中条山油松人工林下物种对群落物种丰富度分布格局的贡献
2021-09-16裴顺祥杜满义辛学兵
裴顺祥,法 蕾,杜满义,郭 嘉,辛学兵
(中国林业科学研究院华北林业实验中心,北京九龙山暖温带森林国家长期科研基地,北京 102300)
物种丰富度的分布格局是生物多样性保护的关键问题之一[1]。物种丰富度格局的形成不仅依赖于群落的构建过程,同样也依赖于群落中的物种组成[2-3]。群落中根据物种多度大小可以将物种划分为常见种、稀有种和介于常见种与稀有种之间的物种,在大多数群落中,稀有种比常见种的数量更多,且由于稀有和常见物种的频率分布不均匀而引发群落物种分布格局[4],因此研究物种对群落物种丰富度分布格局的贡献是探索物种分布格局的重要手段之一[5]。
关于物种对群落物种丰富度分布格局的贡献已开展了大量的研究,并取得了一些关键性结论,但仍存在不足,主要表现在以下2个方面。一是稀有种与常见种对群落物种丰富度分布格局贡献度的大小存在争议。群落的中性过程和个体水平上的随机作用导致群落中存在大量的稀有种[6],进而使群落产生“稀有种优势”[7-8],因此稀有种对整个群落的丰富度格局产生较大的影响[9-10],但也有学者认为常见种的贡献要远大于稀有种[11-13],即常见种“维持”了群落物种分布格局[14],而稀有种对整个群落功能的重要性可能被夸大[15],因为在实际群落中,常见种的生殖成功率要明显高于稀有种[16],且中性理论认为群落中所有物种竞争对称,即不存在“稀有种优势”。研究尺度也影响稀有种和常见种的贡献,在大尺度范围内,稀有种的贡献较大,而常见种在小尺度上影响物种丰富度,在大尺度上影响物种均匀度[17]。稀有种由于个体少,分布范围受限,与常见种相比更易受到环境变化和竞争排斥的干扰[18-19],其通常被认为是特有的、局地的和受威胁的,这使得稀有种比常见种面临更高的灭绝风险[20],但并不能根据稀有种的稀有性(频度、丰富度和生物量低、高灭绝风险)的特征,判定其对生态系统功能、过程中的贡献低于常见种[21]。此外,Mazaris等[22]认为常见种对物种分布格局的贡献大是取样偏差造成的,并非常见种与稀有种二者的生物学差异所致。二是以某一类物种组成的子群落对整体群落物种分布格局的贡献研究较多,单一物种的贡献研究较少,而单一物种的贡献可能对于群落物种多样性的保护更为重要。因此深入解析不同生态系统物种对群落物种丰富度分布格局的贡献,对于区域物种生物多样性保护和提升均具有重要的意义。
中条山地处秦岭与太行山之间,其植物区系是热带亚热带区系向温带区系的过渡和交汇地[23],而油松(Pinustabuliformis)人工林是中条山森林生态系统的重要组成部分,趋于针叶化的、成片高密度人工林导致土壤酸化,林下灌、草难以发育形成乔-灌-草垂直空间结构的森林群落,进而导致人工林下物种多样性低下、群落稳定性和生产力差、病虫害频发、抵御气候灾害能力弱[24-25],而物种多样性的恢复和保护是生态系统恢复和重建的基础。因此,开展油松人工林下物种多样性格局研究具有非常重要的现实和理论意义。本研究以中条山油松人工林群落为研究对象,研究林下不同大小的子群落对群落物种丰富度分布格局的贡献,并确定影响该区域群落物种丰富度分布格局的关键种,为区域物种多样性保护提供理论依据。
1 材料和方法
1.1 数据来源及质量
在对中条山油松人工林全面踏查及尽可能涵盖该地区所有油松人工林类型的基础上,设置12个样地,每个样地设置3个20 m × 20 m的用于乔木调查的样方(其中有3个样方受地形限制为15 m × 15 m),共计36个乔木样方,在每个乔木样方内设置16个5 m × 5 m和1 m × 1 m的小样方调查灌木、草本植物。分别调查了555个灌木和草本样方,记录每个样方内物种种类、数量、盖度、高度等指标,同时记录样地的经纬度、海拔、坡向、坡位、林冠郁闭度等环境特征(表1)。为了确定抽样调查是否可以充分反映群落物种多样性,本研究采用物种累积曲线来判断抽样量是否充分(图1),由图1可知,曲线趋于平缓,表示中条山油松人工林下的物种并不会随样本量的增加而显著增多,表明抽样充分[26]。
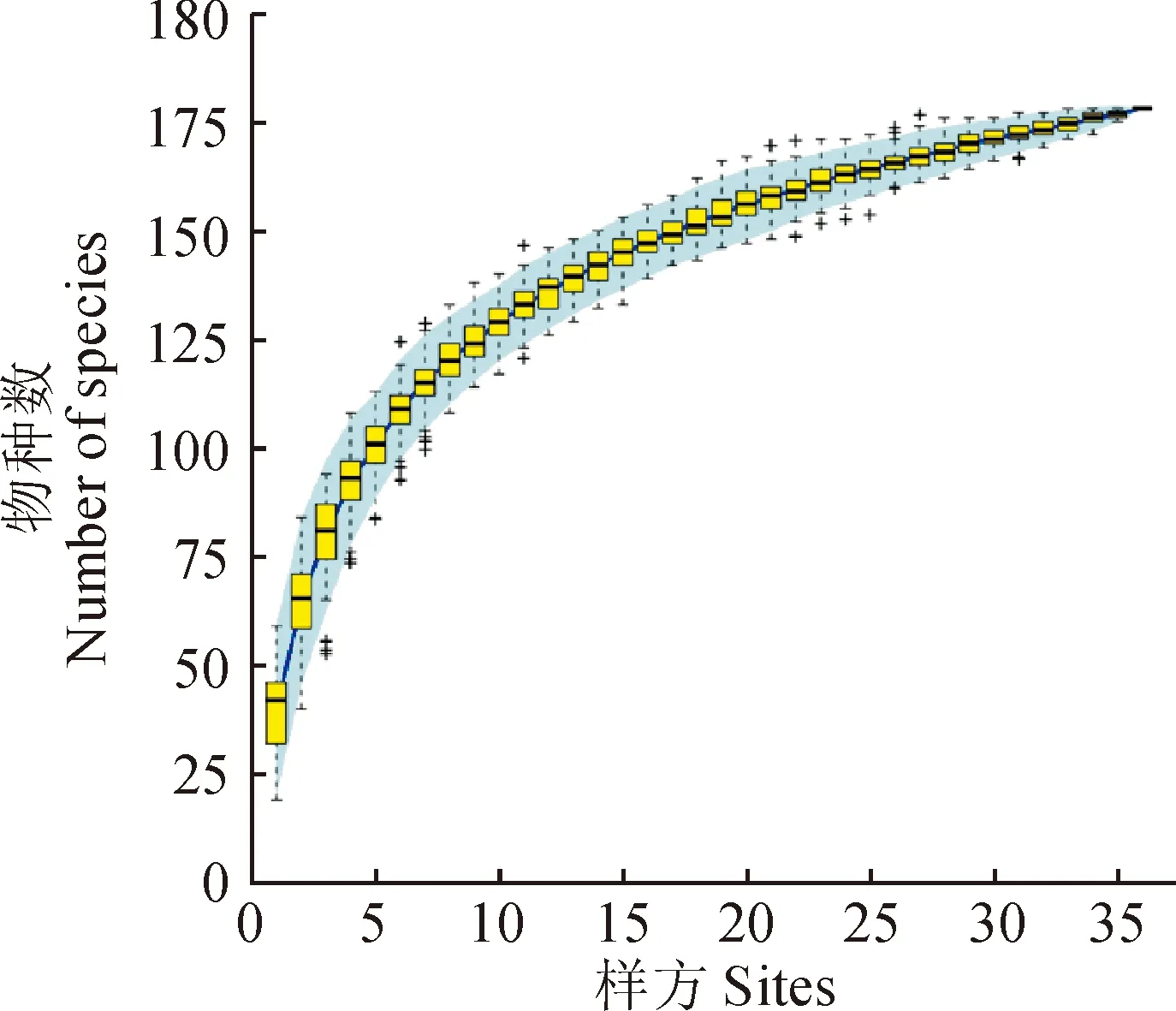
图1 灌木、草本、藤本物种累积曲线Fig.1 Species accumulation curves of shrubs, herbs and lianas
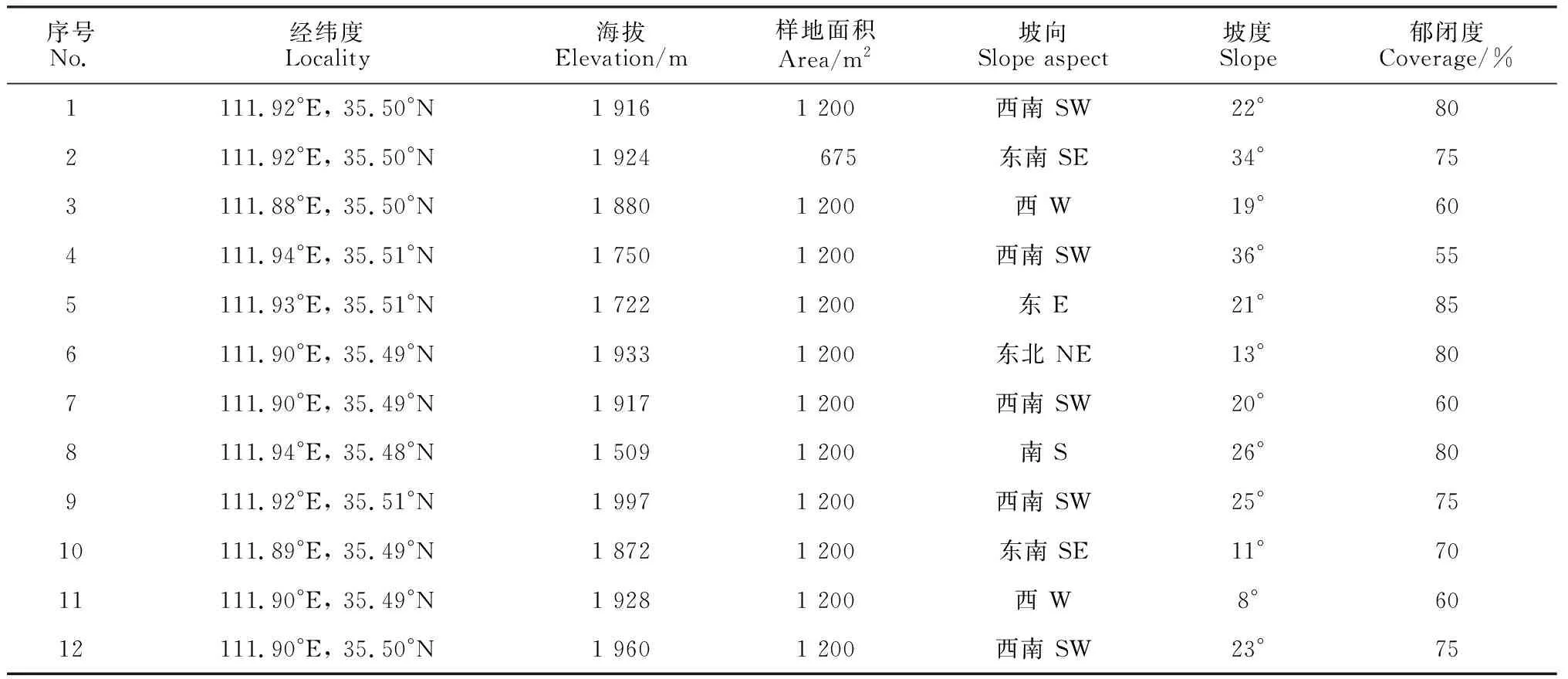
表1 油松人工林群落样地概况
1.2 数据分析方法
本研究选取物种优势度指数(Y)计算群落优势种,其计算公式如下:
式中,N表示群落物种个体总数,ni表示第i种的个体总数,fi表示第i种的频度。当Y>0.02时,该物种为群落优势种;选取物种丰富度(R)表示多样性;采用稀释曲线(rarefaction curve)法分析各样方物种多样性的差异[27];将频度小于等于2的物种为稀有种,频度大于9的物种为常见种[17]。
根据物种在群落中的频度定义物种稀有度和常见度,按频度由低到高或由高到低的序列对物种依次排序,然后根据序列逐一添加(或去除)物种,通过比较添加(或去除)前后引起的群落物种丰富度变化的大小来衡量稀有/常见种对群落物种丰富度格局的贡献[2-3]。
为了进一步分析最稀有/常见物种组成的子群落对整个群落物种丰富度格局的贡献,将占群落总体物种25%的最稀有/常见的物种组成子群落,分析子群落、整个群落、除25%最稀有/常见物种子群落间的相关关系[3]。
将物种依据频度由低到高排序,以稀有—常见为横坐标,常见—稀有为纵坐标,横坐标为“开始”种(starting species),纵坐标为“结束”种(finishing species)[3],178个物种共构建15 931个子群落,然后每个子群落与整个群落做相关分析,然后计算添加(或去除)物种后,子群落与整个群落相关系数变化量,依据变化量大小排序,得到影响群落丰富度格局最大的5个物种。然后将所有物种按频度由高到低排序,以贡献最大的种为例,基于Jaccard指数,分析贡献度最大的种与比其频度更高的物种间的关联度[28]。
物种数据整理采用Excel 2016(Microsoft,USA)完成,除Jaccard指数及种间关联分析及作图分别采用R 4.0.3软件中“spaa”程序包中的“sp.pair”和“plotlowertri”命令完成外,其他分析及作图分别采用R 4.0.3软件中“vegan”程序包中的“specaccum”、“ggplot”、“rarecurve”、“cor”命令完成。
2 结果与分析
2.1 中条山油松人工林下物种多样性
样地物种调查数据显示,中条山油松人工林下共有178种植物,其中灌木47种、草本112种,藤本19种,分属56科124属,优势种为披针薹草(Carexlancifolia)、茜草(Rubiacordifolia)、赤瓟(Thladianthadubia)、金线草(Rubiamembranacea)、南蛇藤(Celastrusorbiculatus)。频度-物种数分布图显示(图2),该地区物种频度分布格局呈明显右偏。具体表现为,频度小于等于2的稀有种54种,频度大于9的常见种59种,分别占全部物种的30.34%和33.15%。频度排名前10的物种分别为披针薹草、粗齿铁线莲(Clematisargentilucida)、金线草、披碱草(Elymusdahuricus)、接骨木(Sambucuswilliamsii)、鞘柄菝葜(Smilaxstans)、茜草、土庄绣线菊(Spiraeapubescens)、南蛇藤、红椋子(Swidahemsleyi),草本多于灌木。
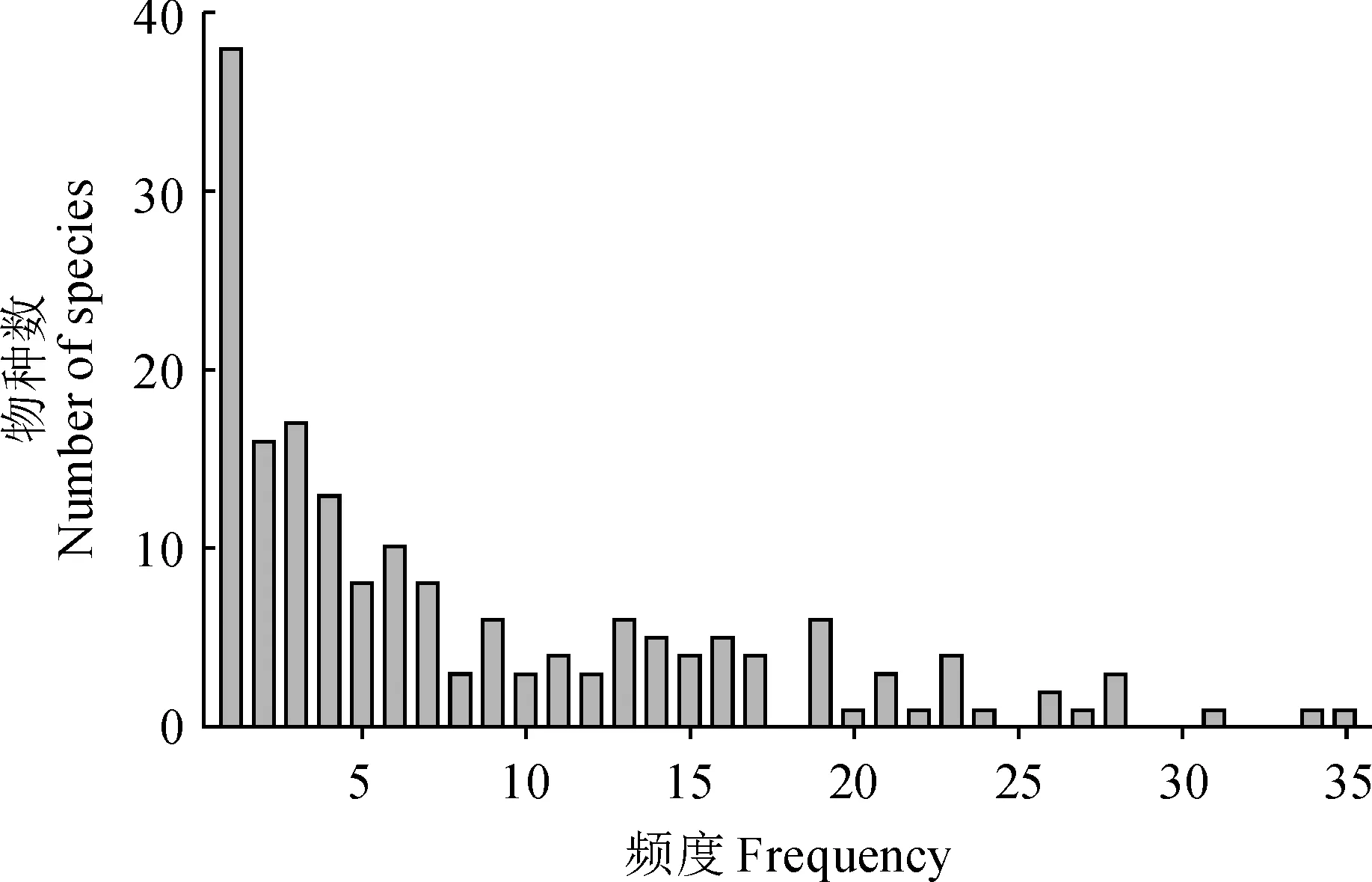
图2 中条山油松人工林下物种频度分布Fig.2 Species-frequency diagram for under P. tabuliformis plantations in Zhongtiao Mountain
基于中条山36个样方林下物种多度数据,绘制稀释曲线图(图3)。由图3可知,含有最多物种的是样方22(55个物种),而含有最少物种的是样方7(19个物种)。为了减小取样规模对物种数量的影响,将样本取样规模标准化,即基于220个个体取样规模的基础上比较不同样方的物种丰富度。标准化后发现5号样方的物种数最多(45.0个物种),而物种最少的样方仍为7号样方(18.9个物种),接近样方真实物种数据(55,19)。此外,随着取样个体数量的增加,5号样方的物种数增加最快,而21号样方增加最慢。
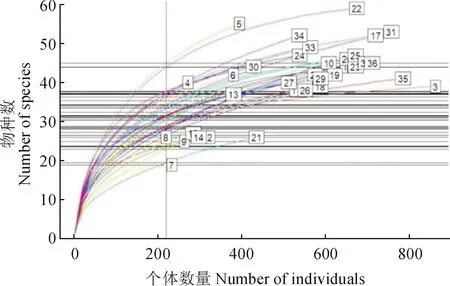
方框内数字(1~36)代表样方编号图3 36个样方林下物种多度稀释曲线图The number in the box (1-36) represent the plots numberFig.3 Refraction curves of 36 sample plots
2.2 稀有种与常见种对群落物种丰富度的贡献
稀有种与常见种对群落物种多样性的贡献分析结果(图4)显示,稀有种与常见种加入后群落的物种丰富度逐渐增大,其中,常见种加入后,群落丰富度指数呈对数增加,而稀有种加入后,群落丰富度指数呈指数增加,说明常见种对群落丰富度分布格局的贡献大于稀有种。
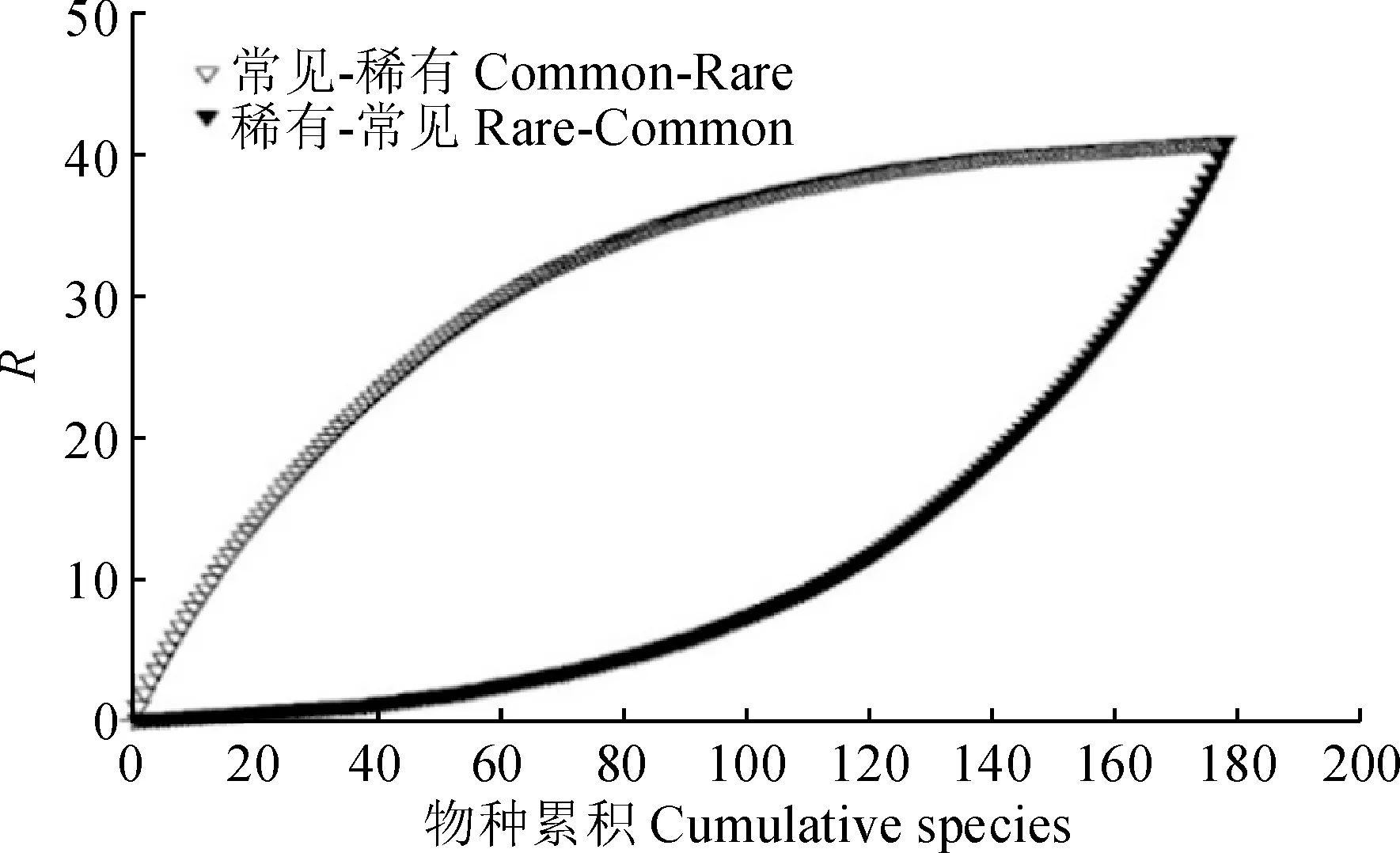
图4 稀有种与常见种对物种丰富度的贡献Fig.4 Contribution of rare and common species to species richness
占群落总体物种25%的最稀有/常见的物种(最稀有共54种,最常见共49种)、整个群落(178种)、除25%最稀有/常见物种(分别剩124种、129种)子群落间的相关关系分析结果如表2。由表2可知,最常见的物种解释了整个群落物种丰富度格局的88.4%(P<0.01),而最稀有物种仅解释了整个物种群落丰富度格局的24.5%(P>0.05)。去除最稀有后,最常见物种可以解释剩余物种的90.8%(P<0.01),而去除最常见物种后,最稀有物种仅能解释剩余物种的48.6%(P<0.01)。进一步说明常见种对物种群落丰富度分布格局的贡献大于稀有种。
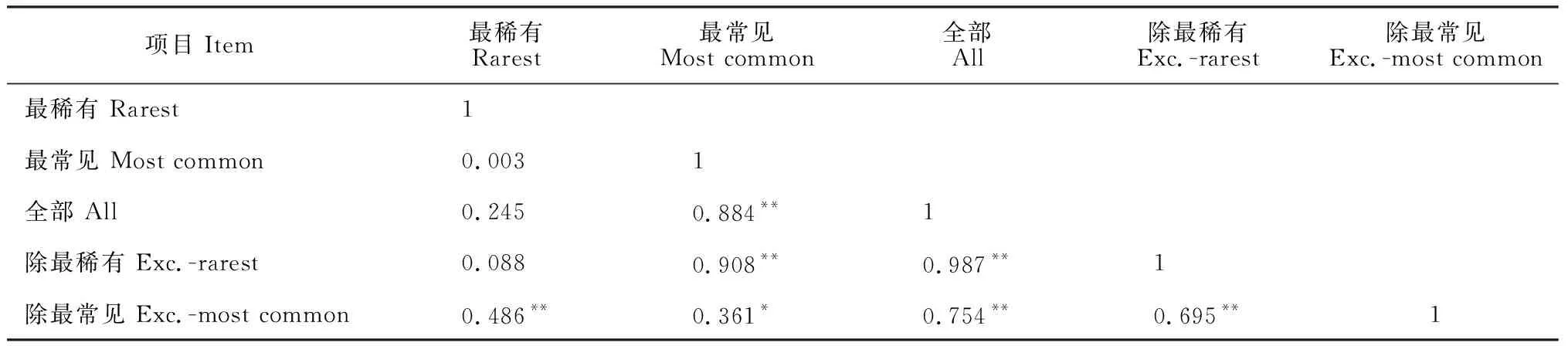
表2 物种丰富度格局间的相关性
2.3 子群落及单一植物种对群落物种丰富度的贡献
根据子群落与整个群落物种丰富度格局相关分析结果(图5)可知,当子群落中的“结束”种相对常见而“开始”种相对稀有时,相关性较高,因为整个群落的大部分物种被子群落选取。与直角三角形斜边平行,子群落的物种丰富度是一个常数,可以看到不同比例的常见物种对稀有物种的影响,即当子群落中常见种越多时,子群落与整个群落的丰富度分布格局相关性越高。
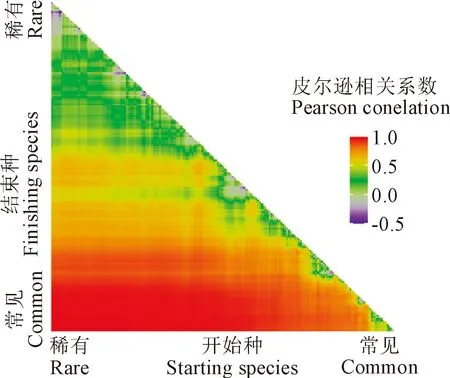
图5 子群落与整个群落物种丰富度格局的相关关系Fig.5 Correlation between sub-assemblages with the full assemblage species richness pattern
根据子群落与整个物种丰富度格局的相关关系结果,计算添加(或去除)某一物种后子群落与整个群落的相关系数变化量(图6)。由图6可知,相关系数变化较大的值均分布于直角三角形斜边上,特别是直角三角形斜边的右下方,其值变化值最大,而直角三角形斜边上的子群落一般由单一物种组成的群落,即一个种组成的群落添加一个物种后,群落的相关系数变化最大,且其呈垂直于横坐标轴的带状分布,即只要有该种存在的子群落,添加(或去除)任意物种后其相关系数变化均最大。按由大到小提取相关系数变化排名前5的物种,即影响该群落物种丰富度分布格局最大的物种,分别为连翘(Forsythiasuspensa)、太平花(Philadelphuspekinensis)、鞘柄菝葜、多歧沙参(Adenophorawawreana)和金花忍冬(Lonicerachrysantha),其中灌木4种,草本1种。将群落物种频度由高到低排序,以上物种频度分别为16、23、28、2和15,其中常见种4种,稀有种1种。由此结果可以得出,并非越常见的物种对群落丰富度格局影响越大。
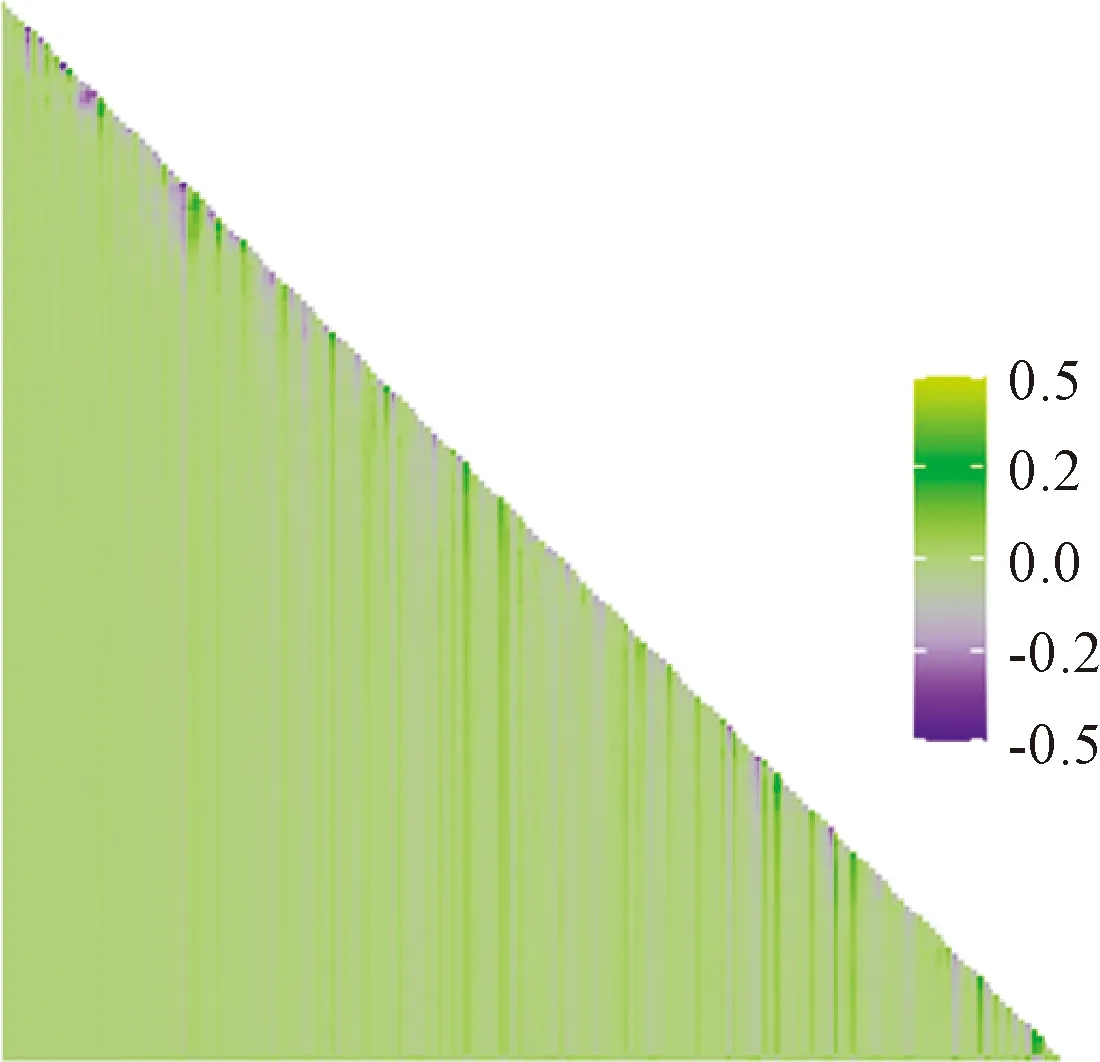
图6 添加(或去除)某一物种后子群落与整个群落的相关系数变化量Fig.6 The change of correlation coefficient between the sub- assemblage and the full assemblage after the addition (or removal) of a species
以连翘为例,基于Jaccard指数,分析连翘与比其频度更高的31个物种间的关联度(图7)。由图7可知,相比较于其他物种,连翘与其他物种之间的关联度最低,即连翘与其他常见种很少出现在同一样方中。
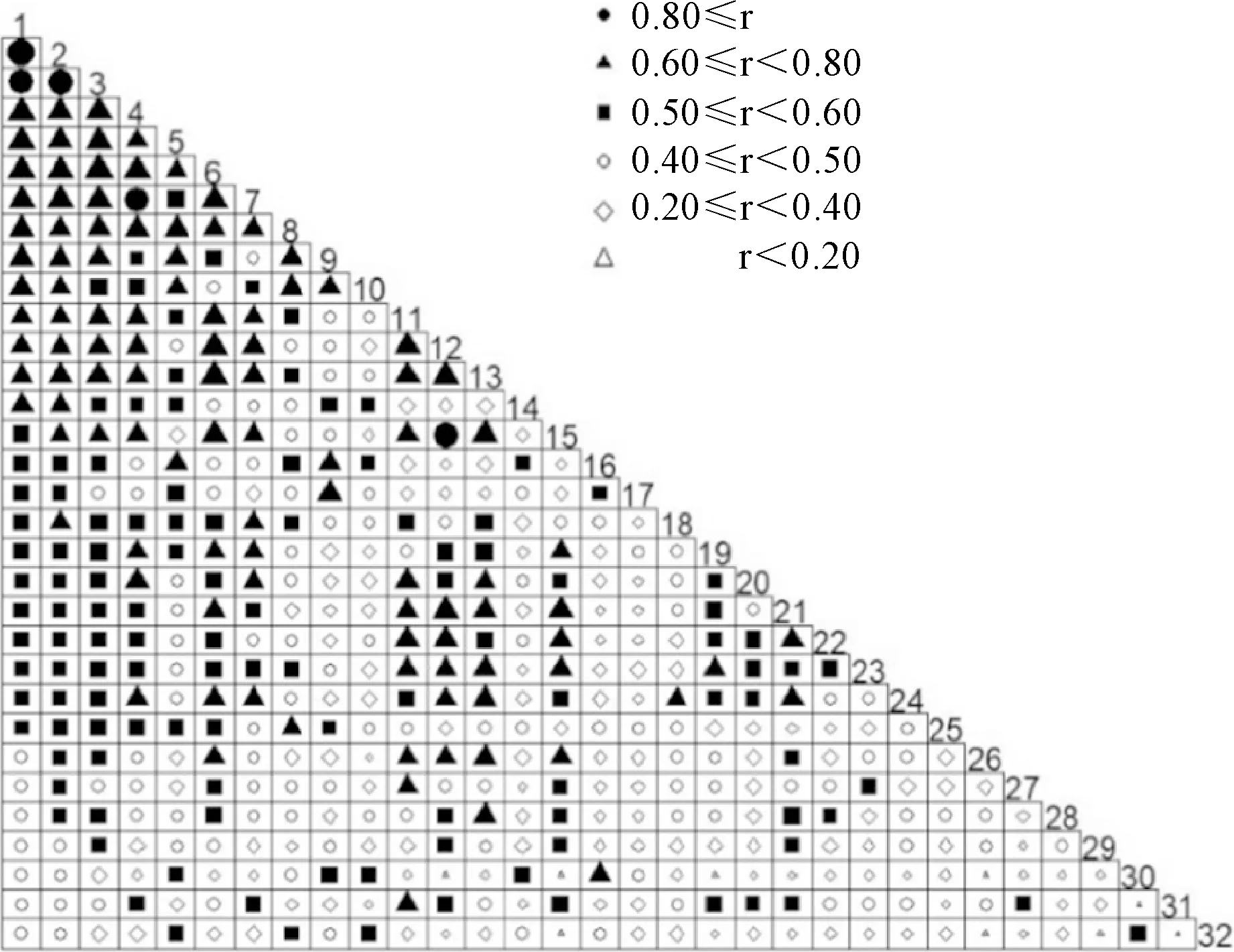
数字1到32物种频度递减,32表示连翘图7 种间关联分析Numbers 1 to 32 indicate decreasing frequency of species, and 32 means F. suspensaFig.7 Interspecific association analysis
3 讨 论
稀有种对群落物种丰富度分布格局的贡献存在非对等性。稀有种虽然对整个群落功能的重要性可能不很显著[15],但其丰富度对整个群落丰富度的贡献很大[9]。例如,本研究发现稀有种虽然对群落物种丰富度分布格局的贡献小于常见种,但稀有种还是占据了整个群落物种丰富度的30.34%,略低于常见种的33.15%,而群落中维持大量稀有种的主要原因是局域群落之间的迁移与生态位分化,同时也是物种能稳定共存的基础[29]。
去除稀有种后,剩余群落物种仍能解释整个群落物种分布信息的98.7%。Schalkwyk等[12]认为群落总体物种丰富度分布格局不反映稀有种,因此在底栖动物生物学评估过程中往往将稀有种按照一定的方式舍去,认为稀有种只能单纯反映一些随机的影响,会对结果产生干扰,而非提供有价值的信息[30],但去除稀有种也可能会损失许多有价值的信息。例如,本研究发现稀有种多歧沙参可能会对群落的物种丰富度格局产生较大的影响。所以在研究中并不能简单舍去,稀有种与常见种相比其对环境胁迫的耐受性要小[31],如果舍去将无法反映群落是否遭受干扰胁迫,进而对群落稳定评价结果产生不确定性。Mouillot等[32]认为在局域和整体水平,稀有种维持生态系统大多数比较显著的功能特性,特别是比较脆弱的生态功能。同时,群落中稀有种相对于常见种对环境或气候变化的响应速度较慢,所以其丧失速率和迁移速率较高[29],进一步说明在生态系统物种多样性保护方面不能忽略稀有种的重要作用。
群落优势种均为常见种,而包含常见种越多的子群落与整个群落丰富度分布格局的相关性越高,即优势种或常见种是群落物种丰富度分布格局指示者[33],相比较于稀有种,常见种受环境扰动的影响更大,因此可以产生最大的变异性[13],但群落基础物种多样性的减少可能会同时导致微环境条件下分化的物种的遗传多样性降低[34],进而导致群落的遗传多样性下降,所以有学者提出应该优先保护常见种[33]。
群落中物种频度、优势度的排序与物种对群落丰富度格局贡献度的排序存在不一致性。本研究发现对群落物种丰富度分布格局贡献最大的前5个物种无一是优势种,且并不是分布范围越广的物种对群落物种丰富度分布格局贡献越大,相反那些与频度较高物种间关联度低的物种对于群落物种的分布格局贡献较大,但此方法并不能充分解释稀有种多歧沙参贡献较高的原因。补偿死亡率假说又称稀有种优势(rare species advantage)或者常见中劣势(common species disadvantage)认为稀有种具有比常见种更高的种群增长率,从而避免其在局域群落种灭绝[7-8],且Connell等[35]用群落补偿趋势(community compensatory trend,CCT)来描述物种增长率与密度的一种负关系,即群落通常并不会通过牺牲稀有种为代价换取常见种的持续增长,从而避免了稀有种在局域群落中灭亡;Mason等[36]认为环境筛的发生是由于物种是稀有种,其个体数太少而不能完全占有所有生境,但稀有种的产生,可能弥补了群落缺失的某些功能,使群落的功能性状空间分布更加均匀。前人研究结论说明稀有种对群落丰富度格局存在较大贡献。
本研究基于α多样性分析了中条山油松人工林下物种对群落物种丰富度分布格局的贡献,丰富了物种丰富度分布格局的研究,但还存在以下几点不足。本研究发现稀有种多歧沙参对群落物种丰富度分布格局具有较高的贡献,且种间关系、补偿死亡率假说和环境筛作用并不能完全解释稀有种贡献度高的原因,所以今后应对此开展深入研究,以补充本研究的不足;研究生物群落构建的过程中,采用物种丰富度的研究方法无法量化物种在生态策略和生态功能等方面的差异,而且也缺少生物多样性应包含的其他重要信息[37],而基于功能性状及功能多样性的研究方法已成为探索物种共存于生物多样性维持机制的一个新的突破口[38-40];本研究基于数据分析结果发现稀有种对群落物种丰富度分布格局也存在较大的贡献,但仅以种间关联度低来解释并不可靠。所以要得出更有意义的结论还应该综合分析其他关键生物学信息。
