1 株产甘露聚糖酶嗜热真菌的鉴定、酶学性质表征及转录组学分析
2021-09-28谈苏慧卢海强张莉娟田洪涛谷新晰
谈苏慧,卢海强,陈 伟,张莉娟,田洪涛,谷新晰*
(河北农业大学食品科技学院,河北 保定 071000)
β-甘露聚糖酶(β-1,4-D-mannanase,EC 3.2.1.78)是内切水解酶,可随机水解甘露聚糖分子内部的β-1,4糖苷键,将甘露聚糖降解为葡萄糖、甘露寡糖等低聚糖[1-2]。β-甘露聚糖酶在食品行业应用前景广阔,其可降解角豆胶[3]、瓜儿豆胶[4]、魔芋胶[5]等富含甘露聚糖的植物胶生产甘露寡糖。甘露寡糖具有促进肠道有益菌生长、调节肠道黏膜免疫、改善肠道菌群环境等作用,是一种新型的优质益生元[6-7]。β-甘露聚糖酶还可用于降低咖啡、巧克力及可可液等的黏性以利于其进一步加工[8]。Suryawanshi等[9]发现甘露聚糖酶的多酶系统固定化后,β-甘露聚糖酶可在澄清苹果汁、猕猴桃汁、橙汁、桃汁等果汁的同时,增加其还原糖含量。Singh等[10]的研究结果表明β-甘露聚糖酶可高效去除含甘露聚糖食物引起的污渍。崔婷婷等[11]研究发现在面团中添加β-甘露聚糖酶处理15 min的体积分数为2.0%魔芋葡甘聚糖,能够显著抑制二硫键的断裂和二级结构的变化,防止面团的水分散失,维持面团的拉伸能力。Comfort等[12]报道的甘露聚糖酶多为最适反应温度在50~60 ℃之间的中低温酶,无法满足工业生产中较高的温度要求。因此,热稳定性较好的嗜热甘露聚糖酶逐渐成为新的研究热点[13]。
嗜热真菌能在45 ℃以上正常生长,且在19 ℃无法生长[14],嗜热真菌分泌的酶类大部分为嗜热酶[15]。嗜热甘露聚糖酶具有热稳定性好、可提高反应的催化效率、高温时底物黏度低、促进酶与底物的结合、具有催化等优势[16]。迄今为止,获得的近30 种嗜热甘露聚糖酶仅有少部分来源于嗜热真菌[17],如来自黑曲霉Aspergillus nigerBK01的酶Man BK[18]、来自小巢状曲霉Aspergillus nidulansXZ7的酶Man5XZ7[19]和来自篮状菌Talaromyces leycettanusJCM12802的酶Man5A[16]。因此,分离鉴定嗜热真菌并获得嗜热甘露聚糖酶,对提高微生物资源利用率以及解决甘露聚糖酶工业应用受限等问题有较高的理论价值和指导意义。
碳水化合物活性酶(carbohydrate-active enzymes,CAZymes)是参与复杂碳水化合物合成或分解蛋白酶的统称,具有降解、修饰及生成糖苷键的功能[20]。根据氨基酸序列相似性、蛋白结构及催化功能的不同,主要分为5 类催化酶及一类非催化模块,催化酶分别为碳水化合物酯酶(carbohydrate esterases,CEs)、糖苷水解酶(glycoside hydrolases,GHs)、糖基转移酶(glycosyltransferases,GTs)、多糖裂解酶(polysaccharide lyases,PLs)、辅助氧化还原酶(auxiliary activities,AAs)、非催化模块即碳水化合物结合模块(carbohydrate-binding modules,CBMs)[21]。近几年,以转录组学测序技术为基础,探究真菌编码碳水化合物酶基因的数量以及种类,并以此为指标筛选高效产碳水化合物酶的菌株,成为分离筛选优良产酶菌株新依据。
本研究以大曲中分离、筛选的嗜热真菌GZFH7为研究对象,对该菌株进行生物学鉴定,并对其诱导的嗜热甘露聚糖酶的酶学性质进行研究。同时,利用转录组学探究该菌株产碳水化合物活性酶的性能,为进一步开发利用该菌株提供理论依据。
1 材料与方法
1.1 材料与试剂
菌株GZFH7筛选自贵州酱香型大曲,并由本实验室于4 ℃保存;马铃薯葡萄糖琼脂(potato dextrose agar,PDA)培养基、马铃薯葡萄糖肉汤(potato dextrose broth,PDB)培养基 上海博微生物科技有限公司;真菌DNA提取试剂盒、真菌通用引物ITS1、ITS4、PCR mix、ddH2O 北京博迈德基因技术有限公司;乳酸酚棉蓝染色液 青岛海博生物技术有限公司;Oligo(dT)磁珠 无锡百迈格生物科技有限公司。。
基础盐溶液:(NH4)2SO4、KH2PO4、MgSO4·7H2O、FeSO4·7H2O、CaCl2的质量浓度分别为1.00、1.00、0.01、5.00、0.20 g/L。
魔芋发酵培养基:基础盐溶液中加入魔芋胶、酵母浸粉质量浓度均为3.00 g/L;多碳源发酵培养基:基础盐溶液中加入魔芋胶、麸皮、酒糟质量浓度均为1.00 g/L。
1.2 仪器与设备
MJX-250B-Z恒温霉菌培养箱 上海博讯实业有限公司医疗设备厂;MODEL YS100电子显微镜 日本Nikon公司;TU-1810PC紫外-可见光分光光度计 北京普析通用仪器有限公司;NDJ-5S数字式黏度计 上海佑科仪器仪表有限公司;CR-G立式高速低温离心机日本Hitachi公司;Biometra Tprofessional聚合酶链式反应(polymerase chain reaction,PCR)仪 德国Biometra公司。
1.3 方法
1.3.1 菌株筛选
取研碎的大曲样品10.0 g于100 mL无菌水中,于37 ℃、150 r/min摇床振荡2 h。静置后取上清液,用无菌水适当稀释,涂布于PDA培养基,45 ℃倒置培养,挑取单菌落点植于PDA培养基上,分离、纯化得到的单一菌株保藏于麸皮管中,4 ℃存放、备用。
1.3.2 粗酶液的制备
将保藏于麸皮管的GZFH7接种于PDB活化培养基中,45 ℃摇床培养1 d。按1%的接菌量从活化培养基中吸取菌液加入魔芋发酵培养基中,45 ℃摇床培养3 d。发酵液于4 ℃、8 000 r/min离心15 min,收集上清液,即为粗酶液,4 ℃备用制备好的粗酶液。
1.3.3 菌株的鉴定
1.3.3.1 菌株嗜热性鉴定
将菌株GZFH7点植于PDA平板中心处,分别于19、45 ℃倒置培养,观察并记录菌株的生长情况。
1.3.3.2 菌落形态特征观察
采用点植法将GZFH7接种于PDA平板中,45 ℃倒置培养,观察菌落的形状、大小、颜色及生长状况等情况并进行记录,对照真菌鉴定手册[22],初步确定菌株的种属地位。
采用棉蓝染色法[23]进行制片观察:于洁净的载玻片中央滴一滴乳酸酚棉蓝染色液,用接种环挑取GZFH7平板中的少量带有孢子的菌丝置于染液中,细心地将菌丝挑散开,加盖玻片,在电子显微镜40 倍镜观察霉菌形态,记录菌株菌丝孢子、孢子囊以及孢子梗的形态特征。
1.3.3.3 菌株的分子鉴定
采用CTAB法提取菌株GZFH7的基因组DNA;获得的基因组DNA经ITS序列PCR扩增后,得到的PCR产物由华大基因科技服务有限公司进行测序。将获得的测序结果提交NCBI数据库(http://www.ncbi.nlm.gov/blast/)进行BLAST同源性分析,采用MEGA 7.0软件构建系统进化树并分析。
1.3.4 生长曲线的测定
采用菌落直线生长法测定菌株GZFH7的生长曲线[23],将菌株GZFH7点植于PDA平板中心处,45 ℃倒置培养,分别在6、11、23、25、29、36、48、54、56 h(菌落长满整个平板)测其菌落直径,绘制菌株的生长曲线。
1.3.5 甘露聚糖酶活力的测定
参考Miller[24]的3,5-二硝基水杨酸(dinitrosalicylic acid,DNS)方法。取100 µL适当稀释酶液和900 µL角豆胶(底物由pH 5.0、50 mmol/L磷酸氢二钠-柠檬酸缓冲液配制、体积分数0.5%),以甘露聚糖酶最适反应温度(1.3.6.1节方法)进行水浴反应10 min后,加入1.5 mL DNS终止反应。对照则在加入1.5 mL DNS后,补加100 µL稀释酶液。沸水浴5 min后冷却至室温,在540 nm波长处测定吸光度。
酶活力单位(U):以每分钟生成1 µmol甘露糖所需的酶量定义为1 U。
1.3.6 酶学性质分析
1.3.6.1 最适反应温度的测定
酶液与底物在不同温度(20、30、40、50、60、70、80、90 ℃)进行反应,按1.3.5节的方法测定酶活力,以最高酶活力为100%,测定酶的最适反应温度。
1.3.6.2 最适反应pH值的测定
在最适反应温度,酶液与不同pH值(2、3、4、5、6、7、8、9、10、11、12)的底物进行反应,按照1.3.5节方法测定酶活力,以最高酶活力为100%,测定酶的最适反应pH值。
1.3.6.3 温度稳定性的测定
在最适反应pH值,按照1.3.6.1节的方法测酶液在最适反应温度,及最适反应温度浮动10 ℃处理不同时间(0、2、5、10、20、30 min和60 min)的酶活力,以最高酶活力为100%,研究甘露聚糖酶的热稳定性。
1.3.6.4 pH值稳定性的测定
用不同pH值(2、3、4、5、6、7、8、9、10、11、12)的缓冲液稀释酶液后,冰浴1 h。最适反应温度按1.3.5节的方法测定酶活力,以最高酶活力为100%,研究甘露聚糖酶的pH值稳定性。
1.3.7 酶解产物黏度的测定
粗酶液与体积分数1%魔芋胶底物1∶2(V/V),70 ℃水浴反应1 h后,迅速冷却至室温,测其黏度。通过比较灭活酶液与正常酶液反应产物的黏度差异,反映甘露聚糖酶降解甘露聚糖酶的能力大小。
1.3.8 平均聚合度(mean degree of polymerization,mDP)的测定及计算
聚合度是指组成寡糖的单糖单位的个数,mDP越接近于1则底物水解越彻底。直接还原糖与总还原糖参照Somogyi等[25]的方法测定,mDP按下式计算:
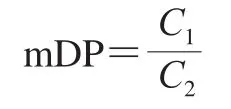
式中:C1、C2分别为酶解液中总糖和还原糖的质量浓度/(mg/mL)。
1.3.9 菌株GZFH7的碳源谱测定分析
不同碳源固体培养基的配制参照Hüttner等[26]的方法,将菌株点分别植于含有不同单一碳源(果胶、纤维素、木聚糖、几丁质、可溶性淀粉、壳聚糖、魔芋精粉)的固体培养基上,相同条件培养相同时间,以PDA平板作为对照,观察菌株GZFH7的生长状况。
1.3.10 菌株GZFH7的转录组学分析
菌株GZFH7接入多碳源发酵培养基,45 ℃摇床培养3 d后收集菌体。采用Trizol法提取总RNA,检测其浓度及纯度,完整性采用RNA专用琼脂糖电泳或者Agilent 2100 Bioanalyzer进行测定、分析。通过Oligo(dT)磁珠富集总RNA中带有polyA结构的mRNA,采用离子打断的方式,将RNA打断约长度为300 bp的片段,并以此为模板构建表达文库。
采用PCR扩增进行文库片段富集,之后根据片段长度进行文库选择,文库长度在450 bp。通过Agilent 2100 Bioanalyzer对文库进行质检,再对文库总浓度及有效浓度进行检测。根据文库的有效浓度及文库所需数据量,将含有不同Index序列的文库按比例进行混合。混合文库统一稀释到2 nmol/L通过碱变性,形成单链文库。
采用第2代测序技术,基于Illumina HiSeq测序平台,对这些文库进行双末端测序,获得长度约380 bp测序reads。对获得的原始测序序列进行分析,去除带接头、长度小于50 bp、序列平均质量低于Q20的Reads。对得到的高质量序列进行从头拼接得到转录本序列。对转录本进行聚类,挑选最长的转录本作为Unigene,最后用Unigene进行后续无参转录组分析,并依据CAZymes数据库(http://www.cazy.org/)分类标准,对测序数据进行统计分析。
1.4 数据分析及处理
每次实验重复测定3 次,利用Excel和SPSS19.0软件对测定结果进行统计分析,数据结果以±s表示。
2 结果与分析
2.1 菌株鉴定
在19 ℃培养3 d,菌株GZFH7几乎不生长。45 ℃培养3 d后,该菌株生长旺盛,菌丝如图1A所示,可铺满整个平皿。生长初期菌落呈白色绒毛状,后期变成灰褐色厚毡状,且菌落表面粗糙。如图1B所示,显微镜可观察到菌丝有较多分支,且匍匐丝彼此有间隔,囊轴呈球形,孢囊孢子的形状有球形,椭圆形以及其他形状。对菌株GZFH7的ITS PCR产物测序结果进行BLAST比对,结果如图1C所示。菌株GZFH7与微小根毛霉(Rhizomucor pusillusT3B)的相似度为100%,并且位于进化树的同一分支中,亲缘关系相近,因此判断该菌为微小根毛霉,命名为R.pusillusGZFH7。
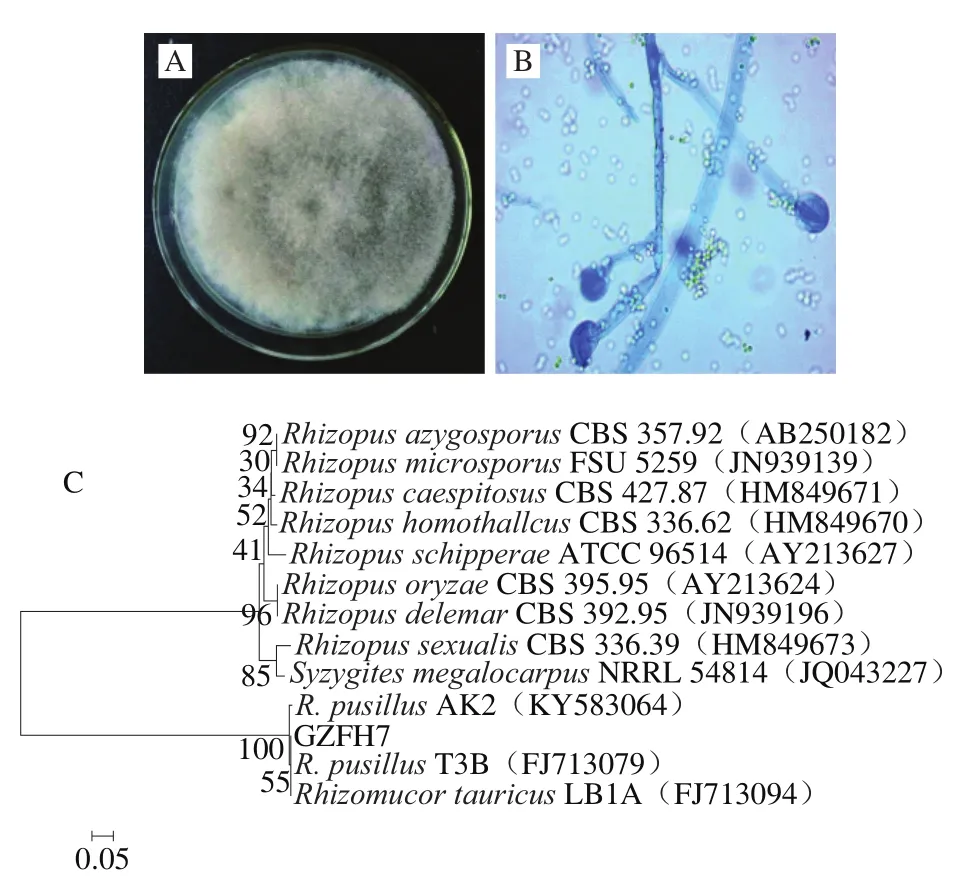
图1 菌株GZFH7菌落特征(A)、微观镜像(B)及系统发育树(C)Fig.1 Colony characteristics (A), microscopic image (B), and phylogenetic tree (C) of strain GZFH7
2.2 菌株生长和产酶曲线的测定
如图2所示,0~10 h为该菌株的生长停滞期,10~54 h之间菌株迅速生长,生物量迅速增加,此阶段为快速生长期,此后随培养时间延长,菌株的生物量趋于平稳,即进入稳定期。通过对GZFH7甘露聚糖酶活力测定分析,发现该菌在迅速生长期开始进行产酶,其产酶量随菌株自身的生长而增加。直至稳定期,甘露聚糖酶产酶量达到稳定值9.5 U/mL,之后随发酵时间延长,产酶量几乎不再增加。因此,GZFH7菌株在45 ℃生长迅速,能够短周期发酵生产甘露聚糖酶。

图2 菌株GZFH7生长及产酶曲线Fig.2 Growth curve and enzyme production curve of strain GZFH7
2.3 酶学性质分析
由图3A可知,菌株GZFH7诱导甘露聚糖酶的最适反应温度为70 ℃,属于嗜热酶。在50~80 ℃范围内,相对酶活力均可达到50%以上。如图3B所示,甘露聚糖酶在60 ℃热处理1 h,其相对酶活力在90%以上;70 ℃处理1 h,酶活力基本保持不变;当该酶在80 ℃处理30 min,能保持高于50%的酶活力,且在处理1 h后,依然能够维持大约50%的酶活力。因此,菌株GZFH7所产的甘露聚糖酶具有较好的热稳定性。如图3C所示,甘露聚糖酶的最适反应pH值为5.0,底物pH 4.0~6.0范围内时,能够维持50%以上的酶活力,具有一定的pH值适应性。如图3D所示,甘露聚糖酶在pH 2.0~12.0范围内均表现一定的酶活力,在酸性范围内(2.0<pH<6.0)酶活力较高,能够维持70%以上的酶活力。酶活力随pH值从5.0开始增加而降低。

图3 甘露聚糖酶的酶学性质Fig.3 Enzymatic properties of mannanase

2.4 酶解产物黏度及mDP计算
经酶水解后,魔芋胶溶液的黏度由125 mPa·s降低为74 mPa·s,该结果表明菌株GZFH7产生的甘露聚糖酶可有效降解魔芋胶,使其由黏度较高的甘露聚糖降解为黏度较低的甘露低聚糖。测得酶解产物的直接还原糖质量浓度为0.39 mg/mL,总还原糖质量浓度为0.37 mg/mL,mDP为0.96,接近于1。该结果表明由菌株GZFH7产生的甘露聚糖酶可较彻底的降解魔芋胶,酶的水解能力较高。
2.5 菌株GZFH7的碳源谱分析
对菌株的碳源谱进行探究,根据其在不同碳源平板上的生长状况,可对该菌株的产酶能力进行初步判断。菌株GZFH7在不同碳源上的生长情况如图4所示,45 ℃培养相同时间后,分别以果胶、纤维素、木聚糖、淀粉、魔芋精粉为单一碳源的生长情况分别如图4B、C、D、F、H所示,菌株长势良好,菌丝均布满整个平板。以几丁质、壳聚糖为单一碳源的生长情况分别如图4E、G所示,菌株长势较差。另外菌株在不同碳源培养基上的菌落形态基本一致,但产生色素情况差异较大。因此,菌株GZFH7碳源谱较广,除了产生甘露聚糖酶外,还具有产果胶酶、几丁质酶、淀粉酶、木聚糖酶、纤维素酶等的可能性,是待开发利用的理想菌株,在食品工业酶制剂生产中有较大应用潜能。
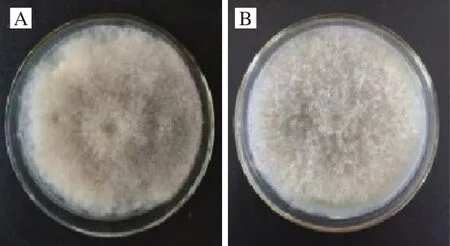

图4 菌株GZFH7在单一碳源平板上的生长情况Fig.4 Growth status of strain GZFH7 on single carbon source plates
2.6 菌株GZFH7的转录组学分析
2.6.1 Unigene的功能注释分析
对Unigene进行基因功能注释。在Nr数据库中被注释到的Unigene最多,共8 586 个,占总数的69.73%;其次为eggNOG数据库和Swiss-Prot数据库,分别为6 876、6 543 个,占总数的55.84%和53.13%;在数据库Pfam中被注释到的最少为1 588 个,仅占12.90%。778 个Unigene在所有的数据库中都被注释到,占总数的6.32%。
通过BLAST程序,将Unigene序列与Nr数据库比对,共有8 586 个Unigene被注释到,占总Unigene的69.73%。微小根毛霉GZFH7转录组测序组装的Unigene与横梗霉的相似性最多,为2 810 个,占32.75%;其次是伞状毛霉菌,比对上的Unigene为2 262 个,占26.36%。值得注意的是,有19.75%的Unigene属于其他序列,可能包含微小根毛霉与大多数物种不同的、自身特有的基因序列。
对微小根毛霉Unigene进行基因本体论(Gene Ontology,GO)功能分析。有4 987 个Unigene被注释到,共分为3 类41 个分支。统计每一类的基因数量发现,在细胞组成类的13 个分支中,涉及本转录组的Unigene最多,为11 124 个;生物过程类次之,为9 951 个;分子功能类最少,为5 452 个。其中细胞过程(2 645 个)、代谢过程(2 491 个)、催化活性(2 398 个)、细胞(2 367 个)、细胞组分(2 354 个)及连接(2 267 个)功能组涉及的Unigene较多,多细胞生物过程(8 个)、病毒(8 个)、病毒组分(8 个)、拟核(5 个)、通道调节活性(2 个)、蛋白标签(2 个)及翻译调节器活动(2 个)功能组涉及的Unigene较少。
经Unigene的京都基因与基因组百科全书(Kyoto Encyclopedia of Genes and Genomes,KEGG)数据库注释分析,微小根毛霉GZFH7共有4 367 个Unigene在KEGG数据库中被注释到,占总Unigene的35.46%,共涉及代谢、遗传信息处理、环境信息处理、生物系统以及细胞过程中5 个一级层级。其中被注释最多基因个数的为遗传信息处理过程中的翻译(494 个),其次是环境信息处理中的信号传导(469 个)、代谢过程中的碳水化合物代谢(464 个)。生物系统过程中被注释最多基因个数的为内分泌系统(232 个),细胞过程中被注释最多基因个数的为运输和分解代谢(313 个)。这些Unigene及其注释信息为今后对菌株GZFH7的代谢途径及相关功能基因的更进一步研究提供一定理论依据。
2.6.2 菌株CAZymes基因表达分析
通过对菌株GZFH7的转录组测序分析,共获得该菌的4.38×107条reads,数据量为6.56×109bp,GC含量为46.80%,Q20值为97.36%,Q30值为92.95%,Clean Data值为93.76%。大部分碱基序列Q值分布在33~37之间,代表碱基质量良好,可进行后续分析。
以魔芋胶、麸皮以及酒糟的混合物为碳源时,菌株GZFH7共有253 个CAZymes基因表达,表达结果如图5所示,其中包括31 个CEs基因,95 个GHs基因,104 个GTs基因,1 个PLs基因,16 个AAs基因以及6 个CBMs基因,与Hüttner等[27]对R.pusillusFCH 5.7的研究结果基本一致(12 个CEs基因、81 个GHs基因、77 个GTs基因、2 个PLs基因、11 个AAs基因、1 个CBMs基因)。其中GTs基因的表达最为丰富,约占CAZymes酶类表达总数的41.1%,约占总GTs家族的27.4%(29 种),主要包括纤维素合成酶、几丁质合成酶、透明质酸合成酶、葡萄糖基转移酶、甘露糖基转移酶、葡聚糖合成酶、乙酰氨基葡萄糖氨基转移酶、磷酸合成酶等。其次为GHs基因,约占CAZymes酶类表达总数的37.5%,约占总GHs家族的22.2%(34 种),主要种类有几丁质酶、半乳糖苷酶、葡萄糖苷酶、葡聚糖酶、甘露糖苷酶、甘露聚糖酶、淀粉酶、木聚糖酶、纤维素酶、海藻糖酶等。因此,菌株R.pusillusGZFH7酶系较丰富,是高效产碳水化合物活性酶的菌株。

图5 R.pusillus GZFH7碳水化合物活性酶基因表达分布Fig.5 Distribution of CAZymes gene expression in R.pusillus GZFH7
3 讨 论
微生物来源的甘露聚糖酶因其资源丰富、成本低、易培养、活性高等优势[17,28],是科学研究以及工业应用的长期热点。大曲所含微生物种类丰富,组成复杂,是极具利用价值的资源库[29-30]。菌株GZFH7为大曲中分离、筛选的嗜热真菌,通过对其菌落形态特征观察以及ITS序列分子鉴定,判定该菌为微小根毛霉(R.pusillus),命名为R.pusillusGZFH7。现阶段研究的较多的是利用微小根毛霉产凝乳酶[31],除凝乳酶之外,还有其他一些已产生并鉴定编码的酶,如聚半乳糖醛酸酶、糖淀粉酶、植酸酶和葡聚糖酶[27]。姚灿等[32]发现该嗜热真菌能产生淀粉酶和酸性淀粉酶,具有应用于白酒生产的开发潜力。但利用微小根毛霉产甘露聚糖酶的研究鲜有报道。
目前,实际投入生产应用的甘露聚糖酶大多为中低温酶且热稳定性较差,无法满足工业生产过程中部分高温环节对温度的要求。如来源于密黏褶菌ATCC 11539的甘露聚糖酶,在50 ℃处理30 min,酶活力剩余60%[33];来源于枯草芽孢杆菌MAFIC-S11的β-甘露聚糖酶Mann S,在60 ℃处理10 min,酶活力仅剩余10%[34]。菌株GZFH7所产的甘露聚糖酶最适反应温度70 ℃,为嗜热酶,并且该酶在70 ℃处理1 h仍能维持原酶活力不变,热稳定性优良。该嗜热甘露聚糖的最适pH值为5.0,与目前已获得的大多数真菌来源的甘露聚糖酶的最适pH值一致[35-37],且在pH 2.0~11.0的范围内均表现较理想的pH值耐受性。该酶满足工业生产需求,有效解决甘露聚糖酶的生产应用受限问题。魔芋胶溶液(1%)经酶解后黏度降低51 mPa·s,且酶解产物的mDP为0.96,而多数β-甘露聚糖酶水解魔芋胶、角豆胶等主要产生聚合度为2~5的甘露寡糖和一些带半乳糖残基侧链的半乳甘露寡糖[38]。此结果很可能是由于粗酶液中的甘露聚糖酶与其中的多种酶协同作用而导致的。
菌株GZFH7在不同单一碳源的平板上45 ℃培养时均能生长,其中以果胶、纤维素、木聚糖、淀粉、魔芋精粉为单一碳源的平板长势较好,以几丁质、壳聚糖为单一碳源的平板,长势较差,这可能是由于R.pusillusGZFH7中缺乏参与几丁质、壳聚糖降解的基因导致的。经转录组分析,共发现253 个CAZymes基因表达,其中包括以纤维素合成酶、甘露糖基转移酶、葡聚糖合成酶、乙酰氨基葡萄糖氨基转移酶等为主的GTs,以及以几丁质酶、半乳糖苷酶、淀粉酶等为主的GHs。综上所述,菌株R.pusillusGZFH7碳源谱较广,酶系丰富,是一株高效产碳水化合物活性酶的菌株,具有较高的研发利用价值。
4 结 论
本研究从大曲中分离嗜热真菌微小根毛霉R.pusillusGZFH7,在对该菌株诱导的甘露聚糖酶进行相关性质研究的同时,对菌株进行转录组学的分析。微小根毛霉GZFH7为嗜热真菌,其诱导的嗜热甘露聚糖酶有较好的热稳定性和pH耐受性。转录组学结果表明,菌株GZFH7有较好的产碳水化合物活性酶的能力。本研究不仅丰富嗜热甘露聚糖酶资源,同时也为微小根毛霉GZFH7在酶制剂的开发与应用提供参考。
