海南蒲桃花芽形态分化过程观测及其生理生化变化特征
2021-09-16张诗艳张毛毛叶清华陈清西
张诗艳,张毛毛,张 涛,叶清华,谢 倩,陈清西
(福建农林大学 园艺学院,福州 350002)
海南蒲桃[Syzygiumcumini(L.) Skeels.]为桃金娘科(Myrtaceae)蒲桃属(Syzygium)常绿乔木,又称乌墨,为热带和亚热带珍贵树种,适应性强,被用作中国海南、云南、广西、广东、福建等地区生物防火林带和观赏绿化树种栽培[1]。海南蒲桃为南方常用行道树,其花量大,结实率高,成熟后大量落果,被踩踏后流出紫黑色汁液,影响路面清洁,因此如何通过调控海南蒲桃花量进而减少挂果量,是亟待解决的问题。
花芽分化是营养生长向生殖生长转化的开端,与开花时间密切相关[2],分化正常与否决定着植物能否健康成花并发育成正常果实[3]。目前,已有玉兰[4]、绦柳[5]等绿化植物花芽形态分化过程划分的研究报道,还有银杏[6, 7]花芽形态分化过程和期间内源激素、碳水化合物和矿质营养变化的研究报道,为花芽分化机理的研究奠定了基础。因此,研究植物花芽分化过程与营养、激素水平,对了解花芽分化过程生理生化变化及指导控制花果量和制定栽培管理措施具有重要意义。
本研究通过对海南蒲桃花芽形态结构及相关生理生化指标的观测,分析海南蒲桃花芽分化过程中形态、营养物质与内源激素特征,以及花芽分化过程及生理生化变化,以期为海南蒲桃花果量调控和栽培管理提供理论依据。
1 材料和方法
1.1 试验材料
供试花序、花芽采自福建农林大学校区的海南蒲桃,树体长势良好,无病虫害。
1.2 试验方法
1.2.1 采样与处理2019年4月3日对长势一致的花序进行挂牌,并进行第一次采样,此后每隔6~8 d采样1次,每次采集花序40个。其中,部分花序直接进行形态结构观测;部分花序分离花芽和花柄,洗净擦干,液氮处理后于-40 ℃冰箱保存,用于生理指标测定。
1.2.2 形态结构观测用精度为0.01 mm的游标卡尺测量花芽的长度和宽度,并在扫描仪(HP G4050)4800 dpi分辨率下采集图片。参照李建霞等[8]的方法稍作修改制作冰冻切片:采用包埋剂包埋新鲜花芽,在冷冻切片机(LEICA CM1950)中-24 ℃冷冻1 h后,切取20 μm薄片,捞片后再展片。用倒置荧光显微镜(LEICA DMI8 物镜×5、×10、×20、×40)进行普通光源镜检,采集图像。
1.2.3 生理指标测定参考《植物生理生化实验原理与技术》[9],采用蒽酮比色法测定花芽和花柄可溶性总糖、淀粉含量,采用考马斯亮蓝G-250法测定蛋白质含量。采用酶联免疫吸附法(ELISA)测定内源激素赤霉素(GA3)、脱落酸(ABA)、吲哚乙酸(IAA)和玉米素(ZT)含量,ELISA检测试剂盒由上海优选生物公司提供。
1.3 数据处理与分析
采用Excel 2016进行数据记录整理与作图,采用DPS 7.5进行方差分析。
2 结果与分析
2.1 海南蒲桃花芽形态分化过程及内、外部形态结构变化特征
2.1.1 花芽形态分化过程及内部结构特征海南蒲桃花芽形态分化过程划分为7个阶段,各阶段对应的日期和花芽内部结构特征如图1所示。(1)花序原基分化期:顶端分生组织细胞体积增大,向上突起呈平滑圆弧形,随着生长锥细胞的分裂和生长,分化形成花序原基In(图1,A)。(2)小花原基分化期:花序原基生长点细胞不断分裂,表面积增大,分化出1~3个突起的小花原基Fl(图1,B)。(3)萼片原基分化期:小花原基中心下凹,四周突出形成萼片原基Se(图1,C)。(4)花瓣原基分化期:萼片原基内侧的小花原基在边缘处形成突起,为花瓣原基Pe(图1,D)。(5)雄蕊原基分化期:花瓣原基内侧产生新的突起,向腔内弯曲,即为雄蕊原基St(图1,E)。(6)雌蕊原基分化期:小花原基的中心部位出现新的突起为雌蕊原基Pi(图1,F)。(7)雄蕊雌蕊形成期:雌蕊原基高度开始超过雄蕊原基,雄蕊雌蕊原基细胞继续进行生长分化,雄蕊原基顶端分化形成花药,基部形成花丝;雌蕊原基向上分化形成花柱和柱头,向下在花托中形成子房(图1,G~I)。至此,海南蒲桃花芽分化基本完成。
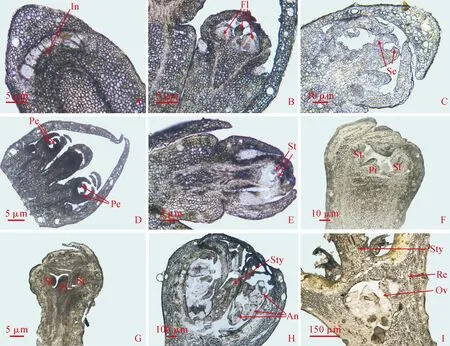
A.花序原基分化期(4月3日);B.小花原基分化期(4月3日-4月11日);C.萼片原基分化期(4月11日-4月18日);D.花瓣原基分化期(4月18日-4月25日);E.雄蕊原基分化期(4月25日-5月2日);F.雌蕊原基分化期(5月2日-5月8日);G~I.雄蕊雌蕊形成期(5月8日-6月2日)。In.花序原基;Fl.小花原基;Se.萼片原基;Pe.花瓣原基;St.雄蕊原基;Pi.雌蕊原基;An.花药;Sty.花柱;Ov.子房;Re.花托图1 海南蒲桃花芽形态分化过程A. Inflorescence primordium differentiation period(April 3); B. Floret primordium differentiation period(April 3 - April 11); C. Sepal primordium differentiation period(April 11 - April 18); D. Petal primordium differentiation period(April 18 - April 25); E.Stamen primordium differentiation period(April 25 - May 2); F. Pistil primordium differentiation period(May2 - May 8); G~I. Stamen and pistil formation period(May 8 - June 2). In. Inflorescence primordium; Fl. Floret primordium; Se. Sepal primordium; Pe. Petal primordium; St. Stamen primordium; Pi.Pistil primordium; An.Anther; Sty. Style; Ov. Ovary; Re. Receptacle.Fig.1 Flower bud morphological differentiation of S. cumini
2.1.2 花芽外部形态的变化特征海南蒲桃花序芽位于花序轴各分枝的顶端,花序芽又可分化形成3个小花芽,位于中间的为顶芽,两侧为侧芽,整个花序呈圆锥状。从2019年4月3日到6月2日,海南蒲桃花芽从花序原基形成到分化结束的时间跨度为60 d。期间,花芽大小和形状随着分化的进行发生了明显变化(图2);顶芽和侧芽的长宽变化趋势基本一致,花芽长度均呈“快—慢”趋势增加,宽度均呈“快—慢—快”趋势增加(图3)。具体变化为:4月3日,处于花序原基分化期的海南蒲桃花序轴各分枝的顶端为单一花序芽(图2,A);4月11日,小花原基分化期的花序轴各分枝伸长,花序芽已经分化形成1~3个小花芽(图2,B);4月18日,萼片原基分化导致花芽上部较花托处明显膨大,成为萼筒(图2,C);4月25日,处于花瓣原基分化期的花芽其纵切面出现空腔(图2,D),颜色较浅;5月2日,处于雄蕊原基分化期的花芽纵切面显现2片深色(图2,E 箭头所示),为雄蕊形成部位;5月8日,处于雌蕊原基分化期的花芽中轴部位形成淡绿色花柱(图2,F);5月16日和5月22日,雄蕊雌蕊形成期之间花芽越来越饱满(图2,G~H);6月2日,花芽明显变黄,形成完整的雌雄蕊,部分花芽发育到即将开花的状态,花丝顶开上部覆盖的圆形帽状体(图2,I 箭头所示),此时顶芽和侧芽的宽度均显著增加,表明花芽完成分化即将开放时会横向膨大。

图2 海南蒲桃花芽分化过程中外部形态的变化Fig.2 Changes of the external morphology of flower buds of S. cumini during its differentiation
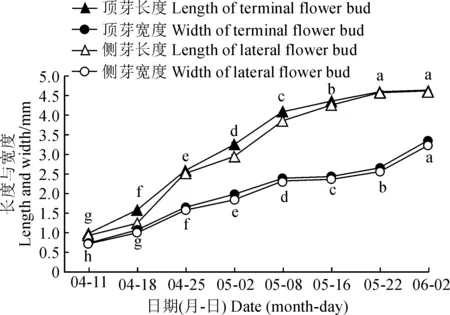
此处所测花芽长宽为小花原基分化期后分化出的小花芽的长宽。不同小写字母表示在0.05水平上差异显著。下同图3 海南蒲桃花芽分化过程中长度和宽度的变化Length and width measured are the length and width of flower buds after the floret primordium differentiation period. Different letters indicate significant differences at 0.05 level. The same as belowFig.3 Length and width of flower buds of S. cumini during its differentiation
2.2 海南蒲桃花芽形态分化过程中营养物质含量变化特征
首先,海南蒲桃花芽形态分化过程中,花芽可溶性糖含量呈先下降后上升的变化趋势,花柄可溶性糖含量呈先下降后上升再下降趋势(图4,A)。其中,4月3日至4月25日,花芽和花柄可溶性糖含量都显著降低,分别达到最低值4.47 mg/g和3.88 mg/g,说明从小花原基开始分化到花瓣原基分化完成,花芽和花柄均需消耗大量可溶性糖;自5月16日起,花芽可溶性糖含量显著上升并于6月2日达最高水平(11.78 mg/g),花柄可溶性糖含量则显著下降,说明花芽形态分化基本完成时,花芽积累大量可溶性糖有利于开花。
其次,海南蒲桃花芽形态分化过程中,花芽淀粉含量呈先下降后上升趋势,花柄淀粉含量则呈先上升后下降再上升趋势(图4,B)。其中,4月11日至4月25日,花芽淀粉含量处于较低水平,花柄淀粉含量显著上升,并在4月25日达到最高值(9.33 mg/g);4月25日至5月8日,花芽淀粉含量显著上升,而花柄淀粉含量显著下降,并于5月8日达到最低值(4.70 mg/g);之后,花芽淀粉含量先降后升,花柄淀粉含量则持续显著上升,均在6月2日达到最高水平,此时花芽形态分化基本完成。说明海南蒲桃花芽形态分化前期和雄蕊雌蕊形成期,花芽大量消耗积累的淀粉,而花芽进入雄蕊雌蕊形成期和开花需要积累淀粉。
另外,海南蒲桃花芽形态分化过程中,花芽蛋白质含量呈先上升后下降趋势,花柄蛋白质含量总体呈下降趋势(图4,C)。到其花芽形态分化基本完成时(6月2日),花柄蛋白质含量处于极低水平,而花芽蛋白质却大量积累。
2.3 海南蒲桃花芽形态分化过程中内源激素含量及比值的变化特征
2.3.1 GA3、ABA、IAA、ZT含量在整个海南蒲桃花芽形态分化过程中,花芽和花柄GA3含量分别在0.56~0.68 ng/g和0.58~0.71 ng/g间变化,GA3含量始终处于较低水平且较为稳定(图5,A)。
花芽和花柄ABA含量均呈近似“W”型变化趋势(图5,B)。在雄蕊原基和雌蕊原基分化期间,花芽ABA含量于5月2日达到最高值(915.33 ng/g),花柄ABA含量于5月8日达到最高值(801.53 ng/g)。
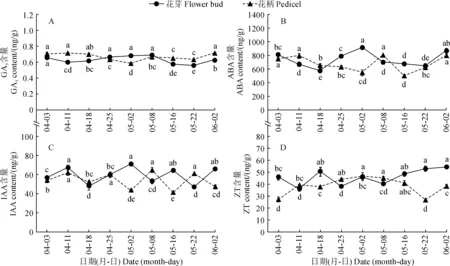
图5 海南蒲桃花芽和花柄内源激素含量的变化Fig.5 Changes of endogenous hormone contents in flower buds and pedicels of S. cumini during its differentiation
花芽和花柄IAA含量分别在47.15 ~71.05 ng/g和40.98 ~64.88 ng/g间波动变化,每次波动幅度都达到显著水平;花芽和花柄IAA含量的变化趋势从4月3日至4月25日几乎同步,自4月25日进入雄蕊原基分化期后,两者的变化趋势相反(图5,C)。
花芽ZT含量先波动变化再缓慢上升,而花柄ZT含量呈先上升后下降再上升趋势;相较分化之初,花芽形态分化基本完成时花芽和花柄ZT含量均显著提高,其中花芽ZT含量达到最高值54.27 ng/g(图5,D)。
2.3.2 花芽内源激素比值由表1可知,在海南蒲桃花芽分化过程中,花芽ABA/GA3比值较大,始终远高于同期其他内源激素比值,并呈先下降后上升、再下降再上升的变化趋势,与ABA含量变化趋势相同,且分化前后差异显著;花芽ABA/IAA比值相对较低,并也呈先下降后上升、再下降再上升的趋势,且变幅相对较小;花芽ZT/GA3比值处于较高水平,并呈波动变化,在雄蕊雌蕊形成期期间(5月22日)达到最大值为94.38,与其他时期花芽ZT/GA3比值差异显著;花芽ZT/IAA比值在各比值中最小,呈较小的波动变化趋势,且分化前后ZT/IAA比值水平无显著差异。以上结果表明,海南蒲桃花芽形态分化过程中,花芽ABA/GA3和 ZT/GA3比值均显著增大,ABA/IAA比值稍有减小。
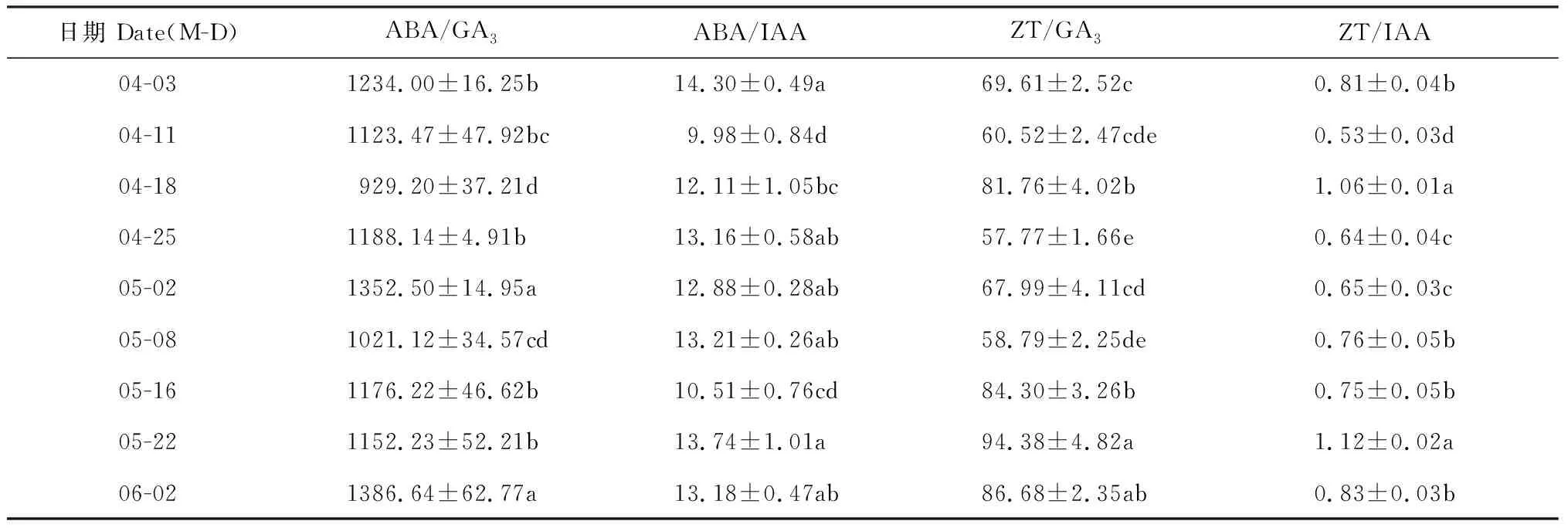
表1 海南蒲桃花芽分化过程中内源激素比值的变化
3 讨 论
3.1 海南蒲桃花芽形态分化阶段的划分
植物花芽形态分化过程的划分没有明确的标准,但多以花芽内各原基出现的顺序作为划分依据,分化顺序一般为萼片、花瓣、雄蕊和雌蕊[10]。海南蒲桃为花序型植物,且小花为两性花,花芽形态分化过程被划分为7个阶段,与馥郁滇丁香‘香妃’[11]、蓝花丹[12]等花芽形态分化规律基本相似,明显与单生花型植物如杏李[13]和雌雄异株型植物如栝楼[14]的花芽形态分化规律存在差异。乔永刚等[15]认为,花芽分化过程中花芽外形与内部结构存在相关性,相邻分化阶段存在一定的重叠现象。本研究中,海南蒲桃花芽内外形态结构也存在对应关系,并据此划分了各阶段发生的时间,发现萼片原基分化期与花瓣原基分化期、雄蕊原基分化期与雌蕊原基分化期在时间上也存在部分重叠现象。花芽形态建成是非常复杂的过程,花芽形态分化规律不仅取决于植物自身遗传性,还会受外界环境条件的影响,如光周期、温度和气候等[16-17],树冠上不同部位的花芽受树体营养分配规律的影响[18],也会导致分化时间上存在差异。
3.2 海南蒲桃花芽形态分化与营养物质含量的关系
碳水化合物是花芽分化过程中重要的营养物质和能量来源[19]。张绿萍等[20]研究发现,莲雾花芽分化过程中叶片碳水化合物含量显著下降,认为可能是莲雾花芽分化消耗所致。本研究中,海南蒲桃花芽和花柄碳水化合物含量在分化过程中都显著升高,由此推测海南蒲桃花芽形态分化和花柄生长发育消耗的碳水化合物主要由叶片输入补充,这同时也体现了花序与周围叶片为“库”和“源”的关系,“源”为“库”提供营养物质[21]。本研究发现,海南蒲桃花芽形态分化初期和雄蕊雌蕊形成期,花芽可溶性糖和淀粉含量较高,这与张丽杰等[22]研究结果一致,说明花芽进入形态分化和开花都需要积累大量的碳水化合物。
蛋白质是细胞增殖、形态建成和生命活性的物质基础,是花芽分化过程中必需的结构物质和营养物质[23]。植物花芽分化过程中蛋白质含量增加[24],为花芽分化提供物质保障。海南蒲桃花芽形态分化过程中花芽、花柄的蛋白质含量变化呈现互补趋势,因此推测海南蒲桃花芽需要从花柄中转运大量蛋白质,而花柄发育只需要较低水平的蛋白质。海南蒲桃花芽分化的顺利进行和开花需要积累大量蛋白质,主要用于各原基形态结构的维持。
3.3 内源激素对海南蒲桃花芽形态分化的调控
植物花芽分化过程受到多种激素的调控,一般认为赤霉素对花芽分化起抑制作用,细胞分裂素对花芽分化起促进作用,脱落酸和生长素对花芽分化的作用没有统一认识[25]。莫文娟等[13]研究认为,高水平的ABA可能引起枝条停长,与GA3形成拮抗,有利于花芽分化;张宁等[26]研究发现,高水平的ZT促进花芽分化。本研究结果显示,海南蒲桃花芽形态分化过程中,花芽GA3含量低且较稳定,ABA、ZT含量较高,这与前人研究结果一致,因此认为高水平的ZT、ABA和低水平GA3有利于海南蒲桃的花芽形态分化。
同时,花芽形态分化并非由单一激素决定,激素间存在着互相协同和拮抗的作用,是一个复杂的调控过程[19]。赵通等[27]研究发现‘李光杏’花芽分化过程中花芽内ABA/IAA、ZR/IAA及ABA/GA的比值升高;覃喜军等[28]研究发现罗汉果花芽分化过程中花芽内ABA/IAA、ZR/IAA比值稳定,认为高比值的ABA/GA3、ZR/GA3有利于其花芽分化。本研究中高水平的ABA/GA3、ZT/GA3也有利于海南蒲桃花芽形态分化的进行。
4 结 论
本研究将海南蒲桃花芽形态分化过程划分为7个阶段:即花序原基分化期、小花原基分化期、萼片原基分化期、花瓣原基分化期、雄蕊原基分化期、雌蕊原基分化期和雄蕊雌蕊形成期。海南蒲桃花芽形态分化和开花均需要积累大量的可溶性糖、淀粉和蛋白质,低含量的GA3,高含量的ZT、ABA、IAA及高水平的ABA/GA3、ZT/GA3比值有利于海南蒲桃花芽形态分化。因此,在海南蒲桃花果量调控和栽培管理中,可以考虑外施GA3来打破花芽GA3的稳定水平,降低ABA/GA3、ZT/GA3比值,达到抑制海南蒲桃花芽分化的效果。
