白蚁品级分化研究进展
2021-10-10嵇保中刘曙雯姜宏健金明霞
嵇保中,张 磊,刘曙雯,姜宏健,金明霞
(1.南京林业大学林学院, 南方现代林业协同创新中心, 江苏 南京 210037;2. 南京中山陵园管理局, 江苏 南京 210014)
社会性昆虫品级分化是一种多型现象(polyphenism),即具有相同遗传背景的个体在胚后发育中因为基因表达差异从而形成了具有不同特征的个体。由于基因表达的不连续变化,导致个体间形成明确的形态和功能分化类群,这种因表型可塑性形成的非连续性形态分化,也称为品级多型(caste polyphenism)[1]。白蚁品级的记载可远溯至1781年Smeathman的工作[2],但对白蚁品级分化的研究则以Grassi等的工作开始为标志[3]。作为社会性昆虫,白蚁兼具“昆虫”和“社会性”双重属性。与其他不完全变态昆虫类似,白蚁一生经历卵、若虫、成虫3个阶段,形成卵、无翅芽若虫、若虫、有翅成虫等非品级个体。但在特定虫态之下会进一步分化,形成工蚁、兵蚁、生殖蚁等品级个体[2]。从胚后发育的角度,社会性昆虫的品级分化是对胚后发育正常方向的偏离[4]。白蚁的偏离主要发生在若虫期,如工蚁、兵蚁、假工蚁、幼体生殖蚁等均属于若虫偏离型个体。而来源于有翅成虫的原始生殖蚁、成虫型生殖蚁则属于成虫偏离型个体[2, 5]。白蚁品级分化包括群体成员对环境的感知、相关信息的传递、个体对外部信息的生理响应、激素及其调控途径等环节[6]。近年来,表观遗传学与白蚁品级分化的关系逐渐受到关注[7]。笔者从品级分化的类型及其影响因子、个体识别、信息传递、激素调控机理等方面,对白蚁品级分化的研究现状进行综述,以期为白蚁群体结构的深入理解及有效防治提供参考。
1 白蚁品级分化的类型及影响因子
1.1 生殖个体分化
白蚁的生殖分化包括初生生殖蚁、次生生殖蚁和成虫型生殖蚁的分化,其有翅成虫分化的起点与胚后发育途径有关,胚后发育途径为直线式的种类分化且发生较晚,无翅芽若蚁孵化后经过数次蜕皮形成的高龄无翅芽若蚁和末龄无翅芽若蚁(假工蚁),均可发育形成有翅成虫。二岐式途径的种类分化一般较早,在前期依赖性无翅芽若蚁或后期无翅芽若蚁蜕皮时进入若蚁路径,进而发育形成有翅成虫[2]。次生生殖蚁可由两性生殖形成的雌雄个体产生,包括由无翅芽若蚁形成的严格意义上的幼体生殖蚁,工蚁形成的工蚁型幼体生殖蚁(拟工蚁),兵蚁形成的兵蚁型幼体生殖蚁,若蚁形成的若虫型幼体生殖蚁(拟若蚁)[2]。次生生殖蚁的形成与系统进化地位有关。对6科61属199种白蚁的统计分析表明,低等白蚁可产生次生生殖蚁的属占比达61.7%,高等白蚁的占比为13.4%[8]。雌性次生生殖蚁也可由初生生殖蚁通过产雌孤雌生殖产生,这种由原始蚁后通过孤雌生殖形成次生蚁后,使原始蚁后遗传信息得到保真和扩增的途径,称为无性蚁后继承(asexual queen succession,AQS)系统。AQS系统已在南方散白蚁(Reticulitermesvirginicus)、栖北散白蚁(R.speratus)、Embiratermesneotenicus等白蚁中发现[9]。
1.2 非生殖个体分化
白蚁的非生殖分化包括工蚁、假工蚁、兵蚁的分化。工蚁为蚁巢与采食场所完全分开或部分重叠的种类所具有,为适应离巢觅食需要,工蚁的足、体壁高度骨化而强健,分化形成其他品级个体的潜力明显下降[2]。假工蚁是若蚁通过返还式蜕皮或高龄幼蚁通过渐进式蜕皮形成的与幼蚁存在形态区别的个体。兵蚁由无翅芽若蚁或工蚁发育形成,形成兵蚁的无翅芽若蚁或工蚁的虫龄不同,形成的兵蚁大小各异,而不同大小的个体在职能上也存在分化,分别称为大兵和小兵[2]。
1.3 品级分化的影响因子
白蚁品级分化的影响因子包括遗传和环境因素两个方面,前者如品级个体的性别连锁,后者包括温度、营养、群体结构和成熟程度等。群体中原有品级个体对同类品级的分化有抑制作用,但对其他品级个体的分化有促进效果。洞白蚁(Porotermesadamsoni)功能性补充生殖蚁可以阻止新补充生殖蚁形成。在缺乏功能型生殖蚁时,许多幼蚁和若蚁均可发育成为补充生殖蚁。功能性雌蚁对其他雌蚁性成熟的抑制效果比对其他雄蚁的抑制效果更强,但这种抑制效果是短暂的。将生殖蚁移出群体2~4 d后,抑制作用则完全消失[10]。山林原白蚁(Hodotermopsissjostedti)雌雄次生生殖蚁能以性特异性方式调节次生生殖蚁的分化。雌性次生生殖蚁的存在可以抑制4~7龄幼蚁(假工蚁)向雌性次生生殖蚁的分化,而雄性次生生殖蚁则可促进雌性次生生殖蚁的分化[11]。营养改善可促进兵蚁分化。内华达动白蚁(Zootermopsisnevadensis)新建群体兵蚁由最早出现的3龄幼蚁约在8日龄时分化形成前兵蚁进而形成兵蚁,生殖蚁对该3龄幼蚁哺喂养的频次明显多于其他幼蚁[12]。第2堆砂白蚁(Cryptotermessecundus)属于单片巢型白蚁,木料是其食物也是其居所,食物的可利用性是决定个体是否发育为分飞蚁的重要因子,食物数量下降伴随着蚁巢寿命的缩短,巢内个体的发育方向就会从助手蚁(工蚁)改变为分飞蚁[13]。台湾乳白蚁(Coptotermesformosanus)工蚁向兵蚁的分化受温度调节,提高温度促使兵蚁形成的数量更多、分化速率更快[14]。品级分化还受群体成熟程度的影响。来自成熟群体的黑胸散白蚁(Reticulitermeschinensis)工蚁形成成熟替代生殖蚁的历期显著缩短,在工蚁群体中添加兵蚁会延长替代生殖蚁产生历期。来自幼年群体的工蚁,在替代生殖蚁分化前期,取食量呈增加趋势[15]。群体数量也影响品级分化。台湾乳白蚁以5头工蚁建群,不能分化出兵蚁;10头以上工蚁组成的群体可产生兵蚁[16]。遗传基因对品级分化的影响涉及性连锁。栖北散白蚁品级的遗传决定符合单个X-连锁位点、两个等位基因(A、B)的遗传模型,工蚁型、若蚁型替代生殖蚁交配产生的后代中具有wkAB和wkAY基因型的个体发育为工蚁,具有wkAA和wkBY基因型的发育为若蚁[17]。一些白蚁的工蚁和兵蚁有性别倾向。长鼻白蚁属(Schedorhinotermes)和棒鼻白蚁属(Parrhinotermes)的工蚁、兵蚁为雌性,而无刺白蚁属(Anacanthotermes)和角象白蚁属(Cornitermes)的工蚁、兵蚁为雄性[18]。
2 白蚁品级分化的群体内信息传递
个体对群体状况的感知是品级分化的前提,主要包括对群体内其他个体的类别、生理情况等信息的感受与传递,群体内的个体兼有释放者和感受者双重职能。同时,较为密闭的巢内环境使白蚁在背景气味、信息化合物以及感受机制等方面有别于其他开放空间活动的昆虫。
2.1 表皮碳氢化合物
表皮碳氢化合物(cuticular hydrocarbon, CHC)是昆虫上表皮中碳链长度在C20—C50、直链或支链、饱和或不饱和烃类混合物,是昆虫表皮蜡层的主要成分。CHC的基本功能是在虫体表面形成疏水层,减少体内水分散失并防止有害物质(如农药)和病原微生物入侵。CHC是白蚁种间、种内以及群体内个体类型和生理状态识别的重要依据[19]。台湾乳白蚁各品级个体间CHC组分相似,但各组分含量不同,工蚁和兵蚁之间差异最显著。不同季节工蚁和兵蚁均有周期性变化的组分出现,这种周期性变化与长翅生殖蚁的产生有关[20]。内华达动白蚁不同生殖状态的生殖蚁、工蚁和兵蚁间CHC组分有明显差异,其中6,9-二十九二烯烃、6, 9-三十一烷、6,9,17-三十二烷和6,9,17-三十三烷仅蚁王和蚁后体内存在[19]。黑翅土白蚁(Odontotermesformosanus)工蚁、兵蚁、幼蚁、长翅生殖蚁的CHC组分种类相同,但组分含量存在差异。不同龄幼蚁和不同发育阶段工蚁,CHC存在特征性组分含量,CHC分泌随个体发育而改变[21]。北美散白蚁(Reticulitermesflavipes)工蚁、兵蚁、若蚁、替代生殖蚁CHC组成成分不同,用保幼激素类似物(juvenile hormone analogue, JHA)诱导工蚁分化为兵蚁时,在工蚁—前兵蚁和前兵蚁—兵蚁蜕皮前后,CHC组成成分均会发生变化[22]。北美散白蚁蚁后的特异性CHC为正二十一烷,属于蚁后识别信息素,应用玻璃假体生测可激发工蚁的行为反应[23]。
2.2 挥发性化合物
高山象白蚁(Nasutitermestakasagoensis)蚁后、工蚁、兵蚁、有翅成虫和卵挥发物中特异性化合物的种类:兵蚁有7种,有翅成虫、卵、蚁后各1种。蚁后特异性挥发物为苯乙醇[24],工蚁挥发物中无特异性化合物。栖北散白蚁雌性替代生殖蚁能产生挥发性抑制信息素,抑制雌性替代生殖蚁的分化。信息素的活性成分为n-丁基-n-丁酯和2-甲基-1-丁醇。用这两种成分配制的人工信息素具有与替代生殖蚁活体类似的效果。卵也能产生上述两种信息素成分,用于吸引工蚁和抑制生殖蚁分化[25]。n-丁基-n-丁酯和2-甲基-1-丁醇还具有抗菌活性,能显著降低栖北散白蚁卵表寄生真菌的孢子萌发率和菌丝生长。推测白蚁最初可能利用这些成分保护卵和蚁后,此后逐渐演变为蚁后的生殖力信号物质[26]。北美散白蚁兵蚁头部提取物中含有γ-杜松烯和γ-cadinenal,分别是兵蚁分化的促进或抑制成分[27]。栖北散白蚁卵表的抗菌蛋白溶菌酶属于白蚁卵识别信息素,对革兰氏阳性细菌有抗菌活性,对工蚁卵搬运和梳理行为有强烈激发效果[28]。简单原鼻白蚁(Prorhinotermessimplex)、桑特散白蚁(Reticulitermessantonensis)、黄颈木白蚁(Kalotermesflavicollis)等功能性生殖蚁体表水洗物中含有与年龄、生殖状态有关的性特异性蛋白,可能参与品级识别和调节[29]。
2.3 信息化合物的传递途径
一般认为大分子挥发性差的CHC通过直接接触传递,小分子挥发性强的信息化合物则可通过分子挥发间接传递。但有时两者缺乏明确的界限。象白蚁(Nasutitermeslujae)兵蚁可以抑制或延滞新兵蚁的产生,这种抑制效果来源于兵蚁颚腺分泌的引物信息素,必须通过直接接触才起作用[30]。台湾乳白蚁工蚁必须与兵蚁或前兵蚁发生直接虫体接触才能产生抑制效果[31]。山林原白蚁生殖蚁抑制工蚁向次生生殖蚁分化也是通过虫体直接接触介导的[11]。用人工合成的蚁后信息素在供试工蚁群体与信息素挥发源分开的情况下,对栖北散白蚁工蚁群体的生殖蚁分化有显著的抑制效果[25]。Korb[32]认为群体规模较小的单片巢型白蚁,群体内信息交流以接触为主;而群体规模较大的离散巢型白蚁,虫体接触无法保证,则需要挥发性信息素。北美散白蚁的蚁后信息素正二十一烷,也能激发工蚁振动和触角反应[23]。上述说明白蚁群体内的信息化合物兼具引物信息素和释放信息素双重效应。
3 白蚁品级分化的激素调节
白蚁品级分化是包含蜕皮变态的胚后发育过程,具有其他昆虫蜕皮变态类似的脑激素(brain hormone, BH)、蜕皮激素(moulting hormone, MH)、保幼激素(juvenile hormone, JH)调控过程。但品级分化是响应多种环境因子的动态过程,其激素调节更加细致复杂[5]。正常发育和多态分化因子作用于JH-MH偶联体,而品级形成和器官分化(上颚、卵巢、翅、性特征)则是对血淋巴内激素动态的应答[33]。
3.1 JH的作用
一般认为,蜕皮间期白蚁的JH水平决定蜕皮分化的结果[34],这与其他昆虫MH启动蜕皮、JH决定蜕皮后个体发育方向类似。关于血淋巴JH水平与白蚁品级决定的关系,Nijhout等[35]、Cornette等[36]、Korb[37]先后提出不同的理论模型。Nijhout的模型表明黄颈木白蚁胚后发育至少存在3个JH敏感期:第1个位于龄期开始阶段,决定生殖系统发育;第2个位于龄期早期,决定翅、复眼、表皮等非生殖性成虫特征;第3个位于龄期后半段,决定兵蚁特征。如果在3个敏感期JH均处于高水平,个体发育为兵蚁;如果均处于低水平,则发育为有翅成虫。如果龄期开始时JH水平低,完成生殖系统发育,在第2个敏感期JH水平升高,抑制了非生殖成虫特征,则发育为次生生殖蚁。如果前两个敏感期JH水平高,抑制了成虫特征,第3个敏感期JH水平低,抑制了兵蚁特征,则进行静态蜕皮形成假工蚁。假工蚁形成替代生殖蚁的蜕皮间期最短,似乎处于兵蚁分化决定敏感期之前(图1)[35]。
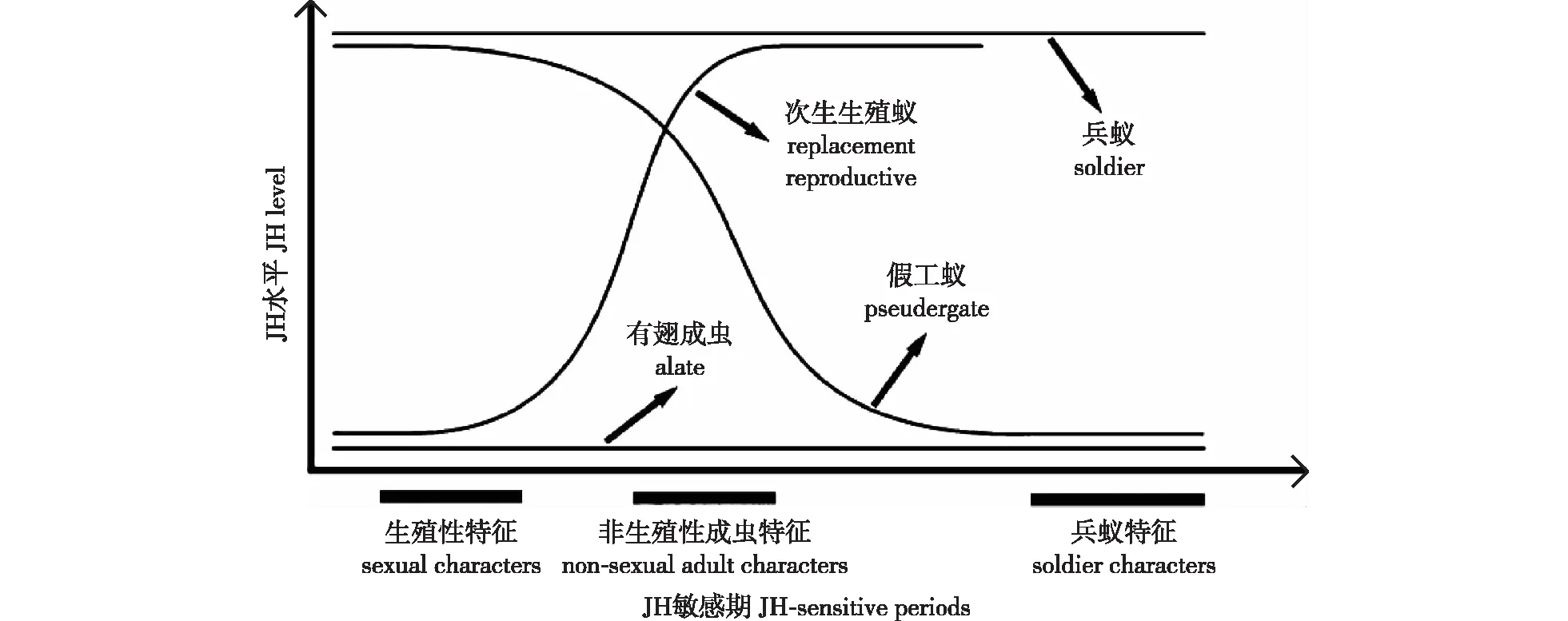
图1 黄颈木白蚁品级决定与JH水平的关系[35]Fig.1 The relationship between Kalotermes flavicollis caste determination and JH level
Cornette等[36]的模型表明,山林原白蚁无翅路线(兵蚁、假工蚁等)发育过程中,在蜕皮前血淋巴JH水平均有一个峰值。如果蜕皮间期JH维持高水平,假工蚁蜕皮形成前兵蚁,蜕皮前血淋巴JH水平升高,蜕皮后短暂下降随即恢复高水平直至蜕皮形成兵蚁。如果蜕皮间期JH处于低水平,则启动静态蜕皮形成假工蚁,蜕皮前JH峰值显著以维持形态不变。Korb的模型[37]表明,第2堆砂白蚁无翅芽若蚁—若蚁蜕皮后血淋巴JH水平显著下降。若虫期JH水平总体呈下降趋势,每龄若蚁蜕皮后有一个短暂升高,随若蚁龄数增加,升高的幅度逐渐减弱。有翅成虫蜕皮前JH水平升高,成虫形成后显著升高。次生生殖蚁分化前JH水平显著下降,蜕皮后升至较高水平。前兵蚁分化前有一个JH峰,分化后下降至低水平进入兵蚁分化[38]。Korb等[39]的模型涉及JH和社会性因子的互作(图2),在缺乏兵蚁或生殖蚁情况下,高水平JH导致工蚁分化形成兵蚁,低水平JH则促进分化形成次生生殖蚁(图2A);在兵蚁、生殖蚁存在的情况下,高、低水平JH分别导致返还式蜕皮和渐进式蜕皮(图2B)。

图2 低等白蚁的JH和社会性因子互作的品级决定模型[39]Fig.2 The caste decision model of the interaction by JH and social factors of lower termites
血淋巴JH水平是合成和分解的动态过程。栖北散白蚁工蚁、若蚁、替代生殖蚁体内存在去甲哈尔满,对JH环氧化水解酶活性具有低浓度促进、高浓度抑制的效果,可通过调节JH水平进而影响品级分化[40]。北美散白蚁脂肪体细胞内储存蛋白可与JH结合,通过对JH的封存抑制血淋巴JH水平,影响品级分化[41]。总体上,白蚁体内JH的作用与其他昆虫类似:在幼蚁体内,具有维持原有状态的作用。在成蚁体内,则具有促性腺作用。除上述JH水平与品级决定之间的关系外,还有一些品级分化现象与JH水平有关。①JH可以调节CHC成分:用JHA诱导北美散白蚁工蚁分化为兵蚁时,在工蚁—前兵蚁和前兵蚁—兵蚁蜕皮前后,CHC组成成分均会发生变化[22]。JH对生殖特异性CHC合成与分泌的促进,可能与促进生殖系统发育有关[42]。②群体数量和群体中兵蚁对兵蚁分化的调节作用,是通过影响工蚁JH水平而实现的。将台湾乳白蚁工蚁和不同数量的兵蚁组成实验群体,一段时间后,低兵蚁组工蚁体内JH水平更高。随着兵蚁比例下降新产生的兵蚁数量增加,工蚁体内的JH水平与群体内兵蚁数量呈显著正相关[43]。台湾乳白蚁10头以上工蚁组成的群体可产生兵蚁。当建群数量从50增加到1 000头时,工蚁JH水平提高。而兵蚁的存在则降低了群体数量对工蚁JH水平的影响[16]。③温度对品级分化的影响可能也与JH水平有关。对北美散白蚁的研究表明,兵蚁抑制性储存蛋白的丰度和储存蛋白对JH的封存能力均呈显著的温度依赖性效果[44]。
3.2 MH的作用
白蚁品级分化是感受环境信息后,BH、MH、JH协同作用的结果[6]。Okot-Kotber[45]报道迈克尔森大白蚁(Macrotermesmichaelseni)胚后发育过程中,从1龄无翅芽若蚁直到1~5龄若蚁,其前胸腺体积持续增加,成虫蜕皮后前胸腺消失。但这种体积变化与生殖蚁特征分化的关系却不清楚。迈克尔森大白蚁体质量较轻的3龄雌性无翅芽若蚁蜕皮形成小工蚁,较重的个体形成小前兵蚁,进而形成小兵蚁。蜕皮变态期间皮层脱离、皮细胞生长、新表皮沉积、蜕皮等表皮变化,与MH水平提高显著相关,在这些变化中体质量较轻的幼蚁个体总是滞后2 d。在前兵蚁向兵蚁转变中,也有两个明显的MH峰[46]。将北美散白蚁工蚁隔离后,血淋巴内JH、MH水平均升高,认为可能是诱导工蚁分化的启动效应。检测发现MH主要为20-羟基蜕皮酮(20E),推测20E对白蚁形态发生更重要,而蜕皮酮(E)对促进生殖系统发育更重要[47]。用早熟素Ⅰ处理第2堆砂白蚁工蚁群体,产生的兵蚁比对照群体更少。处理组会产生工蚁-若蚁中间体、末期若蚁和有翅成虫。说明早熟素Ⅰ处理可抑制兵蚁发育、促进成虫性状发育[48]。将细腰湿木白蚁(Zootermopsisangusticollis)成虫脱翅后按雌雄配对、雌雌配对、雌蚁单独饲养处理,雌雄配对组的雌虫,第5天 MH滴度提高,并保持在高水平。雌蚁独处和雌雌配对处理的雌蚁,MH滴度下降并保持低水平至少15 d,到30 d时显著提高[49]。第2堆砂白蚁胚后发育伴随着JH、MH水平的波动,若蚁和初生生殖蚁体内MH含量一般很低(小于50 pg/μL),但在3、4龄若虫和有翅成虫的某些个体内MH滴度升高[50]。
3.3 JH、MH的下游信号
JH、MH的下游信号主要包括:①基因组信号。内华达动白蚁JH受体基因(Met)同源基因(ZnMet)前兵蚁蜕皮后在头部的表达增加。在前兵蚁蜕皮前用ZnMet双链RNA(dsRNA)注射可以降低这种高表达。处理不会引起前兵蚁形态变化,但会使兵蚁上颚更短,头壳更小[51]。内华达动白蚁前兵蚁蜕皮后,MH受体基因(EcR)即刻高表达。前兵蚁早期敲除EcR,可抑制前兵蚁蜕皮形成兵蚁。在前兵蚁中期敲除,则影响酪氨酸代谢基因表达,抑制兵蚁特异性表皮鞣化[52]。对Met进行RNA干涉(RNAi),可抑制内华达动白蚁前兵蚁蜕皮。敲除Met可抑制20E合成基因的表达。对JHA处理个体中的这些基因进行RNAi,20E信号和核受体基因(HR39/FTZ-F1b)的敲除导致新蜕皮个体出现正常的工蚁表型[53]。与若蚁和工蚁比较,JH信号基因Met、kr-h1、Br-c在栖北散白蚁次生生殖蚁腹部高表达。对Met进行RNAi导致新蜕皮次生生殖蚁卵黄原蛋白基因表达的抑制[54]。②凋亡决定信号。昆虫变态发育过程中,MH会激活遗传级联系统调控的凋亡信号通路,诱导一些组织细胞的凋亡[55]。尖唇散白蚁(Reticulitermesaculabialis)生殖蚁、工蚁、兵蚁生殖力的显著差异受卵子发生早期的细胞凋亡控制[56]。圆唇散白蚁(R.labralis)雄性工蚁分为有精子工蚁(WS)和无精子工蚁(WN)。WN和兵蚁的精子发生止于精原细胞期。与高龄WN和兵蚁相比,高龄WS和生殖蚁的精原细胞凋亡率更低,精母细胞和精子细胞则很少凋亡。说明雄性工蚁的生殖分化受精子发生早期细胞凋亡的控制[57]。③储存蛋白信号。用JH处理北美散白蚁工蚁,4种编码血淋巴蛋白的基因表达显著增强,两种卵黄原蛋白和两种储存蛋白的基因在JH处理后表达显著提高[58]。用JHA蚊蝇醚处理山林原白蚁假工蚁,处理后3 d脂肪体开始发育,约在蜕皮进入前兵蚁时期的前4天充满整个腹腔,并维持这种状态直到蜕皮形成兵蚁。脂肪体发育伴随着细胞质内蛋白质颗粒的积累,但这些颗粒在蜕皮进入前兵蚁前几天消失。这些贮存蛋白消耗的时间与表皮生长的时间窗口一致[59]。④表观遗传信号。De Loof等[60]提出表观-内分泌学(epi-endocrinology)的概念,认为JH、MH除了经典的受体作用模式,在控制关键生理过程方面还存在表观遗传学途径。Lo等[61]首先在澳洲乳白蚁(Coptotermeslacteus)甲基化特异性扩增片段长度多态性检验中发现了DNA甲基化证据,但不同品级个体DNA甲基化水平没有显著差异。在北美散白蚁和台湾乳白蚁表达的基因中,也存在高水平DNA甲基化特征。DNA甲基化喜好以广泛表达的形态基因作为靶标,其功能联系与其他存在功能性DNA甲基化系统的无脊椎动物类似[7]。山林原白蚁DNA甲基转移酶(dnmt1、dnmt2、dnmt3)和甲基-CpG结合域(mbd)EST文库中存在上述4种基因,而在栖北散白蚁和高山象白蚁文库中缺乏dnmt1。在上述3种白蚁中,大部分编码序列CpG含量低于1,含量分布呈双峰型,说明存在DNA甲基化[62]。应用转录组测序技术,从栖北散白蚁蚁王、蚁后中鉴定出74个表观修饰基因,其中15个蚁王特异性表达,52个年龄依赖性蚁王、蚁后差异性表达。DNA甲基转移酶3在蚁王精巢和脂肪体特异性表达,一些组蛋白修饰基因在蚁王精巢和蚁后卵巢内显著表达。说明表观遗传修饰在白蚁蚁王、蚁后的精子和卵子发生过程中发挥重要作用[63]。在内华达动白蚁新建巢群体中,最成熟的3龄幼蚁(1号幼蚁)分化形成前兵蚁。转录组测序检测到1号幼蚁体内7个表观遗传相关基因表达水平显著增加。对这7个基因进行RNAi降低了JH合成、结合、信号基因的表达水平,其中对3个组蛋白修饰基因的RNAi延长了前兵蚁蜕皮期。这说明表观遗传有关基因不会直接影响白蚁兵蚁分化,但有些组蛋白修饰基因对蜕皮间期有影响,可能是对兵蚁分化期间JH作用的调节所导致[64]。从栖北散白蚁工蚁、若蚁以及台湾乳白蚁工蚁体内分别鉴定出97、101、114种小RNA(miRNAs),栖北散白蚁工蚁上调的miRNAs中,miR-13b-3p表达水平最高;若蚁下调的miRNAs中,miR-305-5p表达量最高。台湾乳白蚁工蚁上调的miRNAs中,miR-11-3p表达水平最高。两种白蚁的工蚁似乎使用不同的小RNA调节转录因子基因[65]。
3.4 JH、MH的上游信号
JH、MH的上游信号主要包括:①脑激素。北美散白蚁抑咽侧体神经肽(allatostatins, ASTs)在工蚁、若蚁、兵蚁、次生生殖蚁的心侧体、咽侧体、支配这些器官的脑侧区和中区分泌细胞以及其他脑神经细胞中均有免疫反应[66]。多种来自蜚蠊(Diplopteraspp.)的ASTs能抑制北美散白蚁咽侧体JH的合成,JH合成速率较低的咽侧体对ASTs的敏感性更强。从北美散白蚁脑提取物中也鉴定出多种蜚蠊ASTs[67]。从北美散白蚁和太平洋折翅蠊(Diplopterapunctata)脑提取物中获得的ASTs-7在咽侧体内含量丰富,不但能抑制两种昆虫咽侧体JH的合成,还具有神经和肌肉调节活性[68]。②胰岛素信号。山林原白蚁兵蚁分化期间胰岛素受体相关基因上调,在上颚表皮组织强烈表达,对受体基因的RNAi可扰乱兵蚁特异性形态分化[69]。③生物胺信号。内华达动白蚁新建巢群体兵蚁由最成熟的3龄幼蚁(1号幼蚁)分化形成,0~1日龄1号幼蚁多巴胺(DA)合成基因表达水平很高,3日龄1号幼蚁脑内DA水平显著高于0日龄的。DA可调节生殖蚁交哺活动的频率,进而调节兵蚁分化[70]。山林原白蚁兵蚁酪胺(TA)/章鱼胺(OA)神经元支配区域扩大,脑和咽下神经节内TA水平以及脑内OA水平比假工蚁高,TA/OA水平提高可以诱导更强的攻击和防卫行为[71]。④营养信号。内华达动白蚁新建巢群体之间载脂蛋白(ZnNLaz1)基因最具表达差异性,敲除ZnNLaz1显著抑制幼蚁与蚁后间的交哺活动,降低兵蚁的分化率。该蛋白与通过交哺从蚁后获得的社会性信号的综合作用有关[72]。对3龄幼蚁进行整体RNA测序,发现幼蚁营养敏感性信号途径上调,将要发育为工蚁的幼蚁则表现为代谢途径上调[73]。⑤生长因子信号。对内华达动白蚁转化生长因子——β信号(TGFβ)有关的2个基因进行RNAi,诱导的前兵蚁出现兵蚁样表型和类似于兵蚁的攻击行为,可能是由于MH信号基因表达水平下降,这些前兵蚁不会蜕皮进入兵蚁,说明TGFβ信号具有兵蚁分化期间调节JH和20E信号的功能[74]。
4 结 语
白蚁品级分化是包含昆虫蜕皮变态和社会环境因子影响的复杂过程,已有研究虽然取得了相关进展,但许多机制仍未明确。在纷繁复杂的影响白蚁品级分化的环境因子中,营养可能具有根本性,其他因子可能只是营养因子在不同情况下的表现形式。节肢动物的发育分化主要受营养刺激控制,其他环境因子也很重要,如气候和社会性的相互作用、遗传效应等。从进化的角度看,品级的遗传决定有费解之处。因为它使个体根据环境变化调节品级比例的能力下降,而这种下降对个体并无益处。改变发育方向的原因很可能是由于特定发育时期内分泌系统对营养刺激反应的变化。当幼虫随着食物摄入生长至超过体型大小的临界值时,则启动影响发育的生理变化,幼虫转入生殖特异性发育途径[75]。群体内不同类型个体生理状态信息的传递可能是多途径综合协调的过程。那些生活在密封巢内、群体数量较少的木栖白蚁,主要应用CHC等作为信号。而拥有更大规模群体和不太密封蚁巢的种类,挥发性化合物则可能发挥主要作用[32]。在较为密闭的蚁巢内,携带个体生理信息的挥发性成分可能以弥散的方式形成背景性气味。这些挥发性成分既可引发生理反应,又可诱发行为反应。如内华达动白蚁蚁王、蚁后的缺失可造成群体内头部触碰行为的增加[76]。黄颈木白蚁蚁王在群体内活动和接触范围最高,作为蚁后和群体其他成员社会性相互作用的调节者,在群体社会结构和动态发育过程中发挥基本调节作用[77]。这些行为对信息的传递和放大作用,值得关注。虽然对白蚁BH和MH研究较少,但白蚁品级分化无疑受BH、MH、JH的综合调控,这符合白蚁的昆虫属性和品级分化的变态发育属性。其中,与环境因子的营养属性对应的营养因子信号以及生物胺信号等在品级分化中的作用、与品级分化有关的表观遗传学途径等,均需进一步研究。虽然对信息素和激素的偶联机制目前较少研究,但在破解白蚁品级分化机理方面却尤为重要。
