CIH 所致颏舌肌功能改变中5-HT 调控作用的初步研究
2021-09-10刘雅洁王欢欢张钦鑫蒋冬晖史雅文吴婷殷敏
刘雅洁 王欢欢 张钦鑫 蒋冬晖 史雅文 吴婷 殷敏
阻塞性睡眠呼吸暂停(obstructive sleep apnea,OSA)与睡眠期间上呼吸道塌陷有关,越来越多的研究提示,引起上呼吸道肌群功能改变的因素在OSA的发生发展过程中起重要作用。舌下神经核及舌下神经调控上呼吸道肌,支配伸舌肌如颏舌肌的运动神经元位于腹侧集群,支配缩舌肌如舌骨舌肌的运动神经元分布在背侧。睡眠过程中伸舌肌活性的降低导致上呼吸道倾向于塌陷,引发打鼾与呼吸暂停。另一方面,有研究发现OSA 患者上呼吸道肌肉存在炎性改变和去神经病变,这可能提示OSA 也反过来影响这些肌群的功能[1]。
5-羟色胺(5-hydroxytryptamine,5-HT)又名血清素,是一种生物胺,以其作为神经递质的作用而著称,在调节胃肠运动,外周血管、脑血管紧张度和血小板功能等方面发挥作用,并与情绪障碍,呕吐,偏头痛,肠易激综合征以及肺动脉和系统性高血压的病理生理有关。5-HT 对呼吸功能具有明显的中枢兴奋作用。5-HT 对支配上气道扩张肌的舌下神经运动神经元产生兴奋性刺激。脊髓挫伤所致呼吸功能不全大鼠通过5-HT 系统可恢复正常通气反应[2]。而通过抑制突触前膜对5-HT 的再摄取,突触中5-HT 水平增高,从而减少缺乏单胺氧化酶A 的小鼠的中枢性睡眠呼吸暂停发作[3]。这些研究都提示5-HT 在睡眠呼吸暂停中可能发挥保护性作用[4]。而机体在正常衰老过程中,可用的5-HT 受体数量和5-HT 活性随年龄的增加而减少,这可能是老年人OSA 发生率增加的一个原因[5]。
另一方面,钙稳态失衡与肌肉疲劳的增加密切相关。钙稳态的维持依赖于肌膜、肌浆网和线粒体中存在的复杂而完整的钙转运系统,而肌浆网是转运系统的中心。肌浆网是肌肉中主要的细胞器,负责调节游离钙,在肌肉收缩和舒张中起核心作用。肌浆网钙ATP 酶(sarco-endoplasmic reticulum Ca2+-transporting ATPase,SERCA)和兰尼定受体(ryanodine receptor,RYR)都是Ca2+处理蛋白的成员。骨骼肌中T 膜的去极化产生的膜电位的变化可以诱导二氢吡啶受体的构象变化最终引起肌浆网膜上RYR 的活化。RYR 的活化导致钙离子大量的释放,钙离子通过与肌钙蛋白结合引起肌肉的收缩。SERCA 将游离的钙泵入肌浆网,从而促进肌肉松弛。有研究提示,SERCA 功能的改变可能与骨骼肌疲劳有关[6]。而缺氧可能会损害SERCA 活性,加剧疲劳的发展。
基于既往报道,我们推测在慢性间歇性低氧(chronic intermittent hypoxia,CIH)背景下颏舌肌的功能改变是OSA 病理生理机制的重要环节,而在这一环节中损伤和代偿机制可能并存。我们的前期工作关注了5-HT 对舌下神经核的调节作用。因此本研究我们关注舌下神经支配的颏舌肌功能变化,以及5-HT 对其的调控作用,现报道如下。
资料与方法
1 动物模型
20只清洁级健康成年雄性sprague dawley(SD)大鼠(8 周龄,体重180~200g),由江苏省动物实验中心提供,随机均分为四组,分别为常氧组(control组),以及3 周、5 周和8 周CIH 组。CIH 组大鼠每d 8∶30~16∶30 放置于低氧舱(中国新飞仪器公司)中饲养,循环交替给予60s 氮气和60s 压缩空气,每一循环120s。同时用测氧仪检测舱内氧体积分数变化,使舱内氧体积分数在5%~21%之间循环。对照组在常压、大气环境培养舱中饲养,舱内氧体积分数维持在21%。每日CIH 造模结束后将动物送入常规饲养箱。两组动物进食、饮水、活动均不受限,饲养条件相同。造模分别持续21d、35d、56d。本实验方案由南京医科大学实验动物伦理委员会批准。
2 取材
2.1HE 染色与免疫组化取材
造模结束后,四组动物在规定时间点以2%戊巴比妥钠(4 mg/100 g)麻醉后固定于手术木板上,置于解剖盘中,暴露胸腔并游离出心脏,灌流针经左心室插入升主动脉,剪开右心房,先用生理盐水快速灌注5 min(60 ml)左右以移除血液,直到肝脏发白。然后用4%多聚甲醛400~500 ml 灌注固定,直到动物的肝脏发硬,尾巴僵直,完成灌注。取动物颏舌肌,保存在4%多聚甲醛中4℃冰箱储存,用于后续实验。
2.2Western blot 取材
以2%戊巴比妥钠(4mg/100g)麻醉后立即断头,取颏舌肌后,立即置入冻存管中,存放于-80℃冰箱中。
3 实验过程
3.1HE 染色
肌肉组织经脱水透明后PBS 冲洗,石蜡包埋后切片,横截面设为4 μm。经过展片与贴片放于60℃烘箱2h 避免脱片,随后将切片置于玻片架上后开始脱蜡。经苏木素染液7 min,自来水洗2 min,1%盐酸酒精2~5s 分化,自来水洗5~7 min 返蓝,1%伊红染液2~4 min,自来水洗1 min。再次将切片按浓度梯度放入70%、80%、90%、95%乙醇1min,无水乙醇I、无水乙醇II 各1min,二甲苯I、二甲苯II 各2min,待切片风干后中性树胶封片,最后在显微镜下观察。
3.2免疫组织化学
脱蜡前步骤同HE 染色,随后切片于95℃高压锅进行抗原修复,PBS 冲洗后用免疫组化笔在组织周围画圈,封闭抗原,随后在圈内滴加稀释好的(1∶100)一抗,置于4℃湿盒中孵育过夜。放置复温,PBS冲洗后滴加稀释后的二抗,室温或37℃孵育20~30min。PBS 再次冲洗切片,吸水纸擦干,滴加新鲜配制的DAB 显色液。显微镜下观察到阳性信号(棕黄色或棕褐色)后水洗终止显色。苏木素复染后再次脱水透明,风干后中性树胶封片,显微镜下观察和采集图像。图像采用IPP 图像分析系统分析其平均光密度值(平均光密度值=累积光密度/面积)。
3.3Western Blot
采用GAPDH(glyceraldehyde-3-phosphate dehydrogenase,即甘油醛-3-磷酸脱氢酶,该酶参与糖酵解反应,几乎在所有组织中都高水平表达,且表达量较为恒定,故被广泛用作实验操作的标准化内参)作为内参通过蛋白质印迹法(Western-blot)测定5-HT 2A 蛋白的表达量。向组织中加入配制好的裂解液,机械匀浆后冰上静置30min,全程冰上操作。4℃,12000 转离心15 min,取上清即可得蛋白样品,采用CBA 法测量蛋白样品的浓度。以1∶4 的比例向蛋白样中加入蛋白上样缓冲液,煮蛋白。制备10%SDS-PAGE 凝胶,加样后开始电泳,待蛋白跑至凝胶下方时将SDS-PAGE 凝胶转移到PVDF 膜上。转膜结束后以5%脱脂牛奶震荡封闭2 h。将条带放入稀释好的抗5-HT 2A(1∶500,bs1056R)溶液中,4℃摇床孵育过夜。TBST 洗涤后放入稀释后的二抗(HRP标记山羊抗兔抗体)溶液中,震荡孵育1.5 h。取出条带,再次TBST 洗涤。避光条件下配制ECL 显影液,混匀后,均匀滴在PVDF 膜上,使用化学发光凝胶成像系统进行曝光。
4 统计学处理
采用SPSS 25 统计软件进行统计分析,数据符合正态分布,结果均以均数±标准差()表示,任意两组间比较采用独立样本t-test,趋势检验采用线性回归分析。P<0.05 为差异有统计学意义。
结果
1 颏舌肌HE 染色(图1)
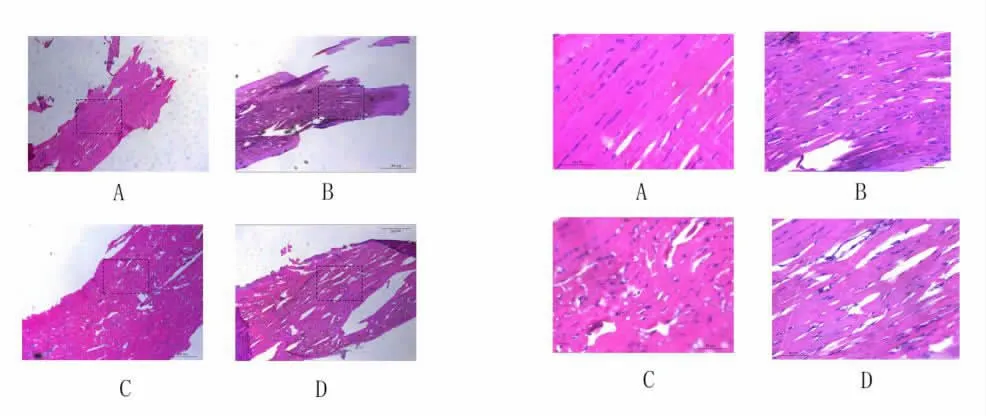
图1 颏舌肌HE 染色
与常氧组相比,CIH 组肌纤维及细胞核排列杂乱,肌纤维分散变细,肌纤维间隙增大。同时,CIH 时间越长,上述变化越明显。
2 5-HT 在颏舌肌的表达(图2)
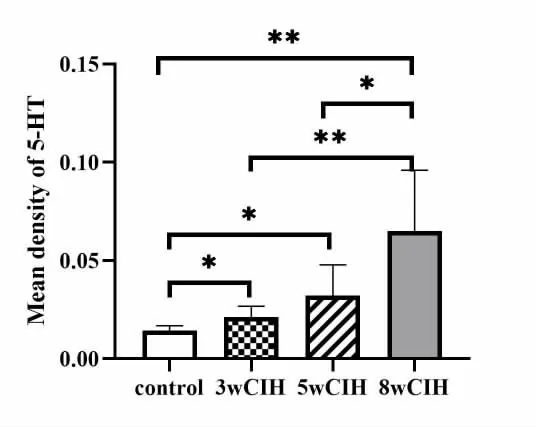
图2 颏舌肌5-HT 免疫组化结果
与常氧组相比,CIH 各组5-HT 表达量明显增高并呈上升趋势,并在CIH-8 周组达到高峰。
3 5-HT 2A 受体的表达(图3)
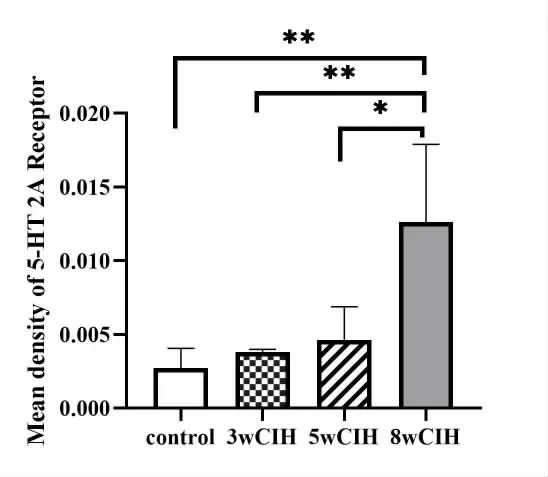
图3 颏舌肌5-HT 2A 受体免疫组化结果
5-HT 2A 受体的表达量在3 周与5 周时升高并不明显,但在8 周达高峰(P<0.01),总体亦呈上升趋势。
4 RYR 免疫组化结果比较(图4)
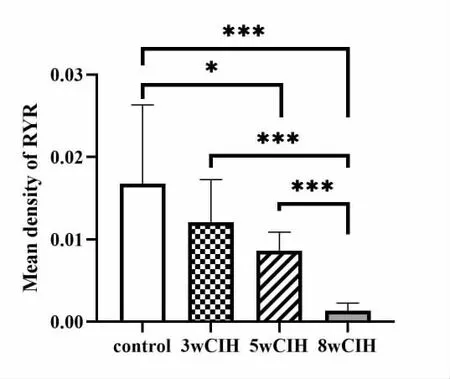
图4 颏舌肌RYR 免疫组化结果
RYR 表达量呈下降趋势,在5 周(P<0.05)、8 周(P<0.001)时明显下降,其中8 周与3 周、5 周组比较差异均具有统计学意义(P<0.001)。
5 SERCA 免疫组化结果比较(图5)
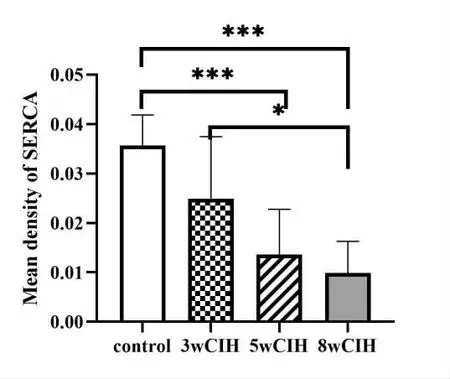
图5 颏舌肌SERCA 免疫组化结果
SERCA 受体表达量与常氧组比较,在5 周、8周时均明显下降(P<0.001)。
6 5-HT 2A 受体的蛋白表达量(图6)
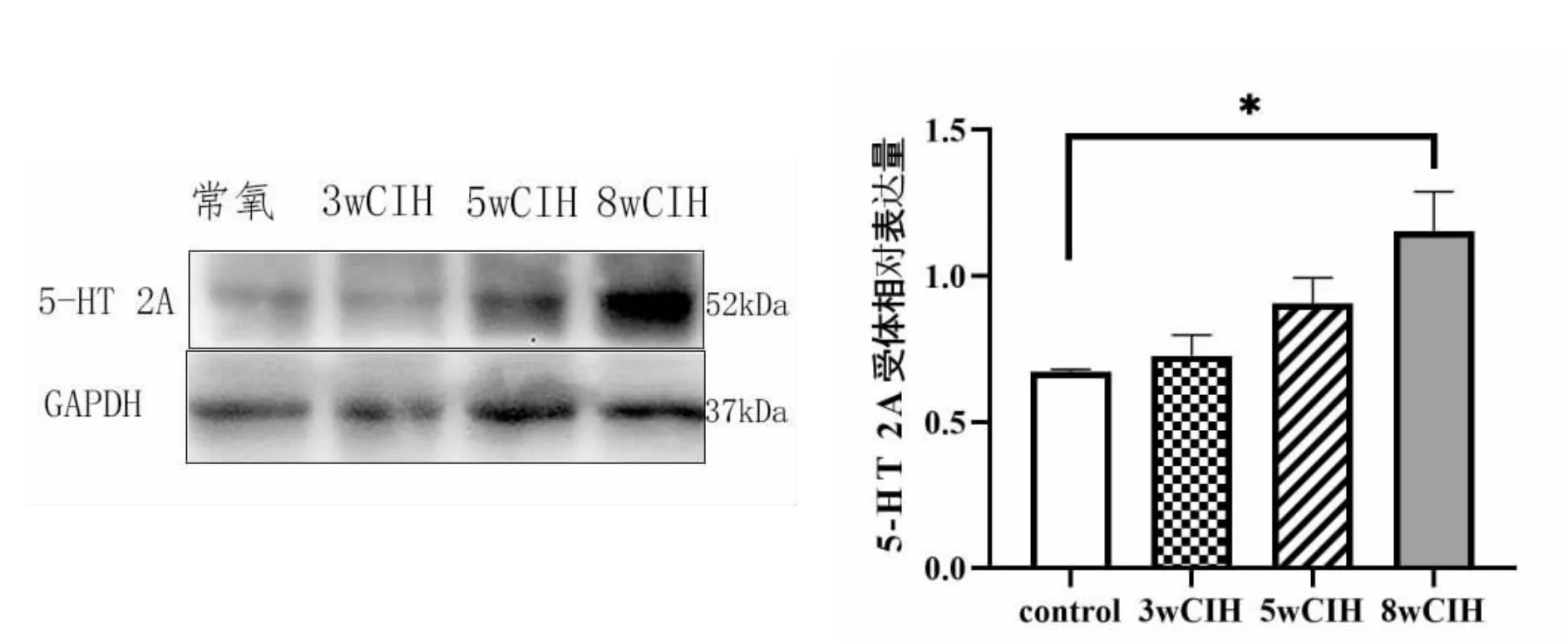
图6 颏舌肌5-HT 2A 受体蛋白表达量
5-HT 2A 受体表达量逐渐增高,与常氧组比较在8 周CIH 时明显升高(P<0.05)。
讨论
我们的研究结果显示,CIH 组肌纤维及细胞核排列杂乱,且随CIH 程度加重变化越明显。5-HT 及其2A 受体在颏舌肌的表达水平随着CIH 时长呈增高趋势,并在8 周时达到高峰。然而钙离子通道调控蛋白SERCA 与RYR 的表达水平则随CIH 时长的增加逐步下降。
一方面,CIH 是造成肌肉损伤的直接原因。有研究表明,慢性缺氧改变了大鼠的呼吸肌肌力和疲劳性,这要么是来自缺氧的直接影响,要么是来自缺氧引起呼吸驱动增加所带来的适应性反应的活化增加[7]。另一方面,在CIH 导致的肌肉损伤反过来可能加重上气道的塌陷。Meng 等[8]通过对临界压力(Pcrit)的动态监测来评估上气道的稳定性,发现随着CIH 的进展,上气道呈现出更高的易坍塌性。
OSA 或CIH 导致肌肉疲劳的潜在机制尚不清楚。一些研究表明,CIH 导致上呼吸道肌肉纤维从缓慢型转变为快速型。CIH 可能会引发肌肉纤维组成的变化,这可能是疲劳的特征,但尚存在争议。Pae等[9]发现,10h 的间歇性缺氧即可导致颏舌骨肌纤维类型由肌球蛋白重链(MHC)2A 转变为MHC 2B。CIH 改变了颏舌骨肌的组成和物理特性,使其更易疲劳,这可能是OSA 上呼吸道肌群容易疲劳的肌纤维类型的变化原因。另一个上气道扩张肌肌肉损伤的机制可能是通过氧化应激的作用实现的。Shortt等[10]发现间歇性缺氧下大鼠膈肌的氧化应激导致呼吸肌的耐力受损。第三,疲劳还被认为是肌浆网钙释放逐渐减少的结果,其中RYR 与SERCA 在骨骼肌功能的调节中起着关键作用。在肌纤维中,Ca2+主要储存在肌浆网末端池内,通过RYR 介导的Ca2+释放通道释放,引起肌纤维收缩;Ca2+通过SERCA 回收到肌浆网内,使肌纤维舒张。若缺乏适当的细胞内游离钙信号来激活收缩装置则会导致疲劳。SERCA 活性的下降会导致钙再摄取的减少,这可使内皮细胞Ca2+浓度升高,进而引起肌间线蛋白的降解,激活蛋白酶和磷脂酶破坏肌肉结构,导致收缩功能下降;另一方面,由于钙的再摄取量和速度降低,导致疲劳性增加,从而减缓肌凝蛋白与肌动蛋白形成“桥”的解离速度,对肌肉舒张也产生影响。
在肌浆网钙释放减少引起疲劳这一方面,我们的结果与在4 周CIH 大鼠心肌组织中RYR 及SERCA 表达水平相似[11],也发现其表达水平随CIH时长的增加后逐步下降。慢性间歇性缺氧类似于心肌出现的缺血再灌注,但骨骼肌与心肌组织的收缩舒张机制对Ca2+的依赖性却不尽相同。一项对骨骼肌缺血再灌注模型的研究中也发现了RYR 的表达下调,但在早期却表现出RYR 水平的不变化或升高。这一现象可能与钙超载及超早期细胞膜内外电荷水平变化有关,随着后期细胞器等的损伤,RYR水平便随之下降。另有研究也发现了颏舌肌SERCA水平的下降,且这可能与CIH 的程度及时程有关[12]。前文中CIH 研究环境为5 周,每周5d,而我们的研究环境分别为3 周、5 周、8 周,每周7d,增加了对SERCA 变化的动态观察,对Ca2+在CIH 的病理机制中起的作用有了更直观的了解。
5-HT 与CIH 引起的呼吸中枢损伤密切相关。中缝核是分泌5-HT 的主要核团,脑干呼吸神经元支配舌下运动神经核,继而将呼吸驱动信号传递至舌部肌群[13],由此,中缝核中5-HT 的降低可能促进OSA 的发生。另外,5-HT 还参与了CIH 大鼠颈动脉窦神经放电的长时程易化作用。在大鼠前包钦格复合体也发现了5-HT 及5-HT 2A 参与对缺氧诱导的呼吸神经可塑性调控的证据[14]。我们在既往研究中[15]也发现了CIH 大鼠舌下神经核腹侧核及背侧核5-HT 及5-HT 2A 受体表达水平的明显上调。同时,5-HT 在肌电活动变化中的也起了相应作用。5-HT通过兴奋舌下运动神经核使得颏舌肌活性增加的作用与睡眠觉醒状态相关[16],从觉醒到睡眠的过程中5-HT 的作用随之减弱,使舌下神经向颏舌肌发放电活动减少,最终导致颏舌肌活性下降,引起上气道阻塞。
目前关于上呼吸道肌群中5-HT 的研究较少。给麻醉大鼠中枢注射5-HT 及其受体亚型,通过上气道肌电活性发现了5-HT 的维持上气道肌群稳定和预防咽部塌陷作用[17]。我们的研究直观地在颏舌肌上发现了5-HT 及其受体水平的增高。并且,颏舌肌5-HT 在3 周、5 周、8 周CIH 时都有明显升高,但5-HT 2A 受体表达的升高仅但8 周CIH 时才具有统计学意义,我们考虑增高的5-HT 是由其上游,即舌下神经核分泌而来,而5-HT 2A 受体则由肌肉本身受刺激产生,因此具有一定的延后性。
总的来说,本研究提示慢性间歇性缺氧早期即出现颏舌肌的损伤,表现为形态学和相关钙通道调控蛋白水平的下降,但在一定程度上也存在相应的代偿性保护,表现为5-HT 及5-HT 2A 受体水平的升高。在一定阶段内,5-HT 的代偿保护机制起着主导作用,舌下神经核传输更多的5-HT 至下游颏舌肌,通过产生更多兴奋性5-HT 2A 受体,使得肌细胞兴奋性增高、活性增强,从而维持上气道的不易塌陷,以代偿性适应上气道的解剖性狭窄。而这为我们今后的研究提供了方向,即何种程度CIH 造成的损伤可被完全代偿,进而推导至整体,何种程度OSA 引起的损伤是机体失代偿的开始,这些探索将对OSA 疾病的认识具有深远意义。
