基于盐胁迫条件下施用氮钾肥对面包果养分吸收及渗透物质积累的影响
2021-09-14苏兰茜白亭玉赵顺松吴刚谭乐和
苏兰茜 白亭玉 赵顺松 吴刚 谭乐和
摘 要:为探究不同氮钾配比影响盐胁迫下面包果生长的营养生理机制,以面包果[Artocarpus altilis (Parkinson) Fosberg]为材料,研究不同氮钾配比对盐胁迫下面包果养分吸收及渗透物质的影响。结果表明:与阴性对照(N0K0+W)相比,单盐胁迫(N0K0+S)对面包果生长指标有一定抑制作用,對根系生长指标有一定促进作用,其中N1K1+S处理的面包果地下部干重较单盐胁迫处理呈增加趋势,根系生长指标呈下降趋势。所有盐胁迫处理均显著增加根茎叶中的Na+含量和Na+/K+。与阳性对照(N0K0+S)相比,N1K1+S处理显著增加根茎叶中K+含量,减少叶中Na+含量和Na+/K+。N1K2+S处理较N1K1+S处理显著减少根茎中的Na+含量,N2K2+S处理较N2K1+S处理显著增加根中的N、P、K+、Na+含量和Na+/K+,茎中的N和K+含量以及叶片中的Na+含量、Na+/K+和叶片可溶性糖含量。N2K1+S处理较N1K1+S处理显著减少根、茎中K+含量,增加叶片N、Na+含量和Na+/K+,降低叶片可溶性糖含量,增加脯氨酸含量。N2K2+S处理较N1K2+S处理显著增加根、茎、叶中的Na+含量和Na+/K+。综上所述,适量增施氮钾肥通过调节K+、Na+向各器官的运输缓解盐胁迫对面包果生长及养分吸收的抑制效应,可为盐碱地的面包果施肥提供参考。
关键词:面包果;Artocarpus altilis (Parkinson) Fosberg;盐胁迫;养分吸收;渗透物质
Abstract: In order to explore the nutritional and physiological mechanism of nitrogen and potassium on the growth of breadfruit under salt stress, this experiment used breadfruit seedlings [Artocarpus altilis (Parkinson) Fosberg] as the test materials to compare the effects of different nitrogen and potassium application on the nutrient absorption and osmotic substances of breadfruit seedlings. Compared with the negative control (N0K0+W), single salt stress (N0K0+S) had an inhibitory effect on breadfruit growth indices and a promotion effect on root growth indices. Among them, the root dry weight under N1K1+S treatment showed an increasing trend, and the root growth indices showed a decreasing trend compared with the single salt stress treatment. All salt stress treatments significantly increased the content of Na+ and Na+/K+ in roots, stems and leaves. Compared with the positive control (N0K0+S), N1K1+S treatment significantly increased the content of K+ in roots, stems and leaves, and decreased the content of Na+ and Na+/K+ in leaves. Compared with N1K1+S, the treatment N1K2+S significantly reduced the content of Na+ in in roots and stems. N2K2+S treatment significantly increased the contents of N, P, K+, Na+ and Na+/K+ in roots, N and K+ in stems, and the contents of Na+, Na+/K+ and soluble sugar in leaves compared with N2K1+S treatment. N2K1+S treatment significantly decreased the content of K+ in roots and stems, increased the contents of N, Na+ and Na+/K+ in leaves, decreased the soluble sugar content in leaves, and increased proline content compared with N1K1+S treatment. N2K2+S treatment significantly increased the content of Na+ and Na+/K+ value in roots, stems and leaves compared with N1K2+S treatment. In summary, proper application of nitrogen and potassium fertilizer could alleviate the inhibitory effect of salt stress on the growth and nutrient absorption of breadfruit by regulating the transport of K+ and Na+ to organs. The results could be used as a reference for fertilization of breadfruit in saline soil.
Keywords: breadfruit; Artocarpus altilis (Parkinson) Fosberg; salt stress; nutrient absorption; osmotica
土壤盐碱化是威胁作物产量和农业生产的全球化问题[1-2]。据联合国粮食及農业组织2012年的数据库估计,全球受盐碱化影响的土地超过8亿 hm2,目前盐碱化仍在扩大,对农业的可持续发展构成严重威胁[3]。我国滨海盐碱地面积较大,但苛刻的土壤条件严重限制了盐碱地的开发利用。例如,近年来西沙各岛发展迅速,驻岛官兵和渔民的粮食以及饮用水资源供应面临极大不便,又因岛上土地盐分高,淡水资源紧缺,发展岛礁农业面临巨大困难[4]。面包果是热带特色木本粮食作物,果肉含丰富的蛋白质、碳水化合物及膳食纤维,可替代粮食,前期研究发现面包果对低盐胁迫有一定的耐受力,但其能否在滨海盐碱地正常生长有待后续探究。提高面包果在高盐胁迫下的耐受力,有利于促进滨海、岛礁土地资源的开发利用和种植业的有效发展,对于保障交通不便的滨海和岛礁居民生活具有重要现实意义。
盐渍土中过多的盐离子易导致植物生理代谢紊乱[5-6],影响植物的生长发育和形态特征,对植物造成离子毒性、营养缺乏和渗透胁迫[7]。作物的耐盐性与养分管理密切相关,盐碱环境下的施肥尤其是氮钾肥的施用量及使用方式越来越引起人们的重视。氮素对作物的许多生理过程、抗逆性和其他营养元素的利用均有重要的调节作用[8-9],其通过促进脯氨酸、可溶性糖等渗透调节物质的积累进而提高植物渗透调节能力[10]。施用氮肥可增加原生质体水合度,提高原生质保水能力,通过影响体内一些重要的酶来控制植物的生理生化代谢。在盐胁迫下,钾素能调控细胞内的离子平衡以及调控细胞膨压,在钠离子高聚集的环境中,钾离子能竞争性地抑制钠离子进入植物体内,从而减缓钠离子对植物的生长造成渗透胁迫和离子毒害[11]。前期研究发现面包果有一定的耐盐性,但在其耐盐阈值范围内如何通过养分管理进一步提高耐盐性有待进一步研究。本研究拟通过分析不同氮钾配比对盐胁迫下面包果养分吸收及渗透物质的影响,明确氮钾配比促进盐胁迫下面包果生长的营养生理机制,不仅能为面包果生长提供必需的养分,同时能缓解盐胁迫效应,为热带盐碱地的开发利用,发展岛礁特色农业(热带木本粮食作物)提供理论依据。
1 材料与方法
1.1 材料
供试面包果苗为中国热带农业科学院香料饮料研究所自主选育的4号品种。
供试海水为石梅湾海水,用电导盐度计测定海水盐度为34.0‰。
供试土壤采自中国热带农业科学院香料饮料研究所面包果种植基地(18°15′ N;110°13′ E),该地区属热带海洋性季风气候,年均降水2 135 mm,土壤类型为砖红壤,pH 6.62,速效磷75.05 mg/kg,速效钾402.78 mg/kg,碱解氮77.10 mg/kg,有机质27.2 g/kg。
1.2 方法
1.2.1 试验设计 盆栽试验于2019年10月— 2020年4月在中国热带农业科学院香料饮料研究所温室进行,采用单因素完全随机设计。根据前期的耐盐性试验结果,当盐分浓度为10.2 g/kg时,面包果叶片光合参数显著下降,当盐浓度继续增加时,各参数值与该浓度时的值无显著差异[4],故本研究设置盐分浓度为10.2 g/kg。预实验结果发现,含高氮、高钾处理(施氮量、施钾量在中氮、中钾的基础上增加60%)的面包果苗生长受阻,表现出明显的叶片黄化脱落现象,因此本研究取消高氮、高钾浓度处理,设置为低氮低钾、低氮中钾、中氮低钾、中氮中钾、无氮无钾浇海水(阳性对照)以及无氮无钾浇灌溉水(阴性对照),共计6个处理,不同处理的施肥量见表1。其中,中氮中钾处理的施肥量是根据前期苗期氮钾最佳施肥量确定,低氮、低钾处理的施氮量、施钾量分别在中氮、中钾的基础上减少60%,每处理设置3个重复,每重复3株,总计54株。每盆(36 cm×15 cm)装土16 kg,N0K0+W处理用灌溉水配置1/2 Hoagland营养液(已经去掉氮钾素),N0K0+S处理用10.2 g/kg的海水配置1/2 Hoagland营养液(无氮钾素)。施肥处理中,氮源用硝酸钠,钾源用氯化钾。试验过程中,为防止面包果产生盐激反应,本研究采用渐增的方式(每天增加3.4 g/kg)达到预设浓度。通过称重法将土壤水分维持在田间持水量的75%左右,于每天18:00用五点法采集土样测定含水量,及时补水,每5天浇一次营养液。
1.2.2 面包果生长指标测定 种植3个月后开始测定各个指标,每个处理随机选取6株长势较一致的面包果苗测定生长指标,苗高为茎基部到叶片最高点的高度;茎粗用游标卡尺测定,统一测定距茎基部2 cm处。植物样品收获后,用去离子水将植株冲洗3次,分成根、茎、叶3部分,置于105 ℃烘箱中杀青30 min,转至65 ℃烘至恒重,并称其干质量。采用WinRhizo 软件分析根系形态指标[12]。
1.2.3 面包果各器官离子含量测定 各器官离子含量采用《土壤农化分析》[13]的方法测定。
1.2.4 面包果叶片渗透物质含量测定 叶片可溶性糖含量采用蒽酮比色法测定,脯氨酸含量采用酸性茚三酮法测定。上述指标测定均参照试剂盒说明书进行,试剂盒购自南京建成生物工程研究所。
1.2.5 土壤理化性质测定 土壤样品理化性质参照《土壤农化分析》[13]进行测定。
1.3 数据分析
采用SPSS 19.0软件进行数据分析,单因素方差分析(ANOVA)进行数据比较,利用Duncan新复极差法检验处理间的差异显著性。
2 结果与分析
2.1 施用氮钾肥对盐胁迫下面包果幼苗生长的影响
从表2可以看出,与阴性对照(N0K0+W)相比,单盐胁迫(N0K0+S)对面包果株高、茎粗、地下部干重和总干重有一定抑制作用,但差异不显著;N1K1+S处理显著减少株高的比例为12.24%;N1K2+S处理显著减少株高、地下部干重的比例分别为18.19%和26.65%;N2K1+S处理显著减少株高、茎粗的比例分别为15.51%和14.70%;N2K2+S处理显著减少株高、总干重的比例分别为18.52%和18.04%。面包果的干物质量在各施肥处理与阳性对照(N0K0+S)中大致呈先增加后降低的趋势,但差异不显著,其中N1K1+S处理中面包果地下部干重较阳性对照有增加趋势,地下部干重在N1K1+S处理中显著高于N1K2+S处理。
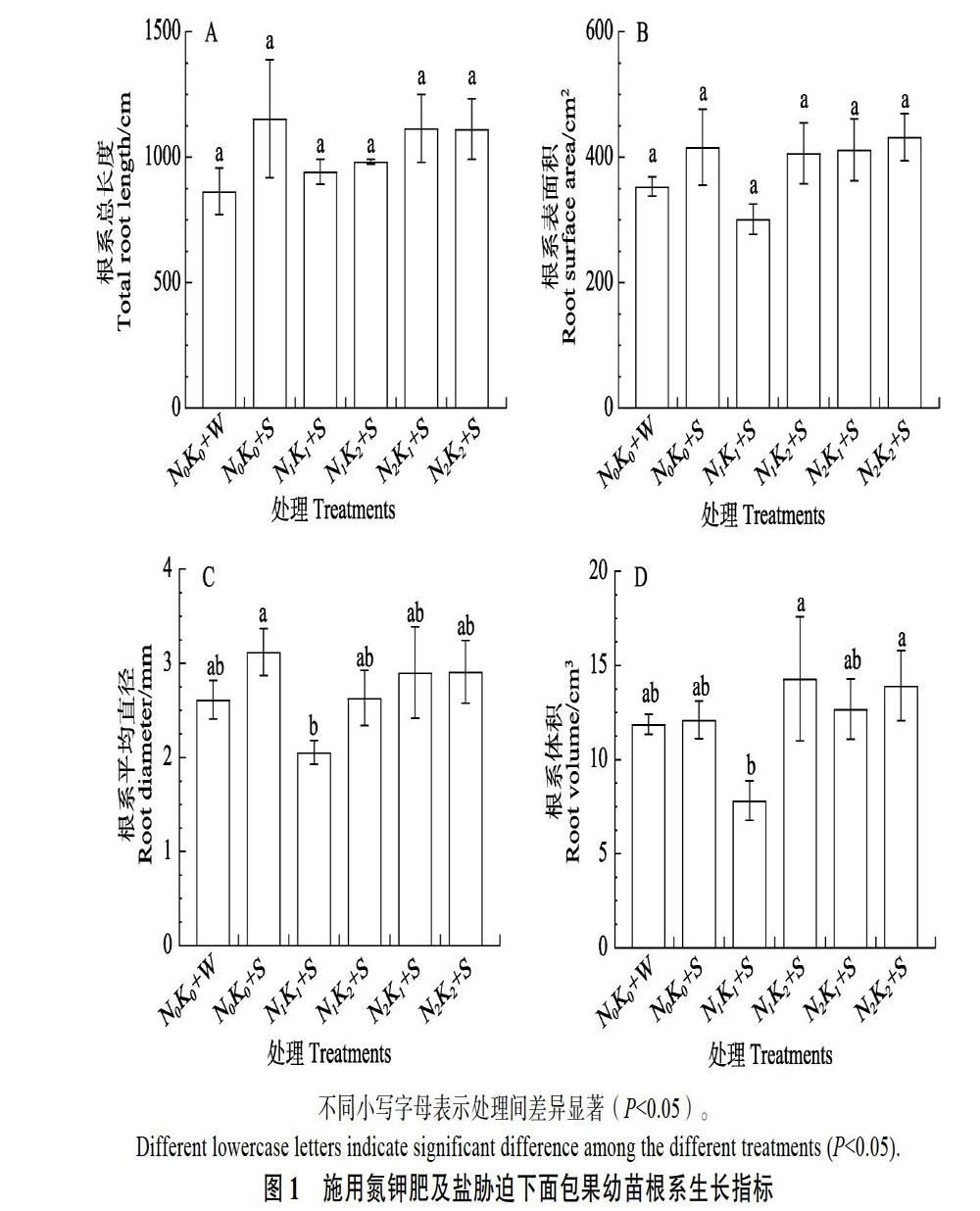
2.2 施用氮钾肥对盐胁迫下面包果根系生长指标的影响
由图1可以看出,与阴性对照(N0K0+W)相比,根系生长指标在单盐胁迫处理中均呈增长趋势。不同处理对面包果根系总长度、表面积的影响无显著性差异,但在N1K1+S处理中呈下降趋势(图1A,图1B)。N1K1+S处理较阳性对照(N0K0+S)显著减少根系平均直径的比例为34.13%(图1C),较N1K2+S处理显著减少根系体积的比例为45.23%(图1D)。随着施肥量的增加,根系生长指标呈一定增长趋势。
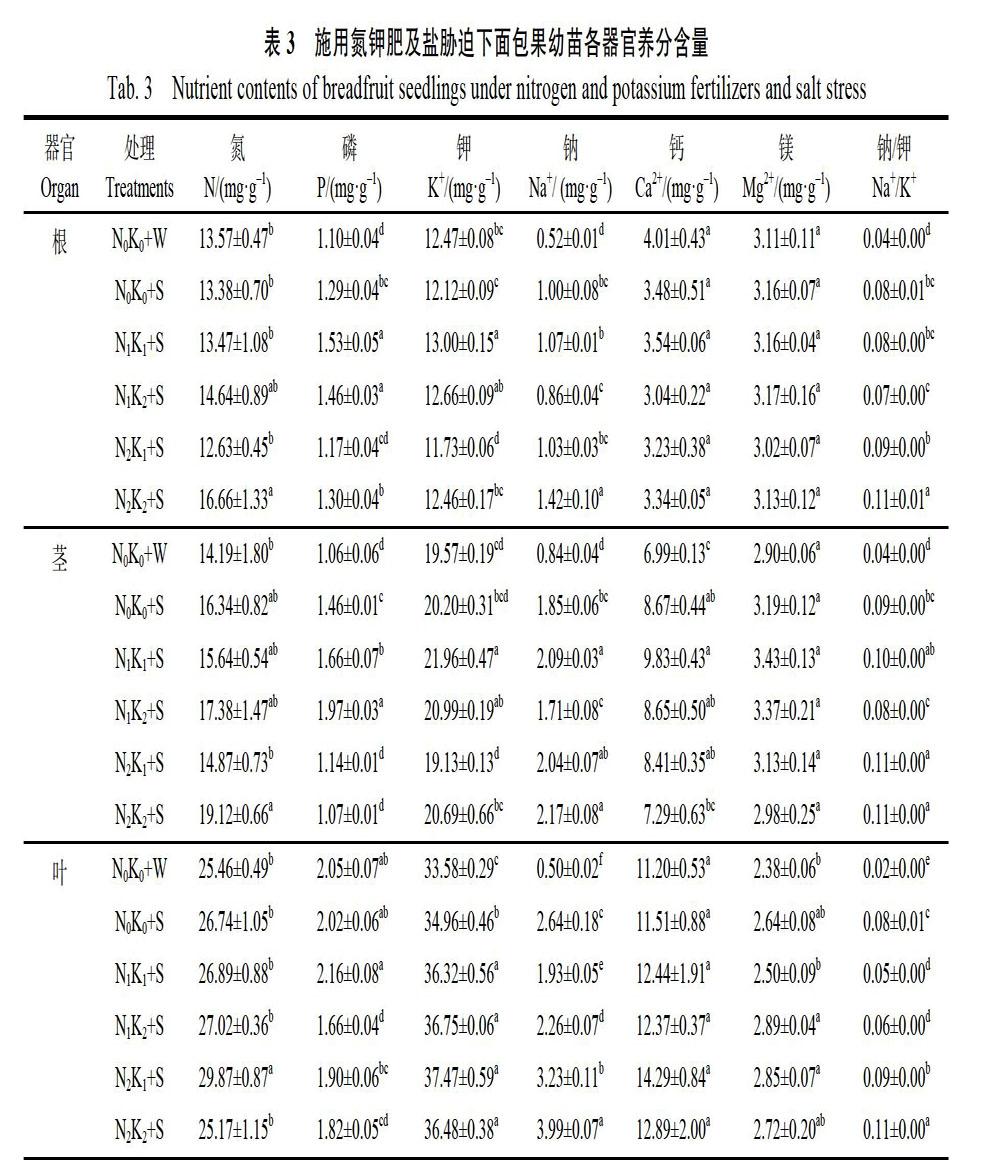
2.3 施用氮钾肥对盐胁迫下面包果各器官离子含量的影响
由表3可见,氮磷钾素在不同处理面包果各器官中的含量大致表现为叶片>茎秆>根系。与阴性对照(N0K0+W)相比,N0K0+S处理显著增加根中P、Na+含量和Na+/K+的比例分别为17.27%、92.31%和100%,增加茎中P、Na+、Ca2+含量和Na+/K+的比例分别为37.74%、120.24%、24.03%和125%,增加叶片中K+、Na+含量和Na+/K+的比例分别为4.11%、428%和300%;N1K1+S显著增加根中P、K+、Na+含量和Na+/K+的比例分别为39.09%、4.25%、105.77%和100.00%,增加茎中P、K+、Na+、Ca2+含量和Na+/K+的比例分别为56.60%、12.21%、148.81%、40.63%和150.00%,增加叶中K+、Na+含量和Na+/K+的比例分别为8.16%、286.00%和150.00%;N1K2+S顯著增加根中P、Na+含量和Na+/K+的比例分别为32.73%、65.38%和75.00%,增加茎中P、K+、Na+、Ca2+含量和Na+/K+的比例分别为85.85%、7.26%,103.57%、23.75%和100.00%,增加叶中K+、Na+、Mg2+含量和Na+/K+的比例分别为9.44%、352.00%、21.43%和200%,减少叶中P含量的比例为19.02%;N2K1+S显著增加根中Na+含量和Na+/K+的比例分别为98.08%和125.00%,减少根中K+含量的比例为5.93%,增加茎中Na+、Ca2+含量和Na+/K+的比例分别为142.86%、20.31%和175.00%,增加叶中N、K+、Na+、Mg2+含量和Na+/K+的比例分别为17.32%、11.58%、546.00%,19.75%和350.00%;N2K2+S显著增加根中N、P、Na+含量和Na+/K+的比例分别为22.77%、18.18%、173.08%和175.00%,增加茎中N、Na+含量和不同小写字母表示处理间差异显著(P<0.05)。
Na+/K+的比例分别为34.74%、158.33%和175.00%,增加叶中K+、Na+含量和Na+/K+的比例分别为8.64%、698.00%和450.00%,减少叶中P含量的比例为11.22%。与阳性对照(N0K0+S)相比,N1K1+S显著增加根中P、K+含量的比例分别为18.60%和7.26%,增加茎中P、K+、Na+含量的比例分别为13.70%、8.71%和12.97%,增加叶中K+含量的比例为3.89%,减少叶中Na+含量和Na+/K+的比例分别为26.89%和37.50%;N1K2+S显著增加根中P、K+含量的比例分别为13.18%和4.46%,增加茎中P含量的比例为34.93%,增加叶中K+含量的比例为5.12%,减少叶中P、Na+含量和Na+/K+的比例分别为17.82%、14.39%和25.00%;N2K1+S显著减少根中K+含量的比例为3.22%,增加茎中Na+/K+的比例为22.22%,减少茎中P含量的比例为21.92%,增加叶中N、K+、Na+含量和Na+/K+的比例分别为11.71%、7.18%,22.35%和12.50%;N2K2+S显著增加根中N、Na+含量和Na+/K+的比例分别为24.51%、42.00%和37.50%,增加茎中Na+含量和Na+/K+的比例分别为17.30%和22.22%,减少茎中P含量的比例为26.71%,增加叶中K+、Na+含量和Na+/K+的比例分别为4.35%、51.14%和37.50%,减少叶中P含量的比例为9.90%。
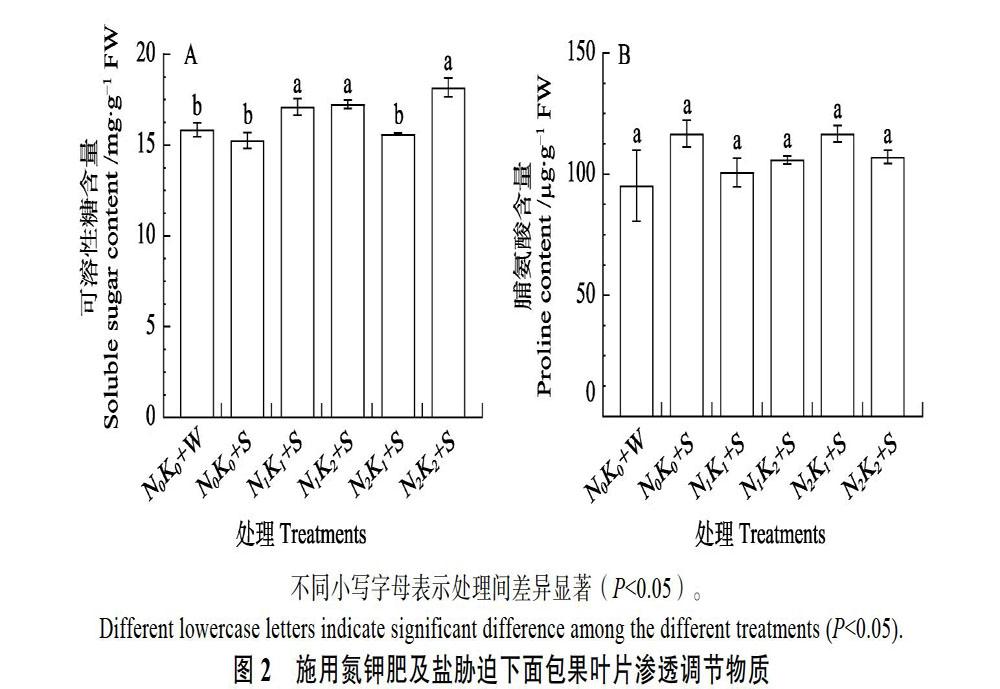
2.4 施用氮钾肥对面包果叶片渗透调节物质含量的影响
由图2A可知,与阴性对照(N0K0+W)相比,叶片中的可溶性糖含量在N0K0+S处理中呈轻微上升趋势,但差异不显著;N1K1+S、N1K2+S、N2K2+S处理显著增加叶片可溶性糖含量的比例分别为7.87%、8.77%和14.67%。与阳性对照(N0K0+S)相比,N1K1+S、N1K2+S、N2K2+S处理显著增加叶片可溶性糖含量的比例分别为12.11%、13.05%和19.18%。N2K1+S处理的叶片可溶性糖含量与两对照相比无显著差异。与阴性对照相比,叶片脯氨酸含量在单盐胁迫处理中呈上升趋势,但对照和各施肥处理无显著性差异(图2B)。
3 討论
盐胁迫会打破植物体内的离子平衡,造成生理功能紊乱,最明显的表现是植物生长受抑制,生物量下降[14]。本研究结果表明,盐胁迫下面包不同小写字母表示处理间差异显著(P<0.05)。
果生长指标呈下降趋势,但与浇灌溉水处理相比差异不显著,这与之前的研究结论一致,面包果对低盐胁迫具有一定的耐受力[4]。研究表明,对于盐生植物来说,盐分浓度在没有超过生长阈值时,同化作用不会被抑制,甚至在较低时还会被促进[15]。增施氮钾肥并没有表现出明显的促生效果,这可能与面包果自身的耐盐性有关。氮钾养分对盐胁迫下面包果的生长指标没有表现出明显的可塑性,说明生长指标对面包果盐胁迫下的吸N、K能力的贡献不大,因此不能作为氮钾提高面包果耐盐性的生长筛选指标。低浓度氮钾素添加对面包果生物量有一定促进作用,这与康爱平等[16]在能源植物杂交狼尾草耐盐性的研究结果相似,钾素对盐胁迫下的植株生长表现出一定的缓解效应。
根系是最先感应盐胁迫的植物器官,根系的生长发育、形态变化与环境胁迫密切相关。研究表明,不同作物对盐分的敏感程度不同,低盐胁迫对部分稀盐盐生植物的根系生长发育有一定促进作用[17],而对甜土植物和农作物的根系有明显抑制作用或作用不显著[18-19]。本研究结果表明氮钾素添加虽对盐胁迫下面包果根系生长指标作用效果不显著,但仍然可以看出一定的变化趋势。盐胁迫下面包果根系生长指标呈一定增长趋势,与浇灌溉水处理差异不显著,可见适当的盐分可促进面包果根系生长,而增施氮钾肥处理的根系指标呈先降低后增加的趋势,表明氮钾素添加对盐胁迫下的面包果根系生长表现出一定的剂量效应。研究表明,增加根区供氮水平,能显著提高高羊茅[20]和盐地碱蓬[21]抵抗盐胁迫的能力。
适量的氮肥有助于补偿和纠正因盐胁迫条件下所引起的离子失衡[22]。在盐胁迫下,限制Na+的积累及K+的外流可以减缓钠离子对植物的生长造成渗透胁迫和离子毒害[23],增强植物的耐盐能力[24]。盐胁迫条件下,植株体Na+含量的显著增加抑制了K+的吸收和积累,Na+/K+显著增加[25]。本研究结果也发现,盐胁迫条件下面包果根茎叶中的Na+含量和Na+/K+均显著高于非盐胁迫。与N1K1+S处理相比,N1K2+S处理能加快Na+从根茎向叶中运输,与N2K1+S处理相比,N2K2+S处理显著促进根系吸收K+和茎中K+含量的积累,同时也促进了根系吸收Na+和叶片中Na+的积累。N2K1+S处理较N1K1+S处理抑制了根茎对K+的吸收,增加叶片中Na+含量的积累和Na+/K+。N2K2+S处理较N1K2+S处理显著促进根茎叶中Na+含量的积累和Na+/K+。可见,盐胁迫下,过量的氮钾肥造成的二次胁迫不仅无法缓解盐胁迫对面包果的抑制效应,反而促进了部分器官对Na+的吸收,抑制对K+的吸收。研究表明,盐胁迫可导致植株氮磷钾的缺乏[26-29],是抑制植株生长的原因之一[30]。与本研究结果不同,盐胁迫下面包果根茎中的P含量显著高于非盐胁迫,而叶片中的K+含量显著高于非盐胁迫,对各器官中氮含量及面包果的生长影响不显著,这可能与面包果本身的耐盐性有一定的关系[4]。与N2K1+S处理相比,N2K2+S处理对面包果根茎中的氮钾素积累有显著促进作用,N2K1+S处理较N1K1+S处理显著促进叶片中氮素的积累。有研究表明,盐胁迫会抑制Ca2+从根向地上部分的转运量[31]。Ca2+的吸收减少会干扰Na+和K+离子的吸收转运[32]。本研究结果表明,盐胁迫下面包果茎中Ca2+的含量显著高于非盐胁迫,也说明营养元素Ca2+与植株的耐盐性有关,具体的关系有待进一步验证。
Kaya等[33]研究发现,盐胁迫条件下,对马铃薯进行叶面喷施钾肥可以加快植株生长、改善细胞膜渗透性。本研究结果发现,N2K2+S处理较N2K1+S处理显著增加叶片可溶性糖含量,而N2K1+S处理较N1K1+S处理显著降低叶片可溶性糖含量,对叶片脯氨酸含量有一定促进作用。Siddiqui等[34]对油菜的研究也认为,在低氮条件下增施氮肥能够缓解盐胁迫对生长的抑制作用,可能与氮肥诱导渗透保护剂脯氨酸的产生有关。
综上所述,盐胁迫条件下,面包果根茎叶中的Na+含量和Na+/K+均显著高于非盐胁迫,而叶片中的K+含量显著高于非盐胁迫,这可能与面包果本身的耐盐性有一定的关系。氮钾养分对盐胁迫下面包果的生长指标没有表现出明显的可塑性,其中钾素对盐胁迫下的植株生长表现出一定的缓解效应。过量的氮钾肥不仅无法缓解盐胁迫对面包果的抑制效应,反而促进了部分器官对Na+的吸收,抑制对K+的吸收。适量增施氮钾肥可改善盐胁迫下的细胞膜渗透性,缓解盐胁迫的抑制效应。
参考文献
Shabala S, Cuin T A. Potassium transport and plant salt tolerance[J]. Physiologia Plantarum, 2008, 133(4): 651-669.
Yuan Y H, Shu S, Li S H. et al. Effects of exogenous putrescine on chlorophyll fluorescence imaging and heat dissipation capacity in cucumber (Cucumis sativus L.) under salt stress[J]. Journal of Plant Growth Regulation, 2014, 33(4): 798-808.
Wu X X, Zhu Z W, Li X, et al. Effects of cytokinin on photosynthetic gas exchange, chlorophyll fluorescence parameters and antioxidative system in seedlings of eggplant (Solanum melongena L.) under salinity stress[J]. Acta Physiologiae Plantarum, 2012, 34(6): 2105-2114.
苏兰茜, 白亭玉, 鱼 欢, 等. 盐胁迫对2种菠萝蜜属植物幼苗生长及光合荧光特性的影响[J]. 中国农业科学, 2019, 52(12): 2140-2150.
Sayyad-Amin P, Jahansooz M R, Borzouei A, et al. Changes in photosynthetic pigments and chlorophyll-afluorescence attributes of sweet-forage and grain sorghum cultivars under salt stress[J]. Journal of Biological Physics, 2016, 42(4): 601-620.
Munns R. Comparative physiology of salt and water stress[J]. Plant Cell Environment, 2002, 25(2): 239-250.
Parida A K, Das A B. Salt tolerance and salinity effects on plants: a review[J]. Ecotoxicology and Environmental Safety, 2005, 60(3): 324-349.
Gamett T, Conn V, Kaiser B. Root based approaches to improving nitrogen use efficiency in plants[J]. Plant, Cell and Environment, 2009, 32(9): 1272-1283.
武文明, 陈洪俭, 李金才, 等. 氮肥运筹对孕穗期受渍冬小麦旗叶叶绿素荧光与籽粒灌浆特性的影响[J]. 作物学报, 2012, 38(6): 1088-1096.
Shangguan Z P, Shao M A, Dychmans J. Interactions of osmotic adjustment and photosynthesis in winter wheat[J]. Journal of Plant Physiology, 1999, 154(5): 753-758.
Jeschke W D, Nassery H. K+-Na+ selectivity in roots of Triticum, Helianthus and Allium[J]. Physiology Plant, 1981, 52(2): 217-224.
杨 萍, 邱慧珍, 海 龙, 等. 表层土壤调控措施对苹果根系形态及活力的影响[J]. 甘肃农业大学学报, 2014, 49(2): 89-95.
鲍士旦. 土壤农化分析[M]. 北京: 中国农业出版社, 2000.
杨淑萍, 危常州, 梁永超. 盐胁迫对不同基因型海岛棉光合作用及荧光特性的影响[J]. 中国农业科学, 2010, 43(8): 1585-1593.
Vicente O, Boscaiu M, Naranjo M ?, et al. Responses to salt stress in the halophyte Plantago crassifolia (Plantaginaceae) [J]. Journal of Arid Environments, 2004, 58(4): 463-481.
康爱平, 刘 艳, 王 殿, 等. 钾对能源植物杂交狼尾草耐盐性的影响[J]. 生态学报, 2014, 34(20): 5793-5801.
原俊凤, 田长彦, 冯 固, 等. 硝态氮对盐胁迫下囊果碱蓬幼苗根系生长和耐盐性的影响[J]. 植物营养与肥料学报, 2009, 15(4): 953-959.
Shraf M Y, Khtar K A, Sarwar G, et al. Role of the rooting system in salt tolerance potential of different guar accessions[J]. Agronomy for Sustainable Development, 2005, 25(2): 243-249.
Snapp S S, Shennan C. Effects of salinity on root growth and death dynamics of tomato, Lycopersicon esculentum Mill.[J]. New Phytologist, 1992, 121(1): 71-79.
劉爱荣, 张远兵, 钟泽华, 等. 盐胁迫对彩叶草生长和渗透调节物质积累的影响[J]. 草业学报, 2013, 22(2): 211-218.
Song J, Ding X, Feng G, et al. Nutritional and osmotic roles of nitrate in a euhalophyte and a xerophyte in saline conditions[J]. New Phytologist, 2006, 171(2): 357-366.
Gomez I, Pedreno J N, Moral R, et al. Salinity and nitrogen fertilization affecting the macronutrient content and yield of sweet pepper plants[J]. Journal of Plant Nutrition, 1996, 19(2): 353-359.
Zhao K F, Fan H. Halophytes and its adaptation physiology to salinity habitats[M]. Beijing: Science Press, 2005.
Bose J, Shabala L, Pottosin I, et al. Kinetics of xylem loading, membrane potential maintenance, and sensitivity of K+-permeable channels to reactive oxygen species: physiological traits that differentiate salinity tolerance between pea and barley[J]. Plant Cell and Environment, 2014, 37(3): 589-600.
Yeo A R, Flowers T J. Mechanisms of salinity resistance in rice and their roles as physiological criteria in plant breeding[M]. New York: Wiely, 1984: 151-170.
Ravikovitch S, Yoles D. The influence of phosphorous and nitrogen on millet and clover growing in saline soils affected by salinity[J]. Plant and Soil, 1971, 35(1-3): 555-567.
Kaya C, Higgs D, Kirnak H. The effect of high salinity (NaCl) and supplementary phosphorus and potassium on physiology and nutrition development of Spinach Bulg[J]. Journal of Plant Physiology, 2001, 27(3): 47-59.
Levitt J. Responses of plants to environmental stress[M]. New York: Academic Press, 1980.
Hoai N T T, Shim I S. Kabayashi K, et al. Accumulation of some nitrogen compounds in response to salt stress and their relationships with salt tolerance in rice (Oryza sativa L) seedlings[J]. Plant Growth Regulation, 2003, 41(2): 159-164.
Bernstein L, Pearson G A. Influence of exchangeable sodium on the yield and chemical composition of plants: I. green beans, garden beets, clover, and alfalfa[J]. Soil Science, 1956, 82(3): 247-258.
Termaat A, Munns R. Use of concentrated macronutrient solution to separate osmotic from NaCl specific effects on plant growth[J]. Australia Journal of Plant Physiology, 1986, 13 (4): 509-522.
Cramer G R, Epstein E, Lauchli A. Effects of Sodium, potassium and calcium on salt-stress barley Ⅱ. Elemental Analysis[J]. Physiology Plant, 1991, 81(2): 197-202.
Kaya C, Kiranak H, Higgs D. Enhancement of growth and normal growth parameters by foliar application of potassium and phosphorous in tomato cultivars grown at high (NaCl) salinity[J]. Journal of Plant Nutrition, 2001, 24(2): 357-367.
Siddiqui M H, Mohammad F, Khan M N, et al. Nitrogen in relation to photosynthetic capacity and accumulation of osmoprotectant and nutrients in brassica genotypes grown under salt stress[J]. Agricultural Sciences in China, 2010, 9(5): 671-680.
責任编辑:崔丽虹
