芒果叶绿体基因组密码子使用偏好性分析
2021-09-14唐玉娟赵英黄国弟付海天宋恩亮李日旺金刚
唐玉娟 赵英 黄国弟 付海天 宋恩亮 李日旺 金刚


摘 要:本研究利用CodonW、SPSS、Excel软件对芒果叶绿体基因组55条蛋白编码序列的密码子开展了中性绘图、PR2-plot、ENC-plot和最优密码子分析。结果表明,芒果叶绿体基因的有效密码子(ENC)偏好性不强,分布在35.38~56.56;GC3s含量26.29%,说明密码子末位碱基以A/T结尾为主;ENC与GC3极显著相关。相对同义密码子分析,RSCU>1的密码子有31个,其中,以A和T结尾的29个,表明A和T是基因组偏爱的密码子。中性绘图、PR2-plot和ENC-plot分析表明,芒果叶绿体基因组密码子使用偏好受突变和选择等因素共同作用的影响。通过高频率密码子和高表达优越密码子相结合的方法进行分析,筛选出主要以A和T结尾的芒果叶绿体基因组最优密码子19个,分别是GCT、TGT、GAA、TTT、GGT、CAT、ATT、AAA、TTA、CCT、CAA、AGA、CGA、CGT、AGT、TCT、ACT、GTA 和CTT。
关键词:芒果;叶绿体基因组;密码子偏好性;最优密码子
Abstract: Codon usages of 55 chloroplast genes in mango were analyzed to obtain to the results of neutrality plot, PR2-plot ENC-plot and optimal codons by using CodoW, SPSS and SPSS softwares. The results showed that the effective number of codons (ENC) ranged from 35.38 to 56.56, indicating that the condon bias was weak. GC base pair content in the third condon position of chloroplast genomes in mango was 26.29%, indicating that the third base in the codon was terminated by A/T. Significant correlation was found between ENC and GC3.There were 31 codons with relative synonymous codon usage greater than 1, of which 29 codons ending with A and T, indicating that the third codon preferred A and T.According to neutrality plot analysis, PR2-plot and ENC-plot analysis, mutational pressure combined with selection influenced the pattern of synonymous codon usage across genes in mango chloroplast genomes. In addition 19 codons were affirmed as the optimal codons by using analysis of high-frequency codons and the high expression codons, ending with A and T. There were GCT, TGT, GAA, TTT, GGT, CAT, ATT, AAA, TTA, CCT, CAA, AGA, CGA, CGT, AGT, TCT, ACT, GTA and CTT.
Keywords: Mango; chloroplast; codon usage bias; optimal codons
遗传密码又称密码子,编码同种氨基酸的密码子称为同义密码。同义密码子提供了基因表达和蛋白质进化间的联系,维持着生物体遗传密码的功能完整性。密码子的使用频率不同,有的密码子使用频率高于其他同义密码子,形成了密码子偏好性[1]。不同物种密码子偏性不同,但同一种物种或者亲缘关系相近的物种间却使用了相似的偏好性[2]。密码子偏好性的使用在基因表达水平、共翻译蛋白质折叠、mRNA的稳定性等[3],反映了翻译优化中突变偏好与自然选择的平衡[4],因此研究物种密码子偏好性,为探究物种的进化模式与其在系统发育史上的地位提供依据。
叶绿体是植物特有的细胞器,具有高度的保守性及较低的进化率,是研究植物条形码最有潜力的基因序列[5-6],在植物系统进化、物种鉴定等方面广泛应用[7-8]。完整的叶绿体基因组为认识进化和自然选择提供机理性的启示。不同基因组有其特有的密码子使用模式,密码子使用模式的分析有助于理解生物体适应性及其在系统发育进化史的地位[9],有益于基因表达水平的调控与基因的改造[10]。
芒果(Mangifera indicate L.)是著名的热带水果,享有“热带水果之王”之美称[11]。外观秀丽、果香四溢,含有丰富的维生素、蛋白质、胡萝卜素等,营养价值极高,深受大众的喜爱,是南方热区果农增值创收的主要水果之一[12]。目前为止尚未发现芒果密码子使用偏好性相关方面的研究报道,本研究通过对芒果叶绿体基因组密码子组成及相关参数进行分析,首次阐明芒果密码子使用偏好性的特点,确定芒果叶绿体基因组的最优密码子及密码子偏好性发生的因素,为芒果资源保护、性状改良及叶绿体基因组的应用和研究提供参考依据。
1 材料与方法
1.1 基因序列
芒果叶绿体基因组GenBank登录号为CM021858(https://www.ncbi.nlm.nih.gov/),共60个基因序列。为保证结果的准确性[13],剔除重复且长度小于300 bp的编码序列,选择以ATG为起始密码子,以TAA、TAG、TGA为终止密码子的序列,最終选出55条序列进行分析。
1.2 中性绘图分析
统计密码子3个位置上的GC含量,以GC12为纵坐标,GC3为横坐标绘制散点图进行相关性分析。
1.3 相对同义密码子使用度分析
利用CodonW软件对相对同义密码子使用度(relative synonymous codon usage, RSCU)进行分析。RSCU表示一个密码子的实际使用频率与无偏好性时理论使用频率间的比值。RSCU<1时,表示该密码子的使用频率低于其他同义密码子;RSCU>1时,表示该密码子的使用频率高于其他同义密码子;当RSCU=1时表示该密码子没有偏好性[14]。
1.4 ENC-plot绘图分析
以有效密码子ENC(effective number of condon)为纵坐标,GC3s为横坐标,绘制散点图,计算公式:ENC=2+GC3s+29/[GC3s2+(1-GC3s)2]。结合ENC比值频率对差异进行量化分析[15]。
1.5 PR2-plot绘图分析
计算各密码子第3位上的A、T、C、G的含量,以A3/(A3+T3)为纵坐标,以G3/(G3+C3)为横坐标进行PR2的偏倚分析。
1.6 最优密码子分析
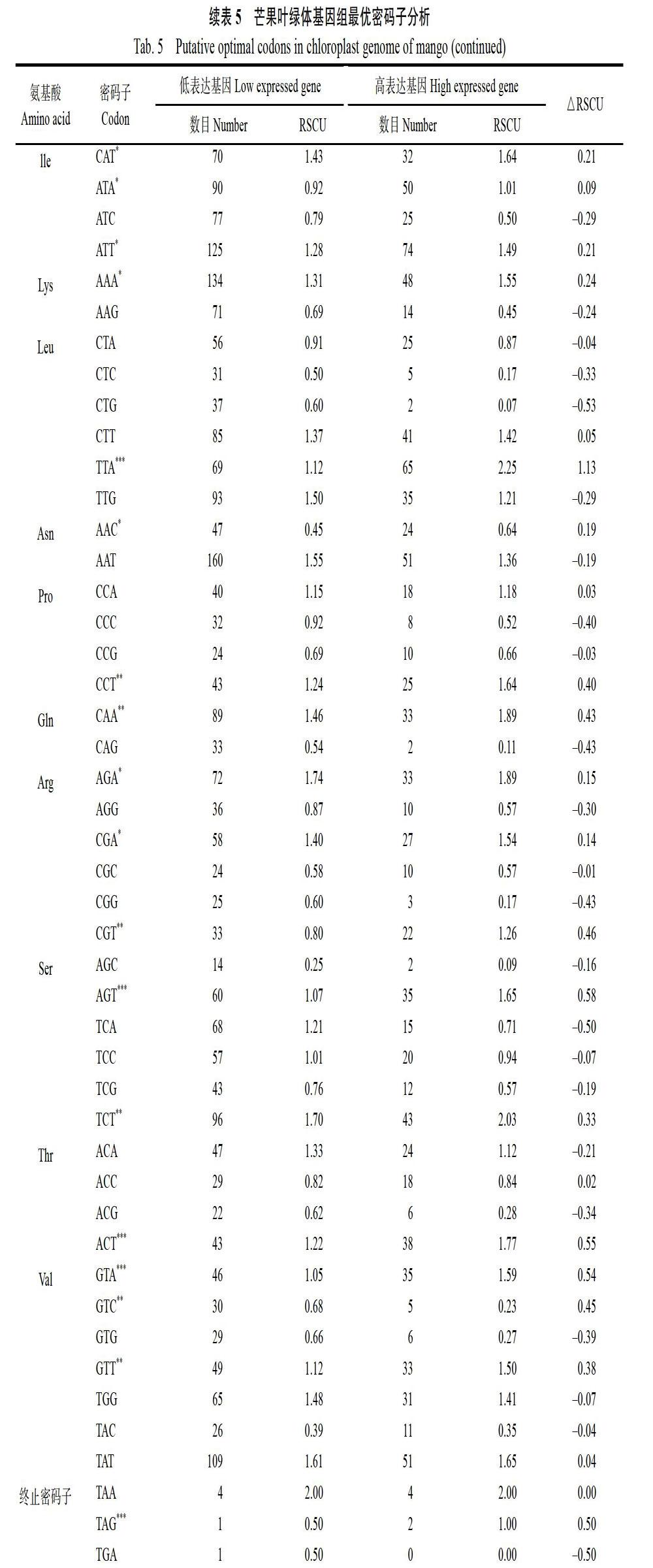
最优密码子以芒果ENC值为偏性参考标准,选择最低和最高10%的基因构建高低偏性库,分别计算两组的RSCU值与△RSCU值,将RSCU>1的密码子确定为高频率密码子,△RSCU>0.08的密码子定义为高表达密码子。同时满足上述2种条件的密码子定义为最优密码子[16]。
2 结果与分析
2.1 密码子使用的总体特征
对55条芒果叶绿体基因组的密码序列的碱基组成进行分析(表1),叶绿体基因组密码子3个位置上的GC平均含量为37.94%。从密码子不同位置的碱基组成来看,GC在密码子的分布不均匀,GC1、GC2、GC3含量分别为47.14%、39.73%和29.35%,第3位上的GC含量低于前两位,分布趋势为GC1>GC2>GC3。GC3s的含量26.29%,表明密码子中的第3位碱基以A/T结尾为主。
ENC表示偏性的强弱,其理论取值范围为21~61,其值越小表示同义密码子的偏性越强,反之同义密码子的偏性越弱。当ENC值为20时,表示同义密码子完全偏倚;当ENC值为60时,表示同义密码子不偏倚[13]。芒果叶绿体基因组编码序列的ENC值均(48.14)值大于40,分布于35.38~56.56之间,由此可以判断芒果叶绿体基因组密码子的偏性不强。
芒果叶绿体基因密码子参数的相关分析显示GCall和GC1、GC2、GC3极显著相关(表2)。GC1和GC2、GC3显著相关,说明芒果叶绿体基因密码子3个位置上的碱基组成相似。ENC值与GC2显著负相关,与GC3极显著相关,表明第2、3位上的碱基组成,特别是第3位对密码子的偏好性影响较大。
2.2 中性绘图分析
芒果叶绿体基因组GC12的取值范围在0.33~0.56之间,GC3的取值范围在0.18~0.32之间,基因全部落在中线对角线上方(图1)。GC12与GC3的相关系数0.279(表2),双尾检测相关性不显著,回归系数(曲线斜率)为0.3544,相关性较弱。结合图1和表2表明芒果叶绿体基因组密码子第1、2位和第3位碱基之间关联性不大,自然选择在芒果叶绿体基因组密码子偏好使用中起主要作用。
2.3 ENC-plot绘图分析
ENC-plot绘图以ENC比值[(预期ENC值~实际ENC值)/预期ENC值]和ENC比值频数判断偏好性影响因素,揭示ENC与GC3s的关系,反映碱基组对密码子偏好性的影响程度[15-19]。基因坐标点离标准曲线的距离近且数量较多说明偏好性主要受到突变的影响;反之说明偏好性受选择的影响较大。图2结合表3显示60%共33个基因分布在–0.05~0.05之间,这33个基因的实际ENC值与预期ENC值接近,与标准曲线的距离较近,;分布在–0.05~0.05之外的基因有22个,这22个基因与预期ENC值较远,在标准曲线较远的位置。说明芒果基因组密码子偏好性与GC3的差异有关,突变对芒果叶绿体基因组密码子偏好性的影响较大。
2.4 PR2-plot绘图分析
PR2-plot绘图通过分析密码子第3位碱基的组成关系,揭示密码子使用偏好性的影响因素。由图3可以看出芒果基因在4个区域内分布不均匀,多数分布于平面图的左下方。T和C在密码子第3位出现的频率较高,G与C、A与T的使用频率不相等,T的使用频率高于A,C的使用频率高于G,嘧啶的使用频率高于嘌呤,说明突变不是影响芒果密码子使用偏好性的唯一因素,选择等其他因素对芒果密码子使用偏好性也产生一定的影响。
2.5 相对同义密码子使用度分析
利用codonW 软件对芒果55条基因序列的RSCU值进行计算(表4)。结果显示RSCU>1的密码子共31个,出现频率相对较高的是以A(13个)或T(16个)结尾的密码子,是芒果基因组偏爱;以G(1个)或C(1个)结尾的密码子出现的频率较低且RSCU值多小于1,说明这些是基因组非偏好的密码子。
2.6 最优密码子分析
芒果叶绿体基因组,以△RSCU≥0.08为标准确定了24个高表达密码子,以A结尾的8个,以T结尾的12个,以C结尾的3个,以G结尾的1个。结合芒果叶绿体基因的相对同义密码子使用度和同义密码子相对使用度(表4和表5),确定19个最优密码子,分别是GCT、TGT、GAA、TTT、GGT、CAT、ATT、AAA、TTA、CCT、CAA、AGA、CGA、CGT、AGT、TCT、ACT、GTA、CTT。其中以A结尾的7个,以T结尾的12个,暗示芒果叶绿体基因组偏爱使用以A和T堿基结尾的密码子。
3 讨论
随着分子生物学的飞速发展,测序技术的持续更新,基因组数据的应用为密码子的偏性研究提供了支持。研究发现GC含量可反映密码子使用偏性突变的整体趋势[20],密码子第3位碱基GC3含量可作为密码子偏好性分析的主要依据[21]。本研究通过对筛选出的芒果叶绿体基因组的55条密码序列进行分析发现,GC3s含量为26.29%,暗示密码子偏好使用末位碱基为A/T的密码子。结果与剑麻[22]、杉木[23]偏好使用末位碱基为A/T的密码子相似。ENC可判断基因表达量的高低,当其大于35时基因表达水平较低,在编码氨基酸时,对密码子的选用偏好性也弱。芒果叶绿体基因组ENC均值为48.14,表明叶绿体基因密码子偏好性不强。ENC越小,基因的密码子使用偏好性越强,越偏好使用A/T结尾的密码子[24],这与本实验RSCU等数据揭示的芒果叶绿体基因组第3位密码子富含A/T的结论相一致。突变和自然选择是影响密码子偏好性使用的主要因素,本研究通过中性绘图分析显示,选择是密码子偏好性的主要影响因素;通过ENC-plot绘图分析则发现,突变是密码子偏好性的主要影响因素,本研究認为芒果叶绿体基因组密码子的偏好性受突变和选择2种因素的影响应该是相对均衡的。
同义密码子的偏好使用影响基因的表达,最优密码子可以提高翻译的效率和准确率[25]。有研究认为,双子叶植物偏好使用A/T结尾的密码子,单子叶植物偏好使用G/C结尾的密码子[26],但也有不同的观点,如有偏好使用A/T结尾的单子叶植物如糜子[27]、霍山石斛[28]等。芒果属于双子叶植物,本研究通过高表达优越密码子和高频密码子相结合的方法筛选出的19个最优密码子均以A或T结尾,表明芒果叶绿体基因组偏爱使用以A和T碱基结尾的密码子,结论与双子叶偏好使用A/T结尾的密码子类似。
本研究系统的对芒果叶绿体基因组密码子的偏好性及影响因素进行了研究,发现芒果叶绿体基因组偏好使用以A/T结尾密码子;突变和选择均衡的影响密码子的偏好性使用;研究还确定了19个芒果叶绿体基因组最优密码子。研究结果为今后芒果分子系统进化、遗传转化、叶绿体基因工程等方面的研究奠定基础,为芒果在叶绿体基因组水平上进行分类、性状改良等提供参考依据。
参考文献
Grantham R, Gautier C, Gouy M. Codon frequencies in 119 individual genes confirm consistent choices of degenerate bases according to genome type[J]. Nucleic Acids Research, 1980, 8(9): 1893-1912.
Grantham R, Gautier C, Gou M. et al. Codon catalog usage and the genome hypothesis[J]. Nucleic Acids Research, 1980, 8(1): 49-62.
文 言. 川桑密码子使用模式与密码子分析工具的开发[D]. 重庆: 西南大学, 2017.
Wu X M, Wu S F, Ren D M, et al. The analysis method and progress in the study of codon bias[J]. Hereditas, 2007, 29(4): 420-426.
Gielly L, Taberlet P. The use of chloroplast DNA to resolve p;ant phylogeny: noncoding versus rbcL sequences[J]. Molecular Biology and Evolution, 1994(11): 769-777.
Shaw J, Lickey E B, Beck J T, et al. The tortoise and the hare:relative utility of 21 noncoding chloroplast DNA sequences for phylogenetic analysis[J]. American Jourmal of Botany, 2005(92): 142-166.
李 汐, 祝 铭, 孙延霞, 等. 基于叶绿体rps16基因和核基因ITS片段研究肉苁蓉属系统位置[J]. 植物科学学报, 2012, 30(5): 431-436.
李丹丹, 郭水良, 于 晶, 等. 基于4个叶绿体基因识别蓑藓属(Macromitrium)植物的可行性研究[J]. 植物科学学报, 2013, 31(1): 23-33.
张月荣. 禾本科叶绿体基因组密码子使用模式及紫茎泽兰叶绿体RNA编辑分析[D]. 杨凌: 西北农林科技大学, 2013.
刘 柱, 胡新文, 刘志昕. 基因及表达调控中遗传密码子选择的偏爱性[J]. 生命科学研究, 2011, 5(3): 106-109.
付海天, 赵 英. 芒果SRAP-PCR反应体系的优化与退火温度的筛选[J]. 福建农业学报, 2014, 29(4): 345-349.
唐莹莹, 黄国弟, 陈永森, 等. 杧果新品种‘桂热杧3号对炭疽病和细菌性黑斑病的抗性评价初报[J]. 中国南方果树, 2020, 49(3): 35-39, 45.
Rosenberg M S, Subramanian S, Kumar S. Patterns of transitional mutation biases within and among mammalian genomes[J]. Molecular Biology and Evolution, 2003, 20(6): 988-993.
冯海悦, 张 羽, 吴正常, 等. 13种哺乳动物MUC4基因密码子使用模式分析[J]. 农业生物技术学报, 2018, 26(9): 1546-1556.
Sueoka N. Near homogeneity of PR2-bias fingerprints in the human genome and their implications in phylogenetic analyses[J]. Journal of Molecular Evolution, 2001, 53(4-5): 469-476.
原晓龙, 李云琴, 张劲峰, 等. 降香黄檀叶绿体基因组密码子偏好性分析[J/OL]. 广西植物, 2021, 41(4): 622-630.
劉 慧, 王梦醒, 岳文杰, 等. 糜子叶绿体基因组密码子使用偏性的分析[J]. 植物科学学报, 2017, 35(3): 362-371.
Sueoka N. Directional mutation pressure and neutral molecular evolution[J]. Proceedings of the National Academy of Sciences of the United States of America, 1988, 85(8): 2653-2657.
李慧姬, 吉雪花, 朱冉冉, 等. 10种植物PSY基因密码子使用偏好性分析, 西北农业学报, 2020,29(2): 276-284.
Zhou M, Long W, Li X. Analysis of synomous codon usage in chloroplast genome of Populus alba[J]. Journal of Forestry Research, 2008, 19(4): 293-297.
冯瑞云, 梅 超, 王慧杰, 等. 籽粒苋叶绿体基因组密码子偏好性分析[J]. 中国草地学报, 2019, 41(4): 8-15.
金 刚, 覃 旭, 龙凌云, 等. 剑麻叶绿体基因组编码序列密码子的使用特征[J]. 福建农林大学学报, 2018, 47(6): 705-710.
郑薇玮, 陈金慧, 郝兆东, 等. 杉木等5种针叶树叶叶绿体密码子偏好性分析[J]. 分子植物育种, 2016, 14(5): 1091-1097.
雷 慧, 李 鸽, 王娜玉. 金莲花叶绿体基因组密码子偏好性分析[J]. 山西农业科学, 2019, 47(8): 1300-1305, 1445.
Sharp P M, Matassi G. Codon usage and genome evolution[J]. Current Opinion in Genetics & Development, 1994, 4(6): 851-860.
Wang L J, Roossinck M J. Comparative analysis of expressed sequences reveals a conserved pattern of optimal codon usage in plants[J]. Plant Molecular Biology, 2006, 61(4-5): 699-710.
刘 慧, 王梦醒, 岳文杰, 等. 糜子叶绿体基因组密码子使用偏性的分析[J]. 植物科学学报, 2017, 3(3): 362-371.
谢析颖, 李 蓉, 林玉玲, 等. 霍山石斛转录因子MYB1R1密码子选用偏好性分析[J]. 分子植物育种, 2017, 15(5): 1683-1690.
责任编辑:白 净
