返青期休牧对退化高寒草地植被特征及优势种光合生理的影响
2021-09-17秦金萍马玉寿王彦龙李世雄
秦金萍,刘 颖,马玉寿,王彦龙,李世雄
(青海省高寒草地适应性管理重点实验室,青海省畜牧兽医科学院,青海大学,青海 西宁 810016)
青藏高原是我国重要的绿色生态屏障,在涵养水源和调节气候等方面发挥着极其重要的作用,同时也是畜牧业的主要基地[1-4]。但近年来,由于人为活动干扰和全球气候变暖等因素的影响,导致高寒草甸面积减小,物种多样性减少及优势牧草的光合速率降低等[5]。放牧作为草地的主要利用方式[6],是一个复合型的影响因子,放牧不仅包括家畜采食作用的影响,还包括践踏对草地土壤水分等的影响,这种由放牧所带来的影响都会直接或间接作用到牧草的形态特征及生理机制,使牧草的株高、盖度,牧草叶片的光合作用、蒸腾速率,以及呼吸作用等发生变化[7-8],进而对草地生产力产生影响。
返青期作为草地最敏感脆弱时期,此期间放牧会明显降低草地的生产力,进一步减少草地生物量[9]。而大量研究表明如在返青期休牧对草地植被株高、盖度和生物量等产生正效应[10-12]。因此在返青期即牧草的第一个忌牧期实行休牧,有利于草地的可持续利用[13-14]。目前在青藏高原已施行返青期休牧的地区均已取得良好成效,马玉寿等和李林栖等认为返青期休牧明显提高了草地的生产力[13-14],同时对牧草叶片光合和呼吸能力具有明显的促进作用,而在返青期施行长期休牧时,对草地牧草的盖度、株高和生物量及光合能力的促进作用尤为显著[15]。
光合作用是植物生长发育的基础,同时也是决定植物生产力的重要因素,可以直观的反映草地植物的生长状况[16-18]。对于天然草地,家畜的放牧践踏和采食是草原植被的重要胁迫因素,会影响植物的光合作用[16-18]。研究表明过度放牧显著降低牧草的光合速率、蒸腾速率和水分利用效率[16,19-21]。孙英等对祁连山披碱草-嵩草型草地4种代表植物的光合特性和叶绿素荧光特性的研究表明,放牧后黄花棘豆(Oxytropisochrocephala)这种毒草的光合能力提高,而其他3种优质牧草如垂穗披碱草(ElymusnutansGriseb.)等的光合能力下降[17]。植物的呼吸作用为植物体的生命活动提供能量,对植物的生长同样尤为重要,草地植物的光合作用和呼吸作用是相互伴随发生的生理过程[22],植物的光合特性和呼吸特性的变化可以反映植物受放牧干扰后的生长状况。因此,为确定返青期休牧6年后植被恢复情况及返青期休牧对牧草光合生理过程的影响,本研究对青海省祁连县默勒镇瓦日尕村退化高寒草地连续6年实施了返青期休牧,通过测定高寒草地优势牧草的数量特征、生物量、光合作用、叶绿素荧光特性及呼吸作用等指标,对实施返青期休牧6年后退化草地的植被状况进行评价,为青藏高原高寒草地的合理及可持续利用提供理论依据。
1 材料与方法
1.1 试验地概况
试验地位于青海省祁连县默勒镇瓦日尕村,地理位置为37°56′ N,100°13′ E,海拔3 650 m;属高原大陆性气候,光能资源丰富,太阳辐射强;年均气温1.4℃,1月均温—13℃,7月均温12.2℃,气温日差较大,冷季长,暖季短;雨热同季,年均降水量614 mm,年均蒸发量为1 164 mm;土壤主要以高寒草甸土为主,草地物种组成丰富,植被主要以莎草科和禾本科植物为主,伴生有大量阔叶型杂类草,主要物种为高山嵩草(Kobresiapygmaea),矮嵩草(Kobresiahumilis),垂穗披碱草和冷地早熟禾(Poacrymophila),是典型的高寒草甸植被类型[14,23]。
1.2 试验设计
选取12 hm2草地状况较均一、利用模式相同的天然高寒草甸草地(冬春草场)为试验区,其中6 hm2作为处理区,于2015年开始实施返青期休牧(每年的5月10日至7月10日),另6 hm2作为对照区,按当地传统放牧方式(放牧强度为中重度,即牧草利用率达到50%以上[24])进行放牧。试验区草地退化等级均为中度(即草地可食牧草比例为5%~15%)[25]。对照区和处理区各划分为 3个重复,每个重复2 hm2。每个重复小区选择具有代表性的9个点分别测定草地总盖度(Coverage,Co),地上生物量(Aboveground biomass,AB),地下生物量(Underground biomass,UB),土壤含水量(Soil water content,SWC),优势种牧草株高(Height,He),叶片净光合速率(Net photosynthetic rate,Pn),蒸腾速率(Transpiration rate,Tr),胞间CO2浓度(Intercellular CO2concentration,Ci),气孔导度(Stomatal conductance,Gs),呼吸速率(Respiratory rate,Re),叶面积(Leaf area,LA),叶温(Leaf temperature,LT),相对叶绿素含量(Relative chlorophyll content,RCC),叶片氮含量(Leaf nitrogen content,LNC),叶片含水量(Leaf water content,LWC),光化学量子效率(Photochemical quantum efficiency,Fv/Fm),有效光化学量子效率(Effective photochemical quantum efficiency,Fv′/Fm′),实际光化学量子效率(Actual photochemical quantum efficiency,φPSⅡ),光化学猝灭系数(Photochemical quenching coefficient,qP)和电子传递速率(Electron transfer rate,ETR)[26]。于2020年8月即高寒草地植物生产力最高时期进行指标的测定。根据本课题组前期试验结果,本试验选取的优势种为垂穗披碱草[14]。
1.3 观测指标及测定方法
草地植被数量特征测定采用样方法,样方面积为50 cm×50 cm,具体方法如下。
盖度(%):采用目测法测定样方内植被的总覆盖度。
株高(cm):采用钢卷尺在各样方内选择优势禾草垂穗披碱草5株测其自然株高,求其平均值为该样方优势禾草的平均株高。
地上生物量(g·m-2):齐地面刈割样方内所有植物,装信封袋并带回室内在75℃恒温下烘干至恒重,其干重为地上植物量。
地下生物量(g·m-3):在样方内用根钻(内径5 cm)取植物地下根系(0~15 cm)5钻,待清水冲洗干净后烘干至恒重,为样方内植被的地下植物量。
叶片含水量(%):选取10株具有代表性的优势种牧草垂穗披碱草,剪取其叶片并称鲜重,并带回室内在75℃恒温下烘干至恒重,得出干重并计算叶片含水量。
土壤含水量(%):采用土壤水分测定仪(ZY1200便携式)测定,在各样方内测定9次土壤含水量,求取平均值作为该处理区土壤含水量。
优势种光合特性、叶绿素荧光参数和呼吸速率:利用Li-Cor 6800(Li-Cor,Lincoln,NE)便携式光合仪测定优势种植物垂穗披碱草叶片的净光合速率、蒸腾速率、胞间CO2浓度、气孔导度、呼吸速率和叶绿素荧光参数[26],叶绿素荧光参数包括光化学量子效率、有效光化学量子效率、实际光化学量子效率、光化学猝灭系数和电子传递速率。数据测量前进行开机检查及预热0.5 h。自然光下诱导1.5 h~2 h后,进行光下相关参数的测量,暗适应0.5 h后进行暗反应相关参数的测量。
相对叶绿素含量、叶片氮含量及叶温:采用TYS-4 N叶绿素仪测定[27],在每个样方内选择3株生长良好的垂穗披碱草进行测定(取平均值)。SPAD测定值为叶绿素估测值,未进行估测值与实测值的拟合。
叶面积:采用手持式扫描仪(iScan 01)扫描优势种牧草叶片,并用Digimizer(3.1.2.0)计算叶面积。
1.4 数据处理
试验数据用Excel 2010进行初步整理统计,运用SPSS 20.0软件对其进行独立样本T检验,显著性差异水平为0.05。各指标间的相关性用pearson相关性进行分析,相关性大小用Student’s t进行检验。数据统计分析与绘图在RStudio中完成。
2 结果与分析
2.1 返青期休牧对高寒草地植被数量特征的影响
在祁连山区高寒草地植物生长盛期对草地植被盖度、株高和生物量进行测定(表1),结果表明:在返青期未进行休牧时,高寒草甸植被总盖度仅为71%,而返青期实施6年休牧后,其草甸植被盖度可达100%,与未休牧草地的植被盖度差异显著(P<0.05);祁连山区高寒草甸优势禾草垂穗披碱草的株高在返青期实施休牧6年处理后为61.59 cm,与未休牧处理区株高差异显著(P<0.05);返青期未休牧时,祁连山区高寒草甸的地上生物量仅为5.99 g·m-2,返青期实施休牧6年后增加了10倍,草地的生产力显著提高;同时,地下生物量也因实施返青期休牧显著增加(P<0.05)。

表1 返青期休牧对高寒草地植被数量特征的影响
2.2 返青期休牧对垂穗披碱草光合特性和呼吸速率的影响
净光合速率反映了植物光合作用的强弱,祁连山区高寒草甸实施返青期休牧显著提高了其优势禾草垂穗披碱草叶片的净光合速率(表2);垂穗披碱草叶片的蒸腾速率在返青期休牧6年后达到5.00 mmol·m-2·s-1,与未休牧区差异显著(P<0.05);气孔导度和胞间CO2浓度在返青期未休牧时分别为0.07 mmol·m-2·s-1和274.88 μmol·m-2·s-1,实施休牧6年后,祁连山区高寒草甸垂穗披碱草叶片的气孔导度和胞间CO2浓度分别达到为0.23 mmol·m-2·s-1和307.39 μmol·m-2·s-1,与未休牧时差异显著(P<0.05)。随着净光合速率的显著增加,祁连山区退化高寒草地优势禾草垂穗披碱草叶片呼吸速率也因实施返青期休牧显著增加(P<0.05)。

表2 返青期休牧对垂穗披碱草光合特性的影响
2.3 返青期休牧对垂穗披碱草叶绿素荧光参数的影响
植物叶绿素荧光参数是研究植物光合生理生态和抗逆能力的主要参数。返青期休牧使高寒草甸优势禾草垂穗披碱草叶片的5个叶绿素荧光参数均发生了变化(表3)。在未休牧区,垂穗披碱草的Fv/Fm,Fv′/Fm′,φPSⅡ,qP和ETR分别为0.74,0.34,0.16,0.43和138.62;返青期休牧6年后垂穗披碱草的Fv/Fm,Fv′/Fm′,φPSⅡ,qP和ETR分别为0.77,0.42,0.19,0.45和164.84。其中优势禾草垂穗披碱草的Fv/Fm,Fv′/Fm′,φPSⅡ和ETR在休牧6年后显著高于未休牧区(P<0.05),而垂穗披碱草的qP在休牧6年后与未实施休牧差异不显著。

表3 返青期休牧对垂穗披碱草叶绿素荧光参数的影响
2.4 返青期休牧对影响高寒草甸优势禾草垂穗披碱草叶片光合作用和呼吸作用关键因素的影响
高寒草甸土壤含水量和优势禾草垂穗披碱草叶片含水量的测定结果显示,返青期未休牧的高寒草甸土壤含水量和垂穗披碱草叶片含水量较低(表4),分别为18.02%和58.50%,经过6年返青期休牧后,土壤含水量和植物叶片含水量均有显著增加,可达到29.30%和72.70%(P<0.05)。如表4所示,返青期未休牧区优势禾草垂穗披碱草叶面积、相对叶绿素含量和叶片氮含量分别为3.09 cm2·叶-1,9.51 SPAD和5.90 mg·g-1,实施6年返青期休牧后,分别增加了7.87 cm2·叶-1,18.74 SPAD和5.67 mg·g-1,均显著高于未休牧区(P<0.05),而叶面温度与未休牧区差异不显著。
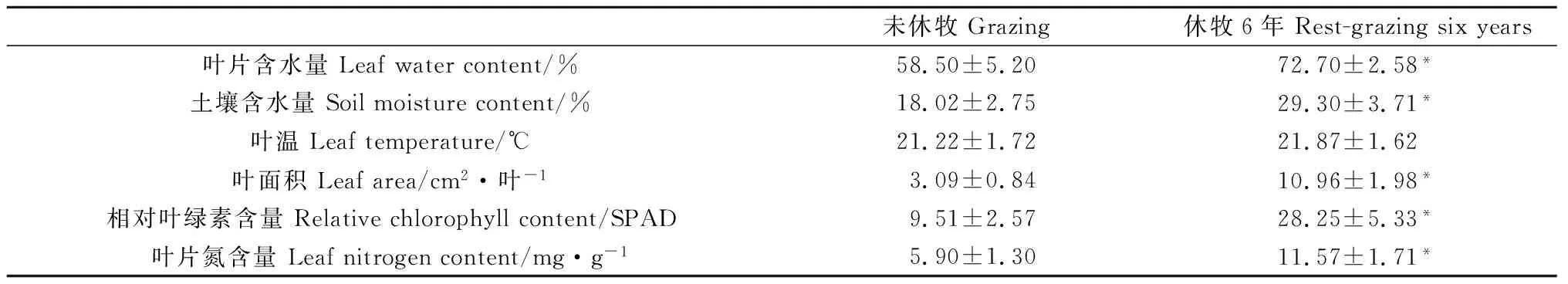
表4 返青期休牧对影响高寒草甸优势禾草垂穗披碱草叶片光合作用和呼吸作用关键因素的影响
2.5 退化高寒草地植被数量特征与光合特性、呼吸速率及关键影响因素之间的相关性
本研究中,退化高寒草地植被盖度与所有指标呈显著相关(P<0.05),相关系数较高的5个生理指标为净光合速率、呼吸速率、叶面积、气孔导度和叶片相对含水量,相关系数分别为0.912,0.892,0.877,0.862和0.842。垂穗披碱草株高与除光化学猝灭系数和叶温外的指标均呈极显著相关(P<0.01),相关系数较高的5个生理指标为净光合速率、呼吸速率、叶面积、气孔导度和蒸腾速率,相关系数分别为0.950,0.951,0.928,0.928和0.858。高寒草地植被地上生物量与除光化学猝灭系数和叶温外的指标均呈极显著相关(P<0.01),相关系数较高的5个生理指标为净光合速率、呼吸速率、叶面积、气孔导度和叶片相对叶绿素含量,相关系数分别为0.953,0.950,0.938,0.936和0.905。高寒草地植被地下生物量与除光化学猝灭系数和叶温外的指标均呈极显著相关(P<0.01),相关系数较高的5个生理指标为净光合速率、气孔导度、呼吸速率、叶片相对叶绿素含量和叶面积,相关系数分别为0.922,0.900,0.890,0.888和0.873。另外,光化学猝灭系数和叶温与所有指标的相关性均较低。

图1 退化高寒草地植被数量特征与光合特性、呼吸速率及关键影响因素之间的相关性分析
3 讨论
3.1 返青期休牧对退化高寒草甸植物数量特征的影响
高寒草地的主要利用方式为放牧[17],而放牧干扰会影响植物的盖度和株高等。植被盖度和株高作为评价草地植物生长状况的重要指标,放牧通常会减少植物的盖度和株高,尤其在高寒草地牧草生长敏感时期返青期进行放牧会明显降低草地植被的覆盖度[13-14]。本研究中,未实行返青期休牧区,退化高寒草地植被覆盖度和其优势禾草垂穗披碱草的株高较低,而返青期休牧6年后草地植被覆盖度和优势禾草垂穗披碱草株高显著提高了38.44%和965.77%。生物量是反映草地生产力的重要指标,在植物受到干扰时,草地植物生物量会减少,特别是在高寒草地牧草返青期放牧对植物生物量的影响尤为明显[28]。另外,实施返青期休牧不仅可以消除高寒草地植物生物量减少的现象,而且可以促进植物的生长发育,提高草地生产力[13,29]。本研究中,退化高寒草地优势禾草垂穗披碱草的地上生物量和地下生物量在实施6年返青期休牧后较未休牧处理分别提高了985.00%和209.56%,这与马玉寿等的研究结果“返青期休牧可明显增加高寒草地植被覆盖度、促进牧草生长及显著提高草地生产力”和李青丰等的研究结果“春季休牧提高了草地的生产力,且增加休牧年限效果越明显”的结果一致[13,29]。此结果可能是由于高寒草地牧草返青期时植物比较脆弱,在此期间放牧采食及践踏干扰了植物正常生长,减少了垂穗披碱草的分蘖、萌芽等数量,迫使植物体内的营养物质重新分配,而牧草根部储存的营养物质无法满足牧草正常生长发育,甚至无法形成正常的种子,导致返青期放牧后高寒草地的盖度、牧草株高和草地生产力降低[15],而实施返青期休牧后可以避免这种现象。另外,正向促进退化高寒草地植被数量特征的光合生理及影响因素的指标主要为净光合速率、呼吸速率、气孔导度、蒸腾速率、叶面积、叶片相对叶绿素含量和叶片相对含水量。因此,本研究认为返青期休牧可增强牧草的光合作用及呼吸作用过程,促进牧草的生长发育,进而促进退化高寒草地的恢复。
3.2 返青期休牧对退化高寒草甸优势禾草垂穗披碱草叶片光合作用的影响
高寒草地生态系统的主要功能是植物通过光合作用固定太阳能用于植物生产,供家畜采食,同时光合作用可以反映高寒草地植物的生长状况[17]。作为反映植物光合作用强弱的重要指标之一,净光合速率通常是指单位时间单位叶面积所吸收的二氧化碳或释放氧气的量,当植物生长过程中受到干扰时,其净光合速率会发生变化,植物叶片的蒸腾速率、气孔导度和胞间CO2浓度作为重要的光合参数,随植物受到干扰也会发生相应的变化[30],这些变化可进一步反映当下植物光合作用的强弱。在本研究中,祁连山区高寒草地优势禾草垂穗披碱草叶片净光合速率与蒸腾速率、气孔导度和胞间CO2浓度均呈极显著正相关(P<0.01),垂穗披碱草叶片的净光合速率在实施6年返青期休牧后提高了139.86%,这与孙英等和胡新振等的研究结果“围封可提高高寒草地优质牧草的光合能力”结果一致[17,31]。此结果可能是由于返青期作为高寒草地植物生长脆弱时期,此时放牧家畜对幼嫩植物的采食及践踏,对植物后期生长具有负面影响[13,17],也可能是由于土壤含水量和垂穗披碱草叶片含水量的降低使叶片气孔关闭[32],导致叶片的蒸腾速率下降,气孔导度也为适应环境而降低,造成高寒草地优势禾草垂穗披碱草叶片光合速率等较低,而实施6年返青期休牧后,植物得到充分生长,发育完全,有利于优势禾草垂穗披碱草叶片的光合速率升高,进一步促进植物生长。
植物叶片叶绿素荧光参数是研究植物光合生理状态及抗逆能力的重要指标[33-34]。叶绿素荧光参数对外界环境条件变化十分敏感,可用于评价外界干扰对植物的影响等[26,34-37],其中Fv/Fm是暗适应条件下PSⅡ的最大光化学效率,反应PSⅡ中心最大光能转化效率,能够快速、敏感地反映植物受到强光胁迫的程度。研究表明,未受到胁迫时多数植物的Fv/Fm通常处于0.75~0.85 之间[38-40],而在逆境或受损伤时植物的Fv/Fm会下降[35,41]。Fv′/Fm′是植物的有效光化学量子效率即实际光化学量子效率,其变化情况与Fv/Fm一致。实际光化学量子效率(φPSⅡ)反映植物实际的光合能力[42]。光化学猝灭系数(qP)是由光合作用所引起的荧光淬灭,指植物激发能被反应中心捕获而转化为光化学能的途径[43],反映了植物光合活性的高低[17,44]。电子传递速率(ETR)表示实际光强下光合电子传递速率,在胁迫下植物叶片的ETR会下降[43]。本研究中,未休牧区的植被因连续放牧一直处于胁迫下,而经过6年返青期休牧的休牧区植被胁迫得以缓解,其优势种垂穗披碱草叶片的叶绿素荧光参数均有显著提高,Fv/Fm,Fv′/Fm′,φPSⅡ,qP和ETR分别提高了4.92%,22.66%,19.07%,4.85%和18.91%。本结果与孙英等的研究结果“放牧时披碱草的Fv/Fm,qP和ETR值低于围封处理”一致[17]。另外,叶绿素荧光参数Fv/Fm,Fv′/Fm′,φPSⅡ,qP和ETR均与垂穗披碱草叶片净光合速率显著正相关(P<0.05),其原因可能是放牧家畜的采食、践踏使垂穗披碱草的光合器官明显受损,光化学活性发生改变,光合电子传递速率降低[17],使垂穗披碱草的实际光合能力降低,有较多的激发能以热的形式散发。
3.3 返青期休牧对退化高寒草甸优势禾草垂穗披碱草叶片呼吸速率的影响
植物呼吸作用为植物生理代谢的基本过程,可为植物的各种生命活动提供所需的能量[45-46],并受植物体内外多种因素的影响。本试验中,返青期休牧6年后垂穗披碱草叶片的呼吸速率较未休牧时显著提高269.37%,与齐杨等的研究结果“放牧降低植物呼吸速率”一致[22]。该结果可能是由于未休牧的高寒草地土壤含水量较低,垂穗披碱草植株矮小,其叶片含水量和净光合速率较弱,垂穗披碱草光合产物降低,致使叶片呼吸作用可利用底物减少进而导致呼吸作用较弱[47]。另外,本研究指出与呼吸速率相关系数最高的就是净光合速率,进一步佐证了上述论点。
3.4 返青期休牧对影响退化高寒草甸优势禾草垂穗披碱草叶片光合作用和呼吸作用关键因素的影响
水是动植物生命活动不可或缺的[48],土壤含水量高低会影响植物的光合生理过程[49-50],进而影响植物的生长。高寒草地土壤含水量在实施返青期休牧6年后较未进行休牧区,其土壤含水量显著提高62.62%,较高的土壤含水量更有利于植物的生长,这与张子荷等的研究结果“放牧地土壤含水量低于围封处理”一致[18]。其原因可能是返青期放牧家畜对草地的践踏,导致土壤的空隙减小,容重增加,土壤水的渗透阻力增加,进而使高寒草地土壤的保水能力下降,土壤含水量降低[51]。植物叶片含水量反映植物生命活动的强弱,一般生命活动越旺盛,含水量越高,生长活跃和代谢旺盛的植物叶片的含水量通常在60%~90%[17-18]。本研究中祁连山区退化高寒草地优势禾草垂穗披碱草的叶片含水量在返青期休牧6年后较返青期未休牧区增加了24.26%,这与晏欣等的研究结果“放牧后狼针草(Stipabaicalensis)的叶片含水量低于围封处理”一致。其原因可能是与放牧使土壤含水量的降低有关。
叶片作为植物进行光合作用的场所,叶面积大小对于植物光合作用具有重要影响,一般植物叶面积越大,叶片光合作用转化的能量越多。本研究表明,优势禾草垂穗披碱草叶片净光合速率与其叶片面积极显著正相关(P<0.01),其面积在实施6年返青期休牧后较未休牧区增加了254.13%,这与李江文关于“放牧干扰下无芒隐子草(Cleistogenessongorica)单片叶面积显著减小”的结果一致[52],认为造成该结果的主要原因是放牧家畜的采食。本研究中,未休牧区优势禾草垂穗披碱草的叶温在21.5℃左右,与返青期休牧区差异不显著,说明返青期休牧对叶温的影响不显著,这可能是由于植物为更好适应外界环境,垂穗披碱草进行内部调控其叶面温度所致。
叶绿素作为光合色素,对绿色植物吸收和利用光能具有重要作用,叶绿素含量的多少与植物光合作用强弱密切相关[34-35]。当植物生长遭到干扰胁迫时,其叶绿素含量较低,使植物的光合原初反应遭到抑制[41]。本研究发现返青期休牧6年后,高寒草甸优势禾草垂穗披碱草的相对叶绿素含量较返青期未休牧区增加197.02%,此结果与秦金萍等的研究结果“返青期休牧增加了高寒草地植物相对叶绿素含量”和孙英等的研究结果“放牧降低了垂穗披碱草叶片叶绿素含量”是一致的[15,17]。这可能是由于返青期放牧干扰后,植物叶片含水量及土壤含水量等的降低,导致垂穗披碱草叶片相对叶绿素含量发生一定的降解[53]。本研究中叶片相对叶绿素含量与叶片氮含量、叶片含水量和呼吸速率极显著正相关(P<0.01),表明返青期休牧是通过改善原料与能量供应促进叶绿素的合成。并且,叶片相对叶绿素含量与光合特性参数也极显著正相关(P<0.01),表明返青期休牧不仅有利于高寒草地优势禾草垂穗披碱草叶片叶绿素的合成,而且可通过增加的叶绿素捕捉更多的光能促进光合作用过程。
氮素作为影响植物生长的主要营养元素[54],叶片氮含量对光合同化物的累积具有一定的影响[19]。在本试验中,实施返青期休牧6年后,高寒草甸优势禾草垂穗披碱草的叶片氮含量较未休牧区增加96.24%,且与净光合速率、叶片含水量和土壤含水量极显著正相关(P<0.01)。原因可能是由于在返青期放牧使得高寒草地土壤含水量和垂穗披碱草叶片含水量降低,而此干旱胁迫会对植物叶片氮素累积产生负面影响[55],降低的叶片氮含量进一步降低了净光合速率。结合本研究结果,表明返青期休牧在维持并增加垂穗披碱草叶片含氮量的同时,也提升了光合同化物的累积,进一步证明实施返青期休牧可促进垂穗披碱草的生长及高寒草地生产力的提升。
4 结论
返青期休牧6年后高寒草地植被的覆盖度、生物量和其优势禾草垂穗披碱草的株高均有显著提高,表明实施返青期休牧提高了高寒草地的生产力;返青期休牧6年后高寒草地土壤含水量、优势禾草垂穗披碱草叶片含水量、叶面积、叶绿素含量、叶片含氮量、净光合速率(Pn)、呼吸速率(Re)和叶绿素荧光参数均有显著提高,表明实施返青期休牧可通过改善植物的生长环境及生理代谢过程所需底物的积累,促进植物的光合作用和呼吸作用,进而提高草地的生产力,促进草地的可持续利用。
