征主要是在新生代以后逐渐形成的,新生代以前,欧亚大陆的许多地区仍被特提斯海所覆盖[2].进入新生代,非洲板块、阿拉伯板块以及印度板块与欧亚板块的构造运动变得更加活跃,导致欧亚大陆南缘全部露出海面并不断抬升,最终形成了一条从西班牙延伸到东南亚的巨大褶皱山脉带——阿尔卑斯-喜马拉雅造山带[20](图1e-h).由于印度板块的俯冲作用,在造山带的东侧形成了有“世界屋脊”之称的青藏高原[75](图1g).造山带的形成加速了特提斯海的退出,并极大地改变了欧亚地区的地貌和气候特征,对欧亚地区动物多样性演化产生了深刻影响.
阿尔卑斯-喜马拉雅造山带的形成重塑了欧亚地区陆生和淡水动物的多样性格局.例如,青藏高原在始新世-渐新世时期的隆升导致了欧亚地区一些地栖蜘蛛的南北分化,山脉的隆升和高原环境的形成促使分布在高原以南的支系快速多样化,并导致分布在高原面及高原以北的部分支系的灭绝,造成了高原两侧物种多样性的差异分布[12].类似的物种多样性分布格局在淡水钩虾中也有发现[76].更新世时期,阿尔卑斯山和喜马拉雅山的低海拔区域以及造山带以南的低纬度地区作为天然的“冰期避难所”,使许多动物类群的物种多样性得以留存[77].这些从末次盛冰期(2.1万年前)中幸存下来的物种在冰期过后往往会再次扩张,来自不同避难所的种群的再次接触增加了物种的遗传多样性,这种现象在欧洲尤为明显[78].
阿尔卑斯-喜马拉雅造山带的形成促进了本地动物的多样化,形成了高加索、伊朗-安纳托利亚、中亚山区、喜马拉雅山区和中国西南山区等多个山地动物多样性热点[3].例如,受喜马拉雅山脉隆升以及南亚季风加强的影响,喜马拉雅地区现生两栖和爬行动物的本地成种和邻域迁入事件在进入中新世以后迅速增加,物种多样性显著升高[79].事实上,在造山带及附近分布的很多动物类群都有相似的多样化模式,例如鸟类[80]、鳞翅目昆虫[81]和蜘蛛[59].山脉的形成还可以通过提供新的生态位来驱动新物种形成,例如在喜马拉雅山地区分布的雀形目鸟类[82].此外,尽管冰川作用经常被视为对生物多样性的破坏,但冰川可以将造山带上连续分布的种群分割成碎片,促进异域物种形成.例如,更新世冰川作用促使在青藏高原地区分布的雪鸡分化成了不同的物种[83].
3.2 淡水生境的出现
渐新世时期(图1f),伴随着副特提斯海海域面积的不断萎缩,海洋生境逐渐消失[16,21].从海洋到半咸水及淡水生态系统的转变,导致了副特提斯海地区的珊瑚、有孔虫和软体动物的灭绝[73,84],同时逐渐淡化的水体生境促进了半咸水和淡水软体动物的迅速演化[85].由于特提斯海闭合切断了特提斯海与印度-西太平洋之间的联系,同时特提斯海与大西洋的连接也受到了限制(图1g),副特提斯海地区的海洋生境大约在1 160万年前彻底消失[86].不同动物群的化石反映了副特提斯海地区发生从海洋到淡水生境转变的时间大约在1 200万年前.该时间也通常被认为是现生黑海-里海软体动物[87]和甲壳动物[88]的起源时间.
特提斯海区域新形成的淡水环境驱使海洋起源的动物转移到新的淡水生境中[89].一些研究显示海洋和淡水动物类群之间存在明显的分歧,表明生境转换并不频繁,从海洋到淡水生境的定殖只发生了1次,并导致了随后的辐射演化.例如,在欧亚大陆广泛分布的淡水钩虾在系统发育树上形成单系,在始新世晚期从海洋生境转移到淡水生境,并在淡水生境中迅速分化和扩张[11];印度-亚洲板块碰撞促进了河鲀在始新世从浅海到淡水的生境转换[90].事实上,类似的从海洋到淡水生境的单一起源模式在一些鱼类[91]、腹足类[92]和桡足类[93]中也有发现,通常与重大的地质事件相关.此外,一些类群在海洋和淡水生境间发生了多次转移.地中海区域的鰕虎鱼存在多次生境转移的现象[94-95].这种模式在一些全球分布的海洋鱼类,以及在海洋和淡水栖息地间迁徙的洄游性鱼类中经常出现[91].
3.3 中亚地区的干旱化
随着副特提斯海的退出,欧亚内陆地区出现了受限的蒸发海洋环境.自始新世晚期以来,中亚已建立了类似沙漠的干旱环境[96].中新世中期(图1g),全球气温下降,山脉(如扎格罗斯山脉和天山山脉)和高原(如伊朗高原、安纳托利亚高原和青藏高原)的隆升对空气流动形成了阻碍,在造山带的北侧形成雨影区,加剧了中亚地区的干旱化[97].目前中亚地区仍然主要由沙漠、草原、干旱林地等干燥的生境构成,这导致了一些喜欢湿润环境的动物在欧亚地区间断分布,两栖动物就是个明显的例子[98].中亚的干旱化还促进了一些动物的多样性分化.例如,中新世晚期,干旱的加剧导致了亚洲麻蜥的分化[99].亚洲跳鼠自晚中新世以来的辐射演化与中亚日益干旱形成的开阔生境密切相关[100].有证据表明,由于干旱和复杂的地形,更新世冰期对中亚地区生物的影响不如北半球其他地区显著[97].
3.4 地中海发生盐度危机
在596万~533万年前的墨西拿期(中新世晚期),直布罗陀海峡的隆升和全球海平面的波动控制了流入地中海盆地的海水流量,持续的蒸发造成地中海盆地海平面的急剧下降,广泛的盐积累导致地中海发生盐度危机[24-25](图1h).海洋生境的消失,引发了地中海盆地海洋动物群的灭绝[101].苔藓虫的化石记录表明海洋动物群在特提斯海闭合后大量减少,并在盐度危机时期接近绝灭[8].浮游生物和珊瑚礁环境的减少进一步印证了地中海盐度危机造成的灭绝事件,该事件也同时波及了以珊瑚礁为栖息地的鱼类、双壳类和腹足类等动物类群[74,102].上新世时期,直布罗陀海峡再次打开,大西洋海水的重新充填结束了地中海的盐度危机[26],为大西洋动物再次定殖地中海提供了一条海上通道,因此地中海和大西洋动物种群之间的遗传差异很小[103].此外,由于在地中海和大西洋之间能够自由扩散,盐度危机结束以后,地中海区域的海洋特有物种数量明显下降[74].
地中海盐度危机导致了一个名为“Lago-Mare”的湖泊系统的形成,使一些海洋物种适应了淡水生境[104].随着大西洋海水重新涌入地中海盆地,有些物种仍维持了淡水生境,因此与广盐性的物种相隔离,促进了现今地中海区域淡水特有物种的形成.例如,地中海的沙鰕虎鱼类群在盐度危机时期适应了淡水生境,危机结束以后,这些物种在海水和半咸水生境中再次定殖并产生了辐射演化,而孤立的淡水沙鰕虎鱼则成为了特有物种[95].
地中海盐度危机时期,干涸的地中海盆地使原来孤立的岛屿与大陆相连,促进了大陆动物群向地中海岛屿的扩散.当地中海盆地重新恢复海洋环境后,一些陆生和淡水动物由于海水的隔离发生了分化,形成了特有物种.例如爱琴海岛屿上的涡虫[105]、从小亚细亚半岛分布到地中海盆地的淡水螃蟹[106],以及塞浦路斯的侧褶蛙[107].此外,一些淡水鱼类在地中海北部的河流中存在着许多特有物种,这是由于地中海的干涸使淡水河道系统产生了连接,当地中海盆地被海水重新填充后淡水连接被中断所导致的[108].
3.5 洞穴和湖泊的形成
中新世晚期,地下水侵蚀导致了地中海边缘洞穴系统的形成,为地上的淡水祖先定植地下生境提供了合适条件[109].洞穴生境使物种更容易发生隔离分化,形成较高的区域物种多样性[110].位于巴尔干半岛的迪纳里克(Dinaric)喀斯特地区存在着相当多样化的地下动物群[111].有研究显示迪纳里克的洞穴虾类在530万~370万年前分化,与当地喀斯特地貌开始形成的时间相一致[109].同样的情况也出现在其他一些甲壳动物中,如等足类[112]和端足类[113]等.海洋洞穴物种则是直接起源于海洋,尽管它们的非洞穴海洋近亲通常已经灭绝或是生活在深海[114].海洋洞穴类群离散的分布可能是板块运动将它们分离在特提斯海边缘导致的.著名的例子如甲壳动物(桨足类和端足类),它们离散的全球分布范围和特提斯海边缘极其吻合[48,115].目前特提斯海隔离假说在一些钩虾类群中得到了验证,研究结果显示其主要的分化发生在9 600万~8 300万年前(图1c),与特提斯海的演化密切相关[116].
更新世时期,冰川的融化在阿尔卑斯-喜马拉雅造山带上形成了大量的高山湖泊,促进了淡水动物的扩散和分化.例如,高山湖泊钩虾(Gammaruslacustris)占据了从南欧到北欧的冰川湖泊,但种群间的遗传分化程度很低[117];而它的近缘物种(G.alpinus)则经历了由阿尔卑斯山高山湖泊扩张导致的隔离分化[118].生活在比利牛斯山高山湖泊中的水蚤也是个很好的例子[119].
4 总结和展望
特提斯海存在了约2亿年的时间,曾覆盖了很大的区域,其漫长的演化变迁史对特提斯海区域动物多样性演化乃至全球动物多样性格局都产生了深远影响.本文通过对特提斯海区域的地质学研究和动物多样性研究进行系统回顾和总结,详细论述了特提斯海演化和变迁是如何驱动全球动物多样性演化的,揭示了现今全球动物多样性格局的形成与地质演化之间的潜在联系.通过比较,发现同一地质事件对于陆生、淡水和海洋动物的影响往往是不同的,甚至呈现出此消彼长的关系.例如特提斯海的开放为海洋动物开辟了交流通道,但促使陆生和淡水动物发生隔离分化;海洋环境的消失引起了海洋动物的灭绝,但也为陆生和淡水动物提供了新的生态机遇.在另外一些例子中,似乎又可以找到一些共同的规律.例如地质构造运动活跃的地区往往物种多样性较高,无论是在阿尔卑斯-喜马拉雅造山带还是印度-西太平洋地区,所有动物都展现出了极高的物种多样性.因此,特提斯海演化和变迁对全球动物多样性格局的影响依然是个令人着迷的话题.
随着地质学研究的不断发展,特提斯海的演化变迁史逐渐明晰,这为大尺度的动物多样性格局研究提供了一个极好的地质学背景.但国内目前在特提斯海变迁主题下开展的动物多样性研究还相对较少,期待本文的发表能够给中国的特提斯海区域动物多样性和动物地理学研究带来一些启发和线索.毕竟,从进化生物学角度上来看,特提斯海的演化变迁是驱动全球动物多样性演化的重要因素之一,而如果只关注欧亚地区的动物多样性演化,可以说几乎都与特提斯海的演化变迁有关.此外,尽管目前成熟的系统发育和生物地理学研究方法已为动物多样性演化研究提供了良好的框架,但目前许多研究都还只局限于在某一特定时间范围内和区域地理尺度上开展单一类群的研究.因此,我们提出未来应在更广泛的时间和地理尺度下开展包括更多动物类群的多样性演化研究,以揭示特提斯海变迁驱动全球动物多样性演化的普遍规律.这将有助于预测未来环境变化对全球动物多样性产生的影响,提高动物多样性保护效率.
致谢:衷心感谢中国科学院动物研究所姜统尧、邵丽丽和张小庆在图版绘制及文献翻译和校对过程中给予的帮助.
