两类典型荒漠生物结皮覆盖土壤碳通量月变化特征及其影响因子
2021-10-11韦应欣周利军卜崇峰鞠孟辰李亚红李炳垠
韦应欣,周利军,卜崇峰,骆 汉,鞠孟辰, 王 春,李亚红,郭 琦,李炳垠
(1.西北农林科技大学 水土保持研究所,陕西 杨凌 712100;2.中国科学院 水利部 水土保持研究所,陕西 杨凌 712100; 3.长江水利委员会 长江流域水土保持监测中心站,武汉 430010;4.黄河水利委员会 黄河上中游管理局,西安 710021)
生物土壤结皮(Biological Soil Crust,BSC,简称生物结皮)是由光合自养型的蓝藻、绿藻、地衣、苔藓类植物和异养型的真菌、细菌、古细菌等生物群落,通过菌丝体、假根和分泌物等与表层土壤颗粒胶结而成的复合体,结皮类型以藓结皮和藻结皮为主,藓结皮是以苔藓类植物为优势组成的生物结皮,通常呈绿色、金黄色、棕色或黑色,藻结皮是以蓝藻、绿藻为优势组成的生物结皮,通常比周围土壤的颜色黑,覆盖了旱区约40%的地表面积[1]。生物结皮作为具有光合活性的土壤—大气系统界面层,在光照条件下,固定大气中的CO2,其光合固定的碳是旱地碳输入的重要来源[2],同时,也能通过其自身所包含的有机成分与生命体的呼吸作用向大气中释放CO2[3]。因此,生物结皮是影响干旱与半旱区生态系统碳循环过程的重要因素[4-5]。
在干旱半干旱区生态系统中,水分是主要的调控和限制因子,能够直接影响生物结皮的生理活性[6-8]。旱地土壤表面湿润时间的长短决定了生物结皮覆盖土壤的碳源—汇效应[6]。此外,生物土壤结皮存在净碳固定的“水分阈值”,低于阈值时出现碳损失,较好的水分条件有利于生物结皮的光合固碳过程[8]。水分充足条件下,温度是影响生物结皮覆盖土壤碳通量的关键因子[9]。高温条件下生物结皮的呼吸活性强于光合活性,使其CO2排放量增加,净碳吸收量减少,低温条件下则相反[7,9]。此外,温度可以通过影响结皮层含水量与湿润周期间接影响生物结皮的光合、呼吸过程[6,9-13],高温会增加结皮层水分蒸发,缩短其代谢活动时长,从而减少净碳输入量[13-14]。根据气候变化情景预测,未来世界上大多数地区的气温将升高,导致降水模式发生变化,干旱期延长[15-16],可能使生物结皮在未来65 a内覆盖面积减少25%~40%[17],气候变暖与生物结皮覆盖度下降的协同作用将削弱旱地吸收大气CO2的能力,在中长期内减少土壤碳储量[18]。因此量化生物结皮覆盖土壤碳通量对大气、土壤温度与湿度等环境因子的响应,对于预测旱地生态系统碳储量的变化以及了解全球碳循环和平衡的意义至关重要[19]。
当前国内外关于生物结皮覆盖土壤碳通量的研究主要集中于极地和苔原地区、美国西南部莫哈维沙漠、科罗拉多高原、以色列内盖夫沙漠和我国的古尔班通古特沙漠、腾格里沙漠、黄土高原等地区[3,20]。这些地区的研究对生物结皮覆盖土壤碳通量的影响因素探讨不足[21],且多集中于室内模拟研究,原位观测数据仍然较少[11],不同类型生物结皮覆盖土壤碳通量研究还很匮乏[11],多数研究只关注生物结皮覆盖土壤碳损失或碳固定的某一方面[2,20-21],很少综合考虑多种环境因素对生物结皮覆盖土壤碳通量的影响,并探讨区域气候变化和土地利用方式对生物结皮覆盖土壤碳通量的影响。这些问题极大地限制了旱区土壤碳交换的估算,及未来气候变化对土壤与大气碳交换影响的预测[22]。毛乌素沙地位于我国农牧交错区,农业、畜牧业发达、盛产石油、煤炭等资源,是生态恢复及评价实践研究的重点区域。然而该区目前还没有生物结皮覆盖土壤碳通量月动态变化及其驱动因子的研究。鉴于此,我们选择毛乌素沙地中处于生长季(6—11月,水、温因子变化剧烈)的藓结皮、藻结皮,进行两类生物结皮覆盖土壤的呼吸速率(DR)、净碳通量(NCF)、光合速率(Pn)以及土壤温度、含水量、光照强度等相关环境因子的动态检测,以了解荒漠生态系统中藓结皮、藻结皮的碳通量月变化特征及其与环境因子的关系,探讨气候变化对两类生物土壤结皮覆盖下碳通量的潜在影响,为该区生物土壤结皮的碳交换研究提供参考资料。
1 研究区概况
研究区地处内蒙古自治区鄂托克前旗敖勒召其镇(东经107°50′40.51″,北纬38°05′53.39″),海拔1 344 m,属于毛乌素沙地西南部。该区气候干旱少雨,风大沙多,日照充足,属中温带温暖型干旱、半干旱大陆性气候。年均气温7.1℃,最低月平均气温-10.2℃,最高月平均气温22.4℃。降水年际变率大,时空分布不均匀。年平均降水量265 mm,主要集中在6—9月,占全年降雨量的70%,年均蒸发量2 514.8 mm。样地位于固定沙丘顶部,样地内土壤类型为风沙土,植被种类较少,主要为沙蒿(Artemisiaordosica)、沙柳(SalixcheilophilaSchneid),藓、藻两类结皮发育良好,分布广泛[23]。
2 方法与数据
2.1 研究方法
2.1.1 试验设计 选取植被灌丛间发育较的藓结皮、藻结皮(两类结皮的覆盖度接近100%,远离植被冠层及其根系,距离植株2.5 m左右(图1A—B)作为研究对象。在鄂托克前旗境内,为两类结皮各选取1处地势平坦、生境条件相似的样地(30 m×30 m)。在样地内生物结皮发育状况相近、不受植被影响的地点设置3个2 m×2 m的样方作为重复,共6个样方,调查样方内生物结皮生长状况及其下伏土壤理化性质(表1)。为降低生物结皮发育状况差异以及样方周围植被的干扰,试验前1月(2016年5月初)移除样方内除生物土壤结皮外其他植物,放置时先将生物土壤结皮润湿,然后在每个样方中心垂直嵌入1个PVC环(不透光,高10 cm,直径15 cm,图1A)和1个有机玻璃环(透光率>99%,高10 cm,直径15 cm,图1B),PVC环和有机玻璃环插入深度为6 cm,插入时保证环内结皮完整,以利于扰动土壤尽快恢复,降低测定误差。表1为调查样方内的各项基本指标。

图1 土壤碳通量观测样点
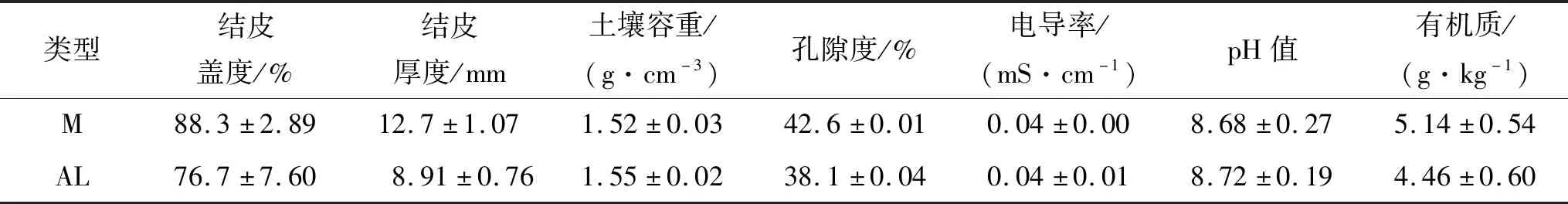
表1 样点生物土壤结皮发育状况及下伏土壤理化性质
2.1.2 指标观测 试验开展于2016年6—11月,为两类生物结皮的生长期,采用LI-8100自动土壤CO2通量系统(LICOR,Lincoln,Nebraska,USA,图1C)连接便携式测量室8100-103和有机玻璃气室LI-104测定BSC土壤CO2通量[24]。在PVC环内,用LI-8100系统连接便携式测量室8100-103(气室使用金属材质,闭合时,密闭遮光,在黑暗条件下测定生物结皮层呼吸作用和下覆土壤呼吸作用总和,图1C)测定两类生物结皮覆盖土壤呼吸速率(DR);在有机玻璃环内,用LI-8100系统连接有机玻璃气室LI-104(气室采用有机玻璃制成,透光率>99%,在光照条件下测定生物结皮层光合作用、呼吸作用以及下伏土壤的呼吸作用总和,图1C)测定两类生物结皮覆盖土壤净碳通量(NCF)。每月选取晴朗的天气(测定3 d前无降雨事件),观测时间为7:00—19:00,观测频次为每2 h一次,每次观测3 min,包括测量时长90 s,测量前与测量后气体平衡时间各45 s,测定前,去除处理样方中的杂草,以降低测定误差。
两类生物土壤结皮的光合速率(Pn)计算公式为[3]:
Pn=DR-NCF
式中:DR为土壤呼吸速率;NCF为净碳通量。Pn为正且光合速率越高,表示结皮的光合速率越高,碳吸收能力越强。
同时,用数字式地温计(MannixTM,USA)测定土壤2,5,10 cm处的土壤温度(ST);用TDR(TRIMEPICO32)测定土壤2,5,10 cm处的土壤含水量(SWC);用数字式温湿度计(GM1361)测定近地表空气温度(AT)和空气湿度(AM);用数显照度计(TES1339)测定光照强度(Li)。
2.2 数据处理
消除光照强度变化对藓结皮与藻结皮光合速率影响的误差,光照强度采用了藓结皮与藻结皮样方内多次测量结果的均值。采用逐步回归分析法对两类生物结皮覆盖土壤碳通量与环境因素(2,5,10 cm土壤温度、2,5,10 cm土壤含水量、光照强度、近地表空气温湿度)进行多元线性回归分析。采用了典范对应分析(CCA)评价了环境参数解释两类结皮的光合速率及其覆盖下的土壤碳通量变化的能力。基于最大调整R2和最小AIC(Akaike Information Criterion,赤池信息量原则)选择了最佳拟合的CCA模型。拟合的最佳CCA模型中所有变量的方差膨胀因子(VIF)均低于10。采用线性模型拟合两类结皮覆盖土壤净碳通量与土壤温度、含水量的关系。数据分析与绘图采用R软件(4.0.2版本)和R-studio(1.3.1093版本)完成。
3 结果与分析
3.1 生物结皮覆盖土壤碳通量月变化特征
如图2所示,藓结皮(M)、藻结皮(AL)的土壤呼吸速率(DR)月变化为先升后降,在8月出现峰值[M:1.96 μmol/(m2·s);AL:1.66 μmol/(m2·s)];光合速率(Pn)月变化为先升后降,9月出现峰值[M:2.75 μmol/(m2·s);AL:1.39 μmol/(m2·s)];净碳通量(NCF)月变化为先升后降再升,7月出现峰值[M:1.38 μmol/(m2·s);AL:1.43 μmol/(m2·s)],9月出现谷值[M:-1.24 μmol/(m2·s);AL:-0.18 μmol/(m2·s)]。
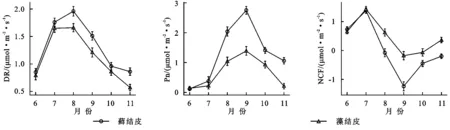
图2 两类结皮覆盖土壤碳通量月变化特征
生长期中,藓结皮覆盖土壤呼吸速率与光合速率显著(p<0.05)高于藻结皮,净碳通量显著低于藻结皮。两类生物结皮覆盖土壤净碳通量均大于0,表现为净碳释放(表2)。

表2 不同处理碳通量的差异 μmol/(m2·s)
3.2 生物结皮覆盖土壤环境因子月变化特征
土壤温度(ST)、土壤含水量(SWC)、光照强度(Li)、空气温度(AT)、空气湿度(AM)的月变化均为“先升后降”的趋势,土壤温度在7月达峰值,光照强度、近地表空气湿度、近地表空气温度均8月达峰值,土壤含水量在9月达峰值(图3)。
藓结皮覆盖土壤含水量、近地表空气湿度均显著(p<0.05)高于藻结皮,而土壤温度、近地表空气温度均显著低于藻结皮(表3)。

表3 不同处理环境因子的差异
3.3 生物结皮覆盖土壤碳通量的关键影响因子
逐步回归分析(表4)显示:5,10 cm土壤温度、2 cm土壤含水量、光照强度是影响藓结皮覆盖土壤呼吸速率的关键因子;2,5 cm土壤温度,5,10 cm土壤含水量,以及光照强度是影响藻结皮覆盖土壤呼吸速率的关键因子。2 cm土壤含水量是影响藓结皮光合速率的关键因子;2 cm土壤温度、2,5 cm土壤含水量、近地表空气温度是影响藻结皮光合速率的关键因子。2 cm土壤含水量是影响藓结皮土壤覆盖土壤净碳通量的关键因子;2 cm土壤温度、5 cm土壤含水量、近地表空气温度是影响藻结皮土壤覆盖净碳通量的关键因子。

表4 两类生物结皮覆盖土壤碳通量与环境因子的逐步回归方程
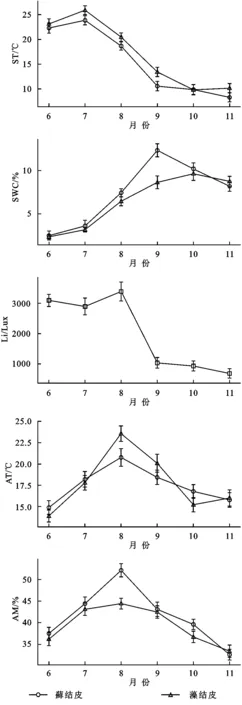
注:藓结皮与藻结皮光照强度(Li)一致,具体原因详见实验方法。
3.4 生物结皮覆盖土壤碳通量与环境因子的关系
CCA分析结果(图4,表5)显示:仅第一排序轴(CCA1)就解释了藓、藻两类生物结皮与环境关系的100%,蒙特卡洛置换检验通过(p=0.001),环境因子解释了藓结皮覆盖土壤碳通量变异的58%,解释了藻结皮56%,对于两类结皮覆盖土壤碳通量与环境因子关系的还原能力和预测能力均较强。所有的环境因子中,2,5,10 cm土壤温度,2,5 cm土壤含水量是影响藓结皮土壤覆盖土壤碳通量的关键因子;2,5,10 cm土壤温度,光照强度、空气湿度是影响藻结皮覆盖土壤碳通量的关键因子(表5)。
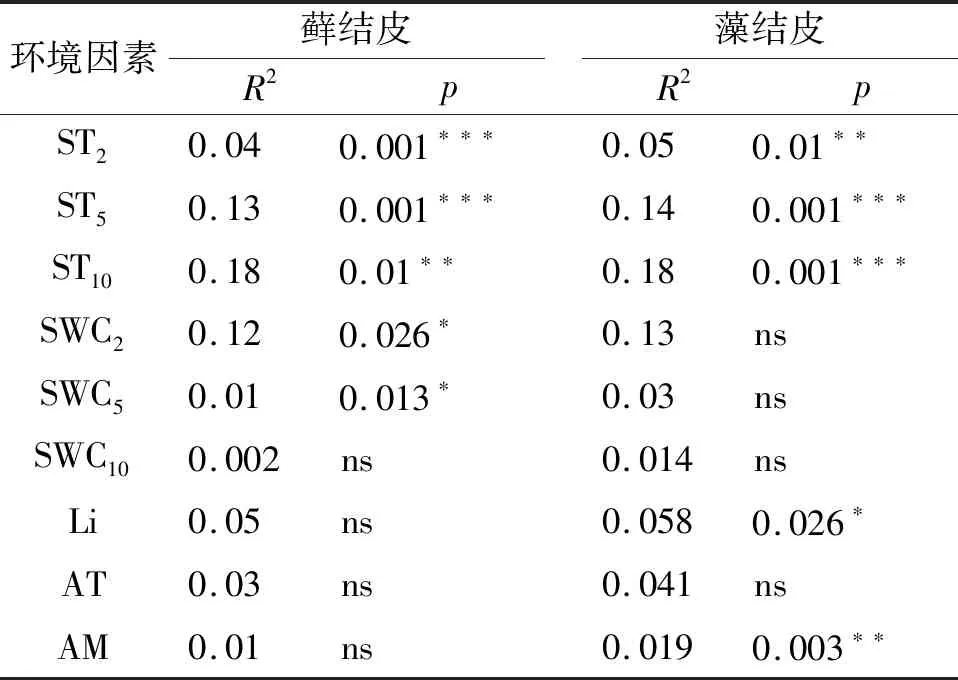
表5 两类结皮覆盖土壤碳通量与环境的分析结果
3.4.1 呼吸速率与环境因子的关系 2 cm土壤含水量与藓结皮覆盖土壤呼吸速率(DR)负相关,与藻结皮正相关;2,5,10 cm土壤温度及5,10 cm土壤含水量与两类结皮覆盖土壤的呼吸速率均正相关;光照强度与藓结皮覆盖土壤呼吸速率正相关,与藻结皮相关性较弱;近地表空气温度与藻结皮覆盖土壤呼吸速率正相关,与藓结皮相关性较弱;近地表空气湿度与藻结皮覆盖土壤呼吸速率正相关,与藓结皮相关性较弱(图4)。
3.4.2 光合速率与环境因子的关系 两类结皮光合速率(Pn)均与2 cm土壤含水量正相关,与2,5,10 cm土壤温度、5,10 cm土壤含水量负相关;光照强度与藓结皮光合速率负相关,与藻结皮正相关;近地表空气温度与藓结皮光合速率负相关,与藻结皮相关性较弱;近地表空气湿度与藓结皮光合速率正相关,与藻结皮负相关(图4)。
3.4.3 净碳通量与环境因子的关系 两类生物结皮覆盖土壤净碳通量(NCF)与5,10 cm土壤温度、光照强度、近地表空气温度正相关,与2,5,10 cm土壤含水量、近地表空气湿度负相关(图4)。
3.5生物结皮覆盖土壤净碳通量变化对土壤温度、含水量变化的响应
CCA分析显示(图4),2 cm土壤含水量(SWC2),10 cm土壤温度(ST10)是两类结皮覆盖土壤碳通量的关键影响因素。拟合ST10,SWC2与两类生物结皮覆盖土壤净碳通量的关系,对比多种线性与非线性的拟合模型,发现线性模型的拟合度与显著性最高,能较好的拟合变量间的关系。其他土层温度与含水量对两类结皮覆盖土壤碳通量影响较小,或与SWC2,ST10对生物结皮覆盖土壤净碳通量的拟合曲线变化规律相似,不再赘述。
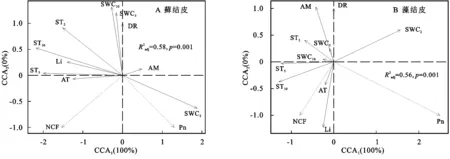
图4 两类结皮覆盖土壤碳通量与环境因子的CCA排序
3.5.1 对土壤温度变化的响应 线性回归分析结果(图5)显示,藓结皮、藻结皮覆盖下土壤净碳通量(NCF)与10 cm土壤温度(ST10)呈线性正相关关系(藓结皮:R2=0.55,p<0.01;藻结皮:R2=0.56,p<0.01)。两类结皮覆盖下土壤净碳通量存在净碳固定(NCF<0,生物结皮开始净碳吸收的土壤温度值,高于此值出现碳损失现象)的土壤温度阈值。藓结皮的阈值(ST=13.5℃)高于藻结皮(ST=7.3℃)。
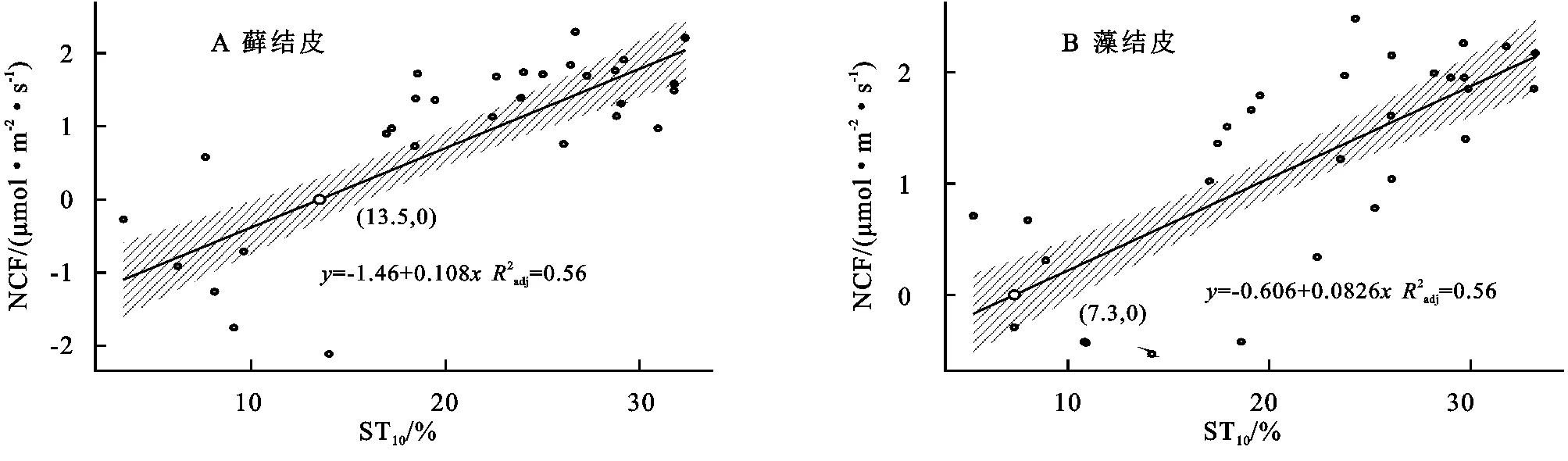
图5 两类结皮覆盖土壤净碳通量与10 cm土壤温度的关系
3.5.2 对土壤含水量变化的响应 线性回归分析结果(图6)显示,藓结皮、藻结皮覆盖下土壤净碳通量(NCF)与2 cm土壤含水量(SWC2)呈线性负相关关系(藓结皮:R2=0.34,p<0.01;藻结皮:R2=0.49,p<0.01)。两类结皮覆盖下土壤净碳通量存在净碳固定(NCF<0,生物结皮开始净碳吸收的土壤含水量值,低于此值出现碳损失现象)的土壤含水量阈值。藓结皮的阈值(SWC2=8.9%)高于藻结皮(SWC10=11.4%)。
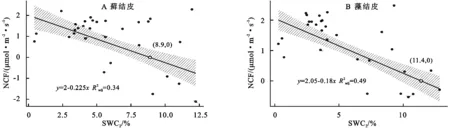
图6 两类结皮覆盖土壤净碳通量与2 cm土壤含水量的关系
3 讨 论
3.1 生物结皮覆盖土壤碳通量月动态变化及其驱动因子
两类生物结皮覆盖土壤的呼吸速率月动态变化为先升后降,在光照强烈、土壤含水量适中的8月达到峰值(图2—3)。在腾格里沙漠,Guan等[21]也发现类似规律:藓、藻两类结皮覆盖土壤呼吸速率在土壤温度最高、降雨适中的7月达到峰值。逐步回归(表4)与CCA分析(图3,表4)表明:5,10 cm土壤含水量,2,5,10 cm土壤温度以及光照强度与两类生物结皮土壤覆盖呼吸速率正相关性较强,是驱动两类结皮土壤呼吸速率月动态变化的关键因子。土壤温度的升高会刺激土壤酶活性,加速土壤有机质的分解[25]。土壤含水量的升高则会直接影响养分矿化速率和土壤微生物活性,改变土体氧化还原电位和气体交换速率,进而改变土壤碳通量[26]。光照强度的升高(图2)会促进两类结皮的生理活性,为其自身呼吸提供了更多碳源[8]。8月前,干燥的生物结皮土壤已经积累了一定的呼吸底物,进入8月,土壤含水量上升、光照强度达到峰值,因此,两类结皮覆盖土壤呼吸速率迅速上升,并在8月达到峰值。生长期中,藓结皮覆盖土壤呼吸速率显著高于藻结皮(表2)。不仅是因为藓结皮比藻结皮有更强的呼吸活性,还存在其他影响因素。一方面藓结皮比藻结皮能够更长时间的保持水分有效性,减缓水分蒸发和下渗,维持其自身呼吸活性。另一方面藓结皮比藻结皮覆盖土壤孔隙度更大,黏粒和养分含量更高,更有利于其下伏土壤呼吸作用[3]。
两类生物结皮的光合速率与土壤含水量月变化同步,先升后降,9月达到峰值。逐步回归(表4)与CCA分析(图4,表5)表明:2 cm土壤含水量与两类生物结皮光合速率正相关性较强,是驱动两类生物结皮光合速率月动态变化最关键的因子,此外2,5,10 cm土壤温度、光照强度与两类生物结皮光合速率负相关性较强,也是影响两类结皮光合速率月动态变化的重要因子。进入9月,光照强度的大幅下降,土壤含水量的大幅上升(图3),一方面使生物结皮土壤的湿润周期变长,另一方面使土壤温度与近地表空气温度处于较低的水平。因此两类生物结皮的光合速率在9月达到峰值。尽管藓结皮光合速率与光照强度显示出了较强的负相关性(图3),太阳辐射最为强烈的8月,藓结皮光合速率仍然处于较高水平。这可能是因为水分充足时,藓结皮的光合过程能够很好地适应光照强度的变化。例如,赵允格等[13]发现光照对黄土丘陵区藓类结皮的光合作用没有抑制作用。同时,藻结皮光合速率在8月同样处于较高水平,表明藻结皮光合作用过程受光照强度变化影响较小。例如,Karsten等[10]发现非洲南部干旱地区的热沙漠和美国阿拉斯加州的冷沙漠的BSC中的4种丝状绿藻在较低光照下光合产氧量依然较高,而高光照条件无抑制作用。藻结皮光合速率与光照强度存相关性较弱(图4)也证实了这点。整个生长期中,藓结皮光合速率显著高于藻结皮。原因在于藓结皮持水能力更强,叶绿素a,b含量高,其光合活性更高,能够吸收更多的CO2。此外,藓结皮覆盖土壤含水量显著高于藻结皮、土壤温度显著低于藻结皮(表3),更有利于其进行光合作用[6,26]。
两类结皮覆盖土壤净碳通量月变化为先升后降再升,7月出现峰值,与土壤温度月动态变化出现峰值的月份一致,9月出现谷值,与土壤含水量月动态变化出现峰值的月份一致。CCA分析表明(图4),两类生物结皮覆盖土壤净碳通量与5,10 cm土壤温度、光照强度、近地表空气温度正相关性较强,与2,5,10 cm土壤含水量、近地表空气湿度存在较强的负相关性较强。毛乌素沙地干湿季分明[27],在降雨集中、光照强烈、土壤温度最高的7月(干季),两类生物结皮覆盖土壤的呼吸速率明显高于光合速率,因此净碳通量达到峰值,而在光照适宜、土壤含水量较高的9月(湿季),光合速率明显强于呼吸速率,因此净碳通量达到谷值(图2—3)。整个生长期中,两类结皮净碳通量均大于0(表2),表明两类生物结皮覆盖土壤表现出碳源作用。可能是因为两类生物结皮的光合速率较低,不能抵消土壤呼吸所致[3]。藓结皮净碳通量显著低于藻结皮(表2)。表明藓结皮的净碳固定能力更强,能够通过光合作用吸收更多自身及其下伏土壤呼吸产生的CO2。
3.2 生物结皮覆盖土壤净碳通量对气候变化的潜在响应
安莉娟[28]研究显示:未来毛乌素沙地气温呈上升趋势,年降水减少,干旱加剧。两类生物结皮覆盖土壤净碳通量的变化与土壤温度及近地表空气温度升高正相关较强,与2 cm土壤含水量及近地表空气湿度负相关性较强(图4),表明气温的上升与降雨量的减少会增加两类生物结皮碳损失的风险。虽然生物结皮光合过程对温度升高的响应具有弹性[29],但鉴于适宜研究区两类生物结皮净碳固定(NCF<0)的土壤温度范围较窄(图5),两类生物结皮的群落组成及其对环境因子的生理适应能力未发生变化时,增温减雨会导致生物结皮覆盖土壤净碳通量有升高,使其释放的CO2更难被光合固定的CO2取代,进入“C饥饿”的状态。藓结皮净碳固定的土壤温度阈值(13.5℃)显著高于藻结皮(7.3℃)(图5),土壤水分阈值(8.9%)明显低于藻结皮(11.4%)(图6),表明增温减雨的气候变化更可能导致藻结皮覆盖土壤的碳损失,因此未来需要关注野外环境中生物土壤结皮碳通量与环境因子之间复杂的联结关系[6],以明确全球气候变化下生物土壤结皮的碳效应。
4 结 论
藓结皮与藻结皮覆盖土壤碳通量月变化相似,并与土壤温度、水分的月变化同步,其中呼吸速率和光合速率的月变化分别与土壤温度、含水量月变化同步,表现为“先升后降”的趋势,净碳通量则受到了土壤温度、含水量的交互作用,表现为“先升后降再升”的趋势。两类结皮的净碳释放量随浅层含水量的增加而降低,随深层土壤温度的增加而升高,光照强度通过改变土壤、大气温度、湿度间接影响两类结皮覆盖土壤的碳通量。在相同的温度与水分范围内,藓结皮的净碳固定能力比藻结皮更强。藓结皮的净碳固定土壤水分阈值明显低于藻结皮,土壤温度阈值明显高于藻结皮,使其在全球气候变化引起的增温减雨的背景下更不易发生碳损失。未来应结合室内模拟试验长期原位观测生物结皮土壤碳通量与环境因子间的相互作用,以反映生物结皮碳通量与环境因子间的复杂关系。
