食用菌多糖结构相对有序性研究概述
2021-09-09聂少平王玉箫殷军艺
聂少平,王玉箫,殷军艺
(南昌大学 食品科学与技术国家重点实验室 南昌 330047)
自上世纪80年代日本科学家Chihara 等[1]首次发现香菇多糖具有显著抑制肿瘤生长的作用起,食用菌的化学成分及生理活性开始备受关注。食用菌不仅味道鲜美、热量低,且富含多糖、多酚、蛋白质、维生素、矿物质、三萜类及甾醇类等营养成分[2],是人们日常膳食的重要组成部分。食用菌细胞壁中80%的干重成分为多糖,以微纤维状排列于由糖蛋白、脂质和非纤维多糖构成的基质中。食用菌多糖因来源广泛、品种丰富且具有多种生物活性等特点而成为人类研究和开发的热点之一[3]。现代药理学研究表明,食用菌多糖具有广泛的生物学活性[4],有望从中发现疗效显著、安全性高的多糖类药物,如香菇多糖(Lentinan)可用于辅助治疗恶性肿瘤及肝炎等疾病[5],灵芝多糖(Ganoderma lucidum polysaccharide)可以改善Ⅱ型糖尿病大鼠的肠道菌群,进而调节其血糖血脂水平[6],羊肚菌多糖(Morchella crassipes polysaccharide)可以缓解人肺泡上皮细胞的氧化损伤等[7]。
食用菌多糖的生理活性与其化学结构息息相关。近年来,各国学者逐渐意识到:明确食用菌多糖的精细结构是将研究的关键成果转化为免疫调节剂等功能性产品的重要前提。然而,其结构的复杂性和多样性,导致对食用菌多糖结构的了解仍有很大的提升空间[8]。以目前研究最广泛的香菇多糖为例,其结构解析结果大致可以分为葡聚糖类和非葡聚糖类。1976年,日本学者Sasaki 等[9]首次发现香菇多糖的主链结构由β-D-(1→3)-Glcp 构成,在O-6 位上有分支,侧链结构由β-D-(1→3)-Glcp 和β-D-(1→6)-Glcp 相连而成(如图1a)。Wang 等[10-11]和You 等[12]分别采用水提醇沉法和1% NaOH/0.05% NaBH4混合溶液提取法从香菇中分离得到β-D-(1→3)-葡聚糖,然而,两者侧链的分支度不同。Zhang 等[13]通过5% NaOH/0.05%NaBH4混合溶液提取的香菇多糖,其主链结构由α-D-(1→3)-Glcp 构成,还含有少量α-D-(1→6)-Glcp 侧链。一些其它常见地香菇β-葡聚糖的重复单元如图1b~1g所示,其主链主要含有β-D-(1→3)-Glcp 和β-D-(1→3,6)-Glcp 残基,支链可能由β-D-(1→6)-Glcp 和/或β-D-Glcp 组成。然而,也有其它文献报道香菇多糖为非葡聚糖结构,例如:半乳聚糖[14]、杂半乳聚糖[15]、甘露半乳葡聚糖[16]和杂聚糖[17]等。同样情况的也存在于灵芝多糖中。继Ukai 等[18]在1982年首次从紫灵芝中发现一种碱溶性的β-D-(1→3)-葡聚糖,高分子的水溶性β-D-(1→3)-葡聚糖在赤灵芝中也被发现,如图2a所示。Zhang 等[19]采用0.25 mol/L LiCl/DMSO 溶液从黑灵芝中分离得到一种线性α-D-(1→3)-葡聚糖。此外,一些非葡聚糖类,如甘露葡聚糖[20]、岩藻半乳聚糖[21](图2b)、半乳葡聚糖[22]和杂聚糖[23]等也在不同种类的灵芝中有所发现。这种多糖结构的异质性往往是由其单糖和糖苷键的多样性造成的,然而,从化学结构的角度来看,不同种类的食用菌多糖也具有一定的“规律性”,如大多数香菇多糖和灵芝多糖都属于β-D-(1→3)-葡聚糖结构,且它们的半乳聚糖和杂半乳聚糖的主链结构均为α-D-(1→6)-半乳聚糖等。
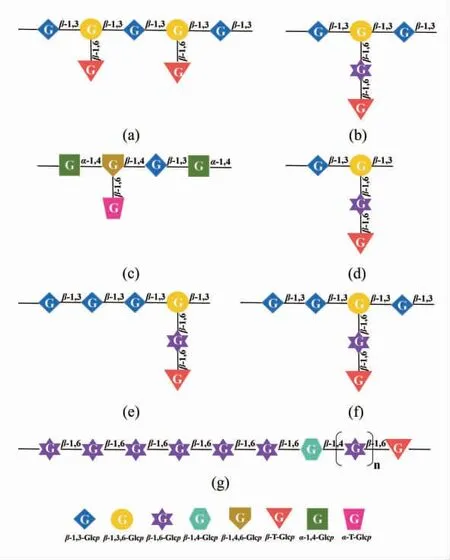
图1 从香菇中分离出的不同β-葡聚糖重复单元结构Fig.1 The different repeating units of β-glucan isolated from Lentinula edodes

图2 从灵芝中分离出的β-(1→3)-葡聚糖(a)和岩藻半乳聚糖(b)的重复单元结构Fig.2 The repeating units of β-(1→3)-glucan (a)and fucogalactan (b) isolated from Ganoderma lucidum
目前对于食用菌多糖一级结构的准确表征仍有提升空间,其原因包括:1)食用菌提取部位不同:食用菌多糖广泛存在于菌丝体、菌核(菌索)、子实体和发酵液中[24],从不同部位或整株提取的食用菌多糖,结构上具有明显差异;2)提取、分离纯化方法不同:除了传统的水提醇沉法[25]和酸/碱提法[26-27]外,超声提取[28]、微波提取[29]、复合酶提取[30]等方法也被广泛应用于食用菌多糖的提取;常用的纯化方法有分级醇沉法[31]、色谱柱法[32]、膜分离法[33]及高速逆流色谱法[34]等;这些因素决定了食用菌细胞壁的提取层次和效率;3)结构分析手段的局限性:目前食用菌多糖定性、定量分析等相关化学研究方法和技术体系还不够完善,仍比较依赖传统方法,缺乏快速精准的分析手段,限制了人们对食用菌多糖结构类型的认识;4)食用菌多糖纯化不到位:以牺牲得率为前提纯化得到的多糖,往往导致其中的有效组分被丢弃,或结构相似的几个组分被当作一个“均一多糖”进行后续的结构解析[35]。这些日益凸显的问题成为食用菌多糖结构鉴定中亟待解决的难题。
笔者在前人大量研究和团队工作的基础上,发现不同多糖分子结构特征(如分支、单个单糖之间的连接和构型)出现的频率和规律是认识天然产物多糖结构的关键,因此提出了“多糖结构的相对有序性”这一理论[36-38],即在某种程度上,多糖的结构有一定规律可循,绝大部分多糖可被归属到某一特征结构类型的化合物,如葡聚糖、葡甘露聚糖、阿拉伯木聚糖、阿拉伯半乳聚糖、果胶等。虽然已有关于食用菌提取、分离纯化、化学组成及活性的综述[39-41],但是多侧重某一种食用菌多糖的结构和功能的阐述,而关于食用菌多糖一级结构的系统性总结归纳鲜有报道。在前期,本团队已成功地将“多糖结构的相对有序性”这一理论应用于海鲜菇多糖[38]和茶树菇[42]多糖的分离纯化。本文采用文献计量学的方法,归纳近10年涉及食用菌多糖化学结构的文献,通过食用菌及其单糖组成、糖苷键及多糖类型之间关系的比较研究,挖掘食用菌多糖的结构规律,以期为食用菌乃至其它天然产物来源多糖的结构研究提供建设性地启示。
1 基于文献计量学的食用菌多糖结构研究进展
1.1 文献来源及关键词
经过文献的去重、数据的清洗以及会议摘要、讲话稿、报道等非相关文献的删除后,检索结果共有17 457 篇相关文献,全部导入COOC/VOSviewer 文献处理软件进行分析,将结果自定义保存为含题名、关键词和摘要的.txt 格式文件。文献下载后,对每篇论文的关键词进行选取、增删、规范、合并,筛选词频大于100 次的关键词做共现分析,可以发现,出现频次大于100 次的关键词共有53个。提取前30 个出现频次大于100 次的关键词,如图3所示,可以发现,目前研究人员对食用菌多糖结构方面的研究较多,且集中在纯化、得率、表观形貌及核磁共振表征技术等方面。功能活性方面,目前研究人员比较关注食用菌多糖的抗氧化、抗肿瘤及细胞毒性等方面的活性研究。另外,目前研究较多的食用菌种类包括香菇、平菇和杏鲍菇等人们日常食用的品种,它们之中大部分属于担子菌亚门 (Basidiomycotina)、层菌纲(Hymenomycetes)、伞菌目(Agaricales)。
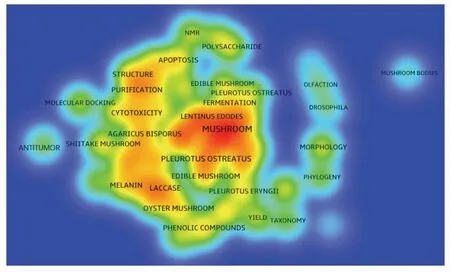
图3 筛选词频大于100 次的前30 个关键词聚类分析Fig.3 Clustering analysis of top 30 keywords screening by word frequency more than 100
1.2 研究热点与发展趋势
在众多关键词中,通过聚类分析可以把关联密切的关键词聚集在一起形成不同的类,以此表示该领域分支的组成情况。利用COOC/VOSviewer统计软件对53 个高频关键词的词篇矩阵进行系统聚类分析,得到相关关系聚类分析图(图4)。其中,横坐标代表53个高频关键词,带有数字的标尺表示分类对象之间的距离。如果2 个关键词聚集到一起的距离越短,说明它们之间的关系越紧密。随着被分类的关键词之间距离变大,最终所有的关键词都将成为一类。图中同种颜色的线指向的关键词具有相关关系,例如:“structure”、“NMR”、“polysaccharide”和“antitumor”同时出现,表明核磁共振技术为食用菌多糖结构解析的常用手段,且食用菌多糖的结构特征与抗肿瘤活性息息相关。由此可以总结出当前食用菌多糖3 个研究热点,分别为:构效关系、资源开发的最大化及菌菇废物的再利用,而这3 个研究热点都需要以食用菌多糖已知结构作为基础进行后续研究。
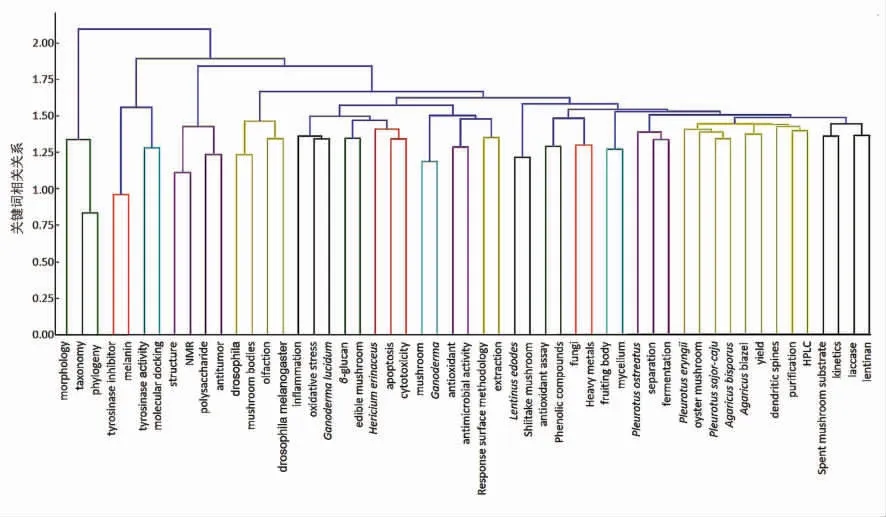
图4 关键词相关关系聚类分析Fig.4 Clustering analysis based on the correlation relationships of keywords
图5展示2011年至2021年食用菌多糖结构类型的风向玫瑰图,可以看出,食用菌多糖的种类很多,类型分布各异。文献报道最多的食用菌多糖结构类型为“β-(1→3)-葡聚糖”,这与其分布的广泛性和较好的活性密切相关。排在第2 位的食用菌多糖结构类型为杂聚糖类(heteropolysaccharide),说明在食用菌多糖的结构和构效关系研究中,纯化不到位的现象普遍存在,导致无法得到均一性较好的多糖。排在第3 位的食用菌多糖结构类型为“β-(1→6)-葡聚糖”,表明目前人们对不同类型的“β-葡聚糖”普遍关注度较高,可能是由于其结构简单、来源丰富且生物学功能较好。此外,在食用菌多糖结构相关的文献报道中,往往存在对同一来源的同种多糖结构定义不同的现象。出现这种情况的原因,除了由于原料来源、生长环境、多糖制备方法等差异外,纯化不完全也会导致多糖均一性不符合结构解析标准,从而得到不准确的多糖结构分析结果[43]。

图5 2011年至2021年食用菌多糖结构类型的风向玫瑰图Fig.5 Wind direction rose map of polysaccharide structure type of edible fungi in 2011—2021
1.3 食用菌多糖的单糖组成
使用Web of Science 检索食用菌多糖结构相关的文献。同样地,选择Web of Science 中的“核心集合” 数据库,以 “mushroom” 或 “fungi”和“polysaccharide”为主题词,设定文献时间为“2011年至2021年”,在检索结果中将主题精炼为“structure”,并将文献类型精炼为“article”或“review”。检索规则如下:
TS=(mushroom)OR TS=(fungi)AND TS=(polysaccharide);时间跨度=2011—2021;精炼依据:主题:(structure);语种:(English);文献类型:(article OR review)。
检索结果显示,从2011年至2021年间,食用菌多糖结构特征相关的文献共有16 108 篇,导入COOC/VOSviewer 软件进行分析,经过文献的去重及数据的清洗、频次统计及可视化分析后,基于共现矩阵绘制所有样本的单糖组成聚类分析。从图6可以发现,存在于食用菌多糖的主要单糖类型共12 种,分别为葡萄糖(Glucose)、半乳糖(Galactose)、甲基-半乳糖(3-O-methylgalactose)、甘露糖(Mannose)、岩藻糖(Fucose)、阿拉伯糖(Arabinose)、木糖(Xylose)、鼠李糖(Rhamnose)、果糖(Fructose)、核糖(Ribose)、葡萄糖醛酸(Glucuronic acid)和半乳糖醛酸(Galacturonic acid)。同种颜色表明这些单糖往往同时出现于食用菌多糖中,比如甘露糖、半乳糖和3-O-Me-半乳糖往往存在于同一种食用菌多糖中。右图列出出现频次最高的9 种单糖,颜色越亮,其出现频次越高,可以发现葡萄糖、半乳糖和甘露糖在食用菌多糖中出现频次最高,其次是阿拉伯糖、木糖、鼠李糖和果糖,最后为葡萄糖醛酸和半乳糖醛酸,表明在食用菌多糖中可能存在少量的酸性糖,且葡萄糖醛酸存在的概率大于半乳糖醛酸。
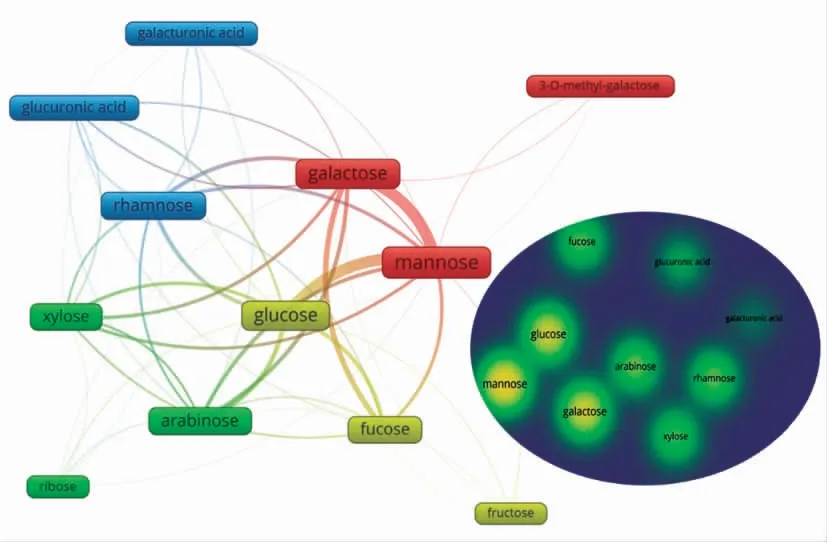
图6 基于共现矩阵的食用菌多糖单糖组成的聚类分析Fig.6 Clustering analysis of monosaccharide composition of fungi polysaccharide based on co-occurrence matrix
1.4 食用菌多糖的结构类型
图7为基于共现矩阵的食用菌多糖结构类型的聚类分析。可以发现,食用菌中出现频次最高的多糖结构类型为β-(1→3)-葡聚糖,约占总数的17.13%,而排在第2 位的是占比为14.81%的heteropolysaccharide。除heteropolysaccharide 外,结构类型的前5 名均为葡聚糖类和杂葡聚糖类,说明近10年研究人员对食用菌中的葡聚糖类和杂葡聚糖的关注较多,可能由于其结构较为简单且来源广泛,易于获得。根据文献报道,香菇、灵芝、冬虫夏草等食用菌中均广泛存在活性较好的β-葡聚糖、α-葡聚糖及杂葡聚糖,具有较高的研究价值[44-45]。此外,对于结构较为复杂的食用菌多糖结构类型,如葡半乳甘露聚糖、岩藻甘露半乳聚糖和杂半乳聚糖等研究较少,虽然已有许多报道证明,食用菌中的杂半乳聚糖和杂甘露聚糖具有很好的功能活性[46-47],但是复杂的纯化过程和化学结构使其分子作用机制的阐明和功能活性的挖掘面临巨大挑战。
后来,由于两人都在美国工作生活,接触频繁起来。胡适为吴健雄做过两件事。一件是胡适在旅行中,在书店淘到一套英国大物理学家卢瑟福的作品,给吴健雄寄去。二是胡适曾给她写过一封长信,内容堪称经典。其实这也没有啥,不过是一位老师对自己钟爱的学生表达激赏与期许吧。胡适曾在写给吴健雄的信里说:我一生到处撒花种子,绝大多数都撒在石头上了,其中有一粒撒在膏腴的土地里,长出了一个吴健雄,我也可以万分欣慰了……
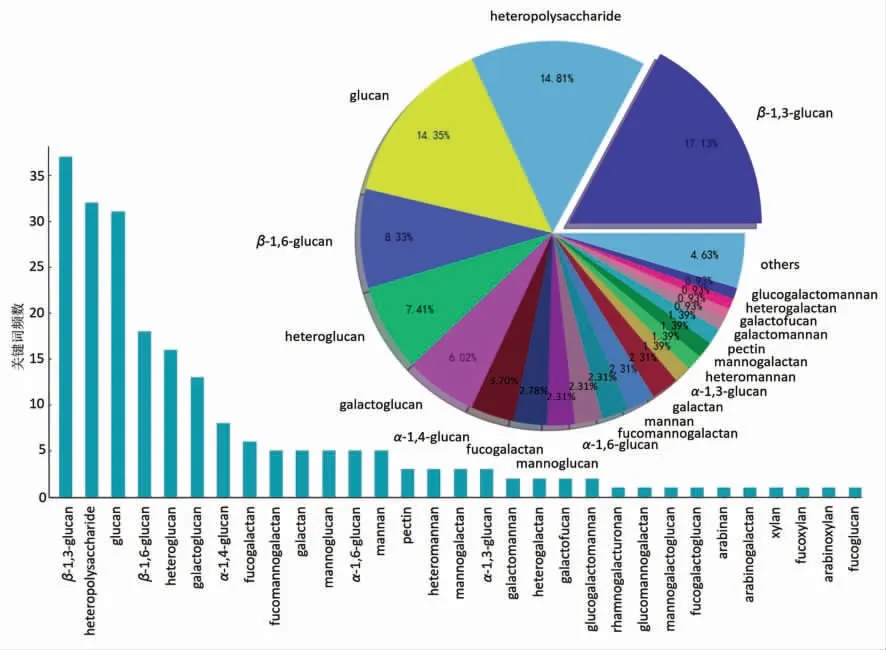
图7 基于共现矩阵的食用菌多糖结构类型的聚类分析Fig.7 Clustering analysis of structure type of fungi polysaccharide based on co-occurrence matrix
1.5 食用菌多糖结构特征的相关性
图8为食用菌及其单糖组成基于二模矩阵的聚类分析。对于香菇多糖来说,葡萄糖出现的频次最高,其次为甘露糖、半乳糖、阿拉伯糖和木糖。在平菇、猴头菇、杏鲍菇、灵芝等市面上常见的食用菌中,葡萄糖出现频次仍然最高。此外,双孢蘑菇和桑黄菌中,半乳糖出现的频次最高。除了中性糖外,葡萄糖醛酸可能存在于鸡腿菇、黑木耳、鸡油菌和银耳等食用菌中,半乳糖醛酸可能存在杏鲍菇和榆黄菇等,进一步证明了酸性糖在食用菌多糖中存在的可能性。
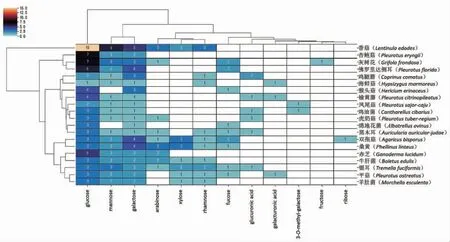
图8 基于二模矩阵的食用菌及其单糖组成的聚类分析Fig.8 Clustering analysis of fungi and their monosaccharide composition based on two-mode matrix
图9为基于二模矩阵的食用菌及其多糖结构类型的聚类分析。对于香菇多糖来说,最常见的类型为β-(1→3)-葡聚糖,还可能存在β-(1→6)-葡聚糖、α-(1→3)-葡聚糖和杂葡聚糖等,这一结果与2.3 节中葡萄糖出现频次较高的结果一致。灵芝中可能存在的多糖类型与香菇相似,除了β-(1→3)-葡聚糖,还可能存在α-(1→3)-葡聚糖和杂葡聚糖。海鲜菇多糖可能存在的多糖类型为半乳葡聚糖、岩藻甘露半乳聚糖和α-(1→4)-葡聚糖,此外还发现存在甘露聚糖。同样地,甘露聚糖也存在于平菇中。其它常见的食用菌,如黑木耳中出现β-(1→3)-葡聚糖的频次最高,此外还可能存在一些结构复杂的杂多糖。在双孢蘑菇中,除了可能存在葡聚糖外,还可能存在岩藻半乳聚糖和甘露葡聚糖。由此可发现,当食用菌多糖的分离纯化较为理想时,常常可以获得一些特定的多糖结构类型,如香菇多糖、灵芝多糖及黑木耳多糖大部分为β-(1→3)-葡聚糖结构,进一步佐证了食用菌多糖结构的相对有序性。

图9 基于二模矩阵的食用菌及其多糖结构类型的聚类分析Fig.9 Clustering analysis of fungi and their polysaccharide structure types based on two-mode matrix
图10为基于二模矩阵的食用菌多糖结构类型及其糖苷键类型的聚类分析。可以看出,对于β-(1→3)-葡聚糖和β-(1→6)-葡聚糖,其中常见的糖苷键类型有4 种,分别为β-(1→3)-Glcp、β-(1→6)-Glcp、β-T-Glcp 和β-(1→3,6)-Glcp。区别在于:β-(1→3)-葡聚糖中,β-(1→3)-Glcp 残基出现的频次最高,证明其可能存在于主链上,而β-(1→6)-Glcp 残基出现频次远低于其它3 种糖苷键,很可能位于侧链上。对于β-(1→6)-葡聚糖来说,β-(1→6)-Glcp 残基出现频率最高,而β-(1→3)-Glcp、β-T-Glcp 和β-(1→3,6)-Glcp 残基出现频次的比例接近1∶1∶1,说明其分支可能位于葡萄糖的O-3 位置,且β-(1→3)-Glcp、β-TGlcp 可能均为支链。对于半乳聚糖及杂半乳聚糖来说,其最常见的糖苷键类型为α-(1→2,6)-Galp、α-T-Fucp、α-(1→6)-Galp 和α-T-Galp。这4 种糖苷键中,α-(1→6)-Galp 出现频次最高,可能位于主链上,而分支可能位于半乳糖的O-2位。此外,对于甘露聚糖及杂甘露聚糖来说,出现频次最高的糖苷键类型分别为α-(1→4)-Manp、α-(1→6)-Manp、α-(1→2,6)-Manp 和α-(1→2)-Manp。根据出现频次来看,α-(1→4)-Manp 很可能存在于主链中。由此可以看出,食用菌多糖的糖苷键存在方式具有一定的规律性,一些特定的糖苷键类型及其组合方式上具有一定的共性,而这种规律性需要更多样本的食用菌多糖结构来进行归纳总结。

图10 基于二模矩阵的食用菌多糖结构类型及其糖苷键类型的聚类分析Fig.10 Clustering analysis of fungi structure types and their linkages patterns based on two-mode matrix
2 食用菌多糖结构分类
与其它天然产物来源多糖相比,食用菌多糖结构虽较为简单,但仍很难用一种或几种方法解析出其一级结构,必须结合大量的化学分析和现代仪器分析方法进行综合解析。在文献检索的基础上,继续对文献中常见食用菌多糖的分子质量、单糖组成、糖苷键类型、取代位置等信息进行归纳,通过与文献计量学结果的比较分析来探讨食用菌多糖结构的分类特征和规律[48]。
如表1所示,担子菌中最常见的多糖类型为α-葡聚糖和β-葡聚糖。就杂葡聚糖而言,其主链往往含有β-D-(1→6)-Glcp、β-D-(1→6)-Glcp和β-D-(1→3)-Glcp 残基,大部分在O-3 或O-6被β-D-Glcp、α-D-Glcp、α-D-Galp 和α-L-Fucp等残基取代。侧链还可能含有α-D-(1→6)-Galp、α-D-(1→4)-Galp、α-D-(1→3)-Galp、β-D-Manp和α-D-Xylp 等。杂半乳聚糖的主链结构通常为α-D-(1→6)-半乳聚糖,还可能含有Me-半乳糖,在O-2 处可能被L-Fucp、D-Manp、D-Galp 或3-O-α-Manp-α-L-Fucp 残基取代。杂甘露聚糖的主链常含有α-D-(1→3)-Manp、α-D-(1→2)-Manp和α-D-(1→6)-Manp 残基,在O-4 位被β-DXylp 和/或β-D-(1→3)-Xylp 取代。相比于这3类,杂聚糖类数量较多且结构复杂,其主链的可能存在葡萄糖、半乳糖、甘露糖、岩藻糖、阿拉伯糖、木糖和鼠李糖等单糖,糖苷键类型往往包含α-D-(1→2)-Glcp、α-D-(1→6)-Glcp、α-D-(1→2)-Galp 和α-D-(1→6)-Galp 等,这些信息为提升食用菌多糖结构研究提供一定的科学数据支持。
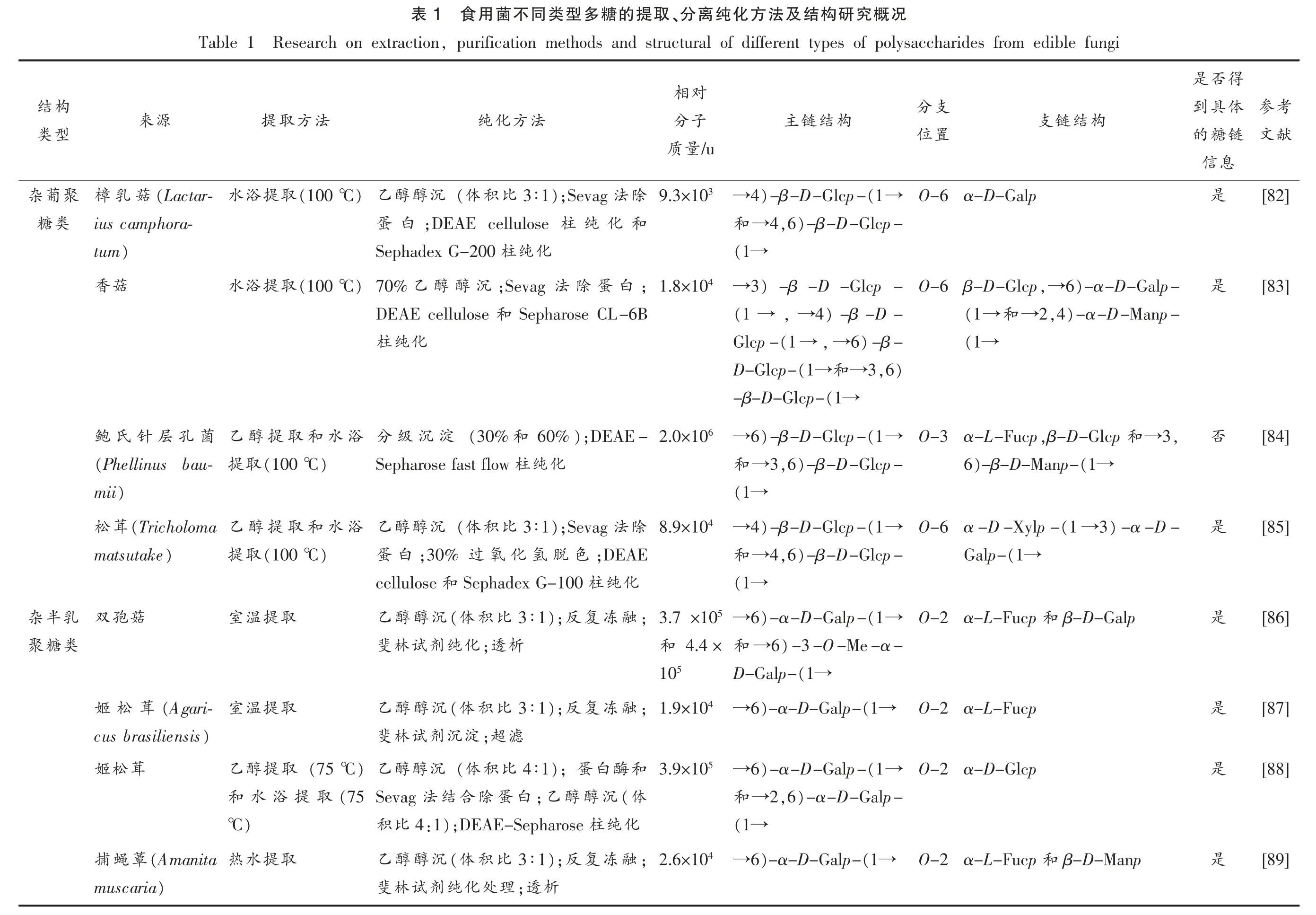
考参献文[82][83][84][85][86][87][88][89]得否是体具到链糖的息信是是否是是是是是况概究研构结及法方化纯离、分取提的糖多型类同不菌用食1表,purification methods and structural of different types of polysaccharides from edible fungi Research on extraction Table 1 对相支分构结链支构结链主子分法方化纯法方取提源来置位/u量质O-6 α-D-Galp(1→)-β-D-Glcp-4→3 9.3×10除法);Sevag 3∶1比积(体沉醇醇)乙(100 ℃取提浴水(Lactar-菇乳樟)-β-D-Glcp-,6 4→和和化纯柱;DEAE cellulose白蛋ius camphora-(1→化纯柱Sephadex G-200)tum)-α-D-Galp-6,→O-6 β-D-Glcp)-β-D-Glcp-3→4 1.8×10;白蛋除法;Sevag沉醇醇乙)70%(100 ℃取提浴水菇香)-α-D-Manp-,4 2→和(1→)-β-D-4,→(1 →Sepharose CL-6B和DEAE cellulose(1→)-β-6,→(1 →Glcp-化纯柱),6 3→和(1→D-Glcp-(1→-β-D-Glcp-,3→和,β-D-Glcp O-3 α-L-Fucp(1→)-β-D-Glcp-6→6 2.0×10);DEAE-60%和(30%淀沉级分浴水和取提醇乙菌孔层针氏鲍(1→)-β-D-Manp-6)-β-D-Glcp-,6 3→和化纯柱Sepharose fast flow)(100 ℃取提(Phellinus bau-(1→)mii)-α-D-3(1 →O-6 α-D-Xylp-(1→)-β-D-Glcp-4→4 8.9×10除法);Sevag 3∶1比积(体沉醇醇乙浴水和取提醇乙(Tricholoma茸松(1→Galp-)-β-D-Glcp-,6 4→和;DEAE色脱氢化氧过;30%白蛋)(100 ℃取提)matsutake(1→化纯柱Sephadex G-100和cellulose β-D-Galp和O-2 α-L-Fucp(1→)-α-D-Galp-6→5 3.7 ×10;融冻复);反3∶1比积(体沉醇醇乙取提温室菇孢双)-3-O-Me-α-6→和4.4×和析;透化纯剂试林斐(1→D-Galp-5 10 O-2 α-L-Fucp(1→)-α-D-Galp-6→4 1.9×10;融冻复);反3∶1比积(体沉醇醇乙取提温室(Agari-茸松姬滤;超淀沉剂试林斐)cus brasiliensis O-2 α-D-Glcp(1→)-α-D-Galp-6→5 3.9×10和酶白);蛋4∶1比积(体沉醇醇乙)(75 ℃取提醇乙茸松姬)-α-D-Galp-,6 2→和(体沉醇醇;乙白蛋除合结法Sevag(75取提浴水和(1→化纯柱);DEAE-Sepharose:1 4比积)℃β-D-Manp和O-2 α-L-Fucp(1→)-α-D-Galp-6→4 2.6×10;融冻复);反3∶1比积(体沉醇醇乙取提水热(Amanita蕈蝇捕析;透理处化纯剂试林斐)muscaria构结型类聚葡杂类糖乳半杂类糖聚

考参献文[90][91][92][93][94][95][96][97]得否是体具到链糖的息信是是是是是是是否支分构结链支置位O-2 β-D-Manp O-2 α-L-Fucp O-2 3-O-D-Manp-L-Fucp,βα-L-Fucp D-Manp 和,(1 →)-α-D-Manp-6 O-2 →Glcp和Fucp O-2 3-O-D-Manp-L-Fucp(1→,α-D-Galp-O-2 α-L-Fucp,(1→)-2-OAc-α-D-Galp-6 6)-α-Dα-D-Galp-(1 →4)-β-D-Glcp-Galp-(1 →α-D-,α-D-Galp 和(1 →Manp O-2 α-L-Fucp 3-O-Me-Rha和O-2 α-L-Fucp构结链主(1→)-α-D-Galp-6(1→)-α-D-Galp-6 6)-α-D-Galp-(1→(1→)-α-D-Galp-6(1→)-α-D-Galp-6(1→)-α-D-Galp-6(1→)-α-D-Galp-6(1→)-α-D-Galp-6对相子分/u量质4 →3.2×10→-→-4 →1.3×10 4 →1.5×10 4 →6.9×10 4 →1.2×10 4 →2.0×10法方化纯结法Sevag和酶白;蛋沉醇醇乙95%;DEAE沉醇级分;95%白蛋除合Sepharose-6和Sepharose fast flow析;透沉;醇化纯柱fast flow);DEAE-3∶1比积(体沉醇醇乙;Sepharose化纯柱Sepharose CL-6B柱;Sepharose CL-4B化纯柱CL-6B化纯复;反析3∶1);透比积(体沉醇醇乙滤;超理处化纯剂试林;斐融冻Sephacryl S-400和Sephacryl S-300化纯柱化纯柱;DEAE-cellulose沉醇醇乙除法);Sevag 1∶1比积(体沉醇醇乙);4∶1比积(体沉醇醇;乙析;透白蛋化纯Superdex-200 柱纯柱;DEAE-Sepharose fast flow滤超化纯柱;Sephacryl S-300化;DEAE-Sepharose fast flow沉醇醇乙;Sephacryl S-400 high reso-化纯柱化纯lution 柱水法方取提取提醇乙95%取提水热和取提醇乙95%取提水热和取提水热取提水热取提0.9% NaCl取提水热取提水热热和取提醇乙取提)1源来菇钉铆红血(Chroogomphis)rutilus蘑腿鸡(Flam-菇针金velumulina)tipes菇针金菌孔层(Fomitella frax-)inea(Ganoder-芝灵)ma atrum芝赤菇头猴表(续构结型类
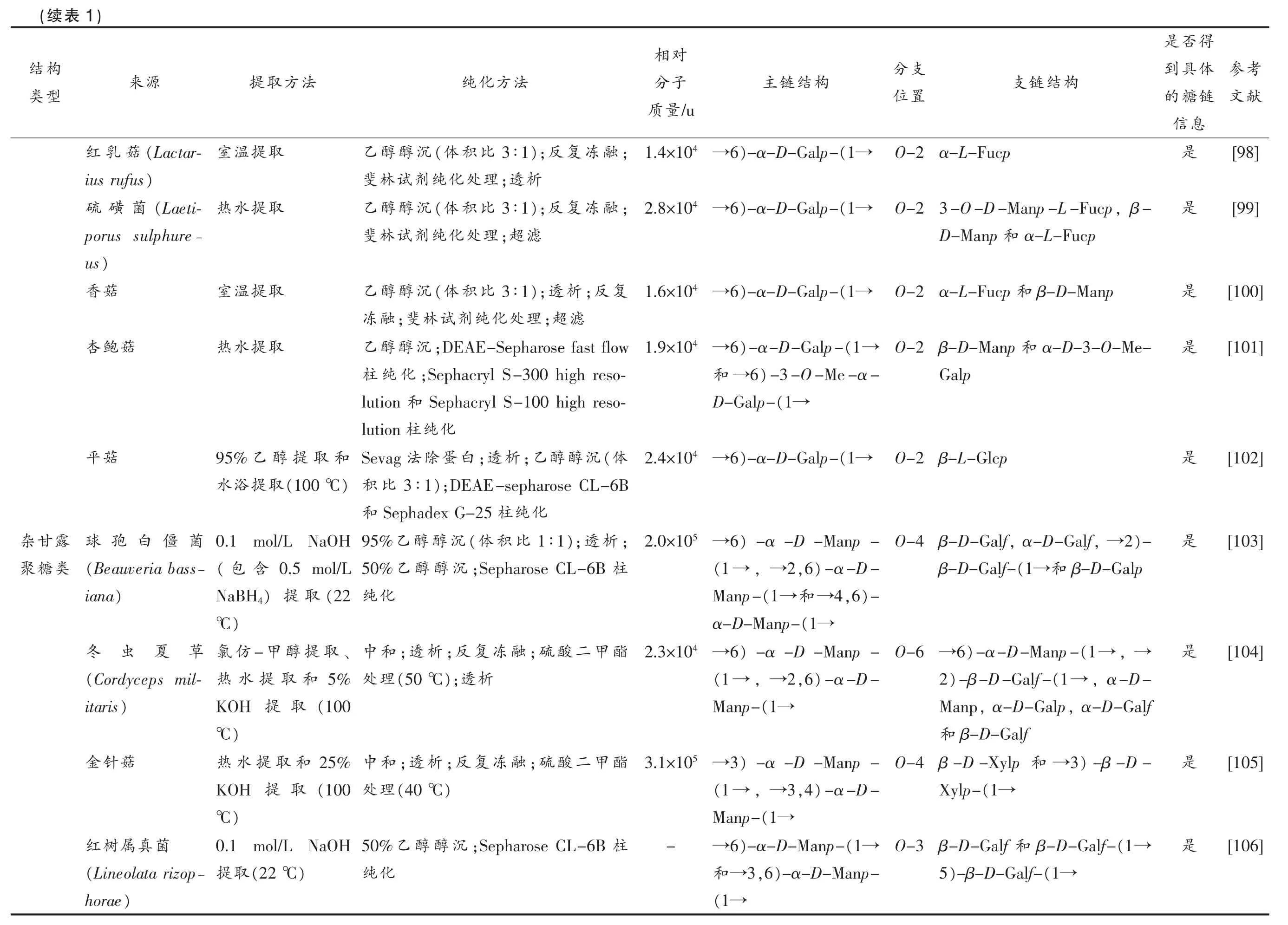
考参献文[98][99][100][101][102][103][104][105][106]得否是体具到链糖的息信是是是是是是是是是支分构结链支置位O-2 α-L-Fucp,β-O-2 3-O-D-Manp-L-Fucp α-L-Fucp和D-Manp β-D-Manp O-2 α-L-Fucp 和α-D-3-O-Me-和O-2 β-D-Manp Galp O-2 β-L-Glcp)-2,→,α-D-Galf O-4 β-D-Galf β-D-Galp和(1→β-D-Galf-,→6)-α-D-Manp-(1→O-6 →,α-D-(1→)-β-D-Galf-2,α-D-Galf,α-D-Galp Manp β-D-Galf和)-β-D-3→和O-4 β-D-Xylp(1→Xylp-(1→β-D-Galf-和O-3 β-D-Galf(1→)-β-D-Galf-5对相构结链主子分/u量质(1→)-α-D-Galp-6 4 →1.4×10(1→)-α-D-Galp-6 4 →2.8×10 6)-α-D-Galp-(1→4 →1.6×10(1→)-α-D-Galp-6 4 →1.9×10)-3-O-Me-α-6→和(1→D-Galp-(1→)-α-D-Galp-6 4 →2.4×10)-α-D-Manp-6 5 →2.0×10)-α-D-,6 2,→(1 →)-,6 4→和(1→Manp-(1→α-D-Manp-6)-α-D-Manp-4 →2.3×10)-α-D-,6 2,→(1 →(1→Manp-)-α-D-Manp-3 5 →3.1×10)-α-D-,4 3,→(1 →(1→Manp-(1→)-α-D-Manp-6→-)-α-D-Manp-,6 3→和(1→法方化纯;融冻复);反3∶1比积(体沉醇醇乙析;透理处化纯剂试林斐;融冻复);反3∶1比积(体沉醇醇乙滤;超理处化纯剂试林斐复;反析3∶1);透比积(体沉醇醇乙滤;超理处化纯剂试林;斐融冻;DEAE-Sepharose fast flow沉醇醇乙;Sephacryl S-300 high reso-化纯柱Sephacryl S-100 high reso-和lution化纯柱lution(体沉醇醇;乙析;透白蛋除法Sevag);DEAE-sepharose CL-6B 3∶1比积化纯柱Sephadex G-25和;析);透1∶1比积(体沉醇醇乙95%柱;Sepharose CL-6B沉醇醇乙50%化纯酯甲二酸;硫融冻复;反析;透和中析);透(50 ℃理处酯甲二酸;硫融冻复;反析;透和中)(40 ℃理处柱;Sepharose CL-6B沉醇醇乙50%化纯法方取提取提温室取提水热取提温室取提水热和取提醇乙95%)(100 ℃取提浴水0.1 mol/L NaOH 0.5 mol/L含(包(22取)提NaBH4)℃、取提醇-甲仿氯5%和取提水热(100取提KOH)℃25%和取提水热(100取提KOH)℃0.1 mol/L NaOH)(22 ℃取提)1源来(Lactar-菇乳红)ius rufus(Laeti-菌磺硫porus sulphure-)us菇香菇鲍杏菇平菌僵白孢球(Beauveria bass-)iana草夏虫冬(Cordyceps mil-)itaris菇针金菌真属树红(Lineolata rizop-)horae表(续构结型类露甘杂类糖聚
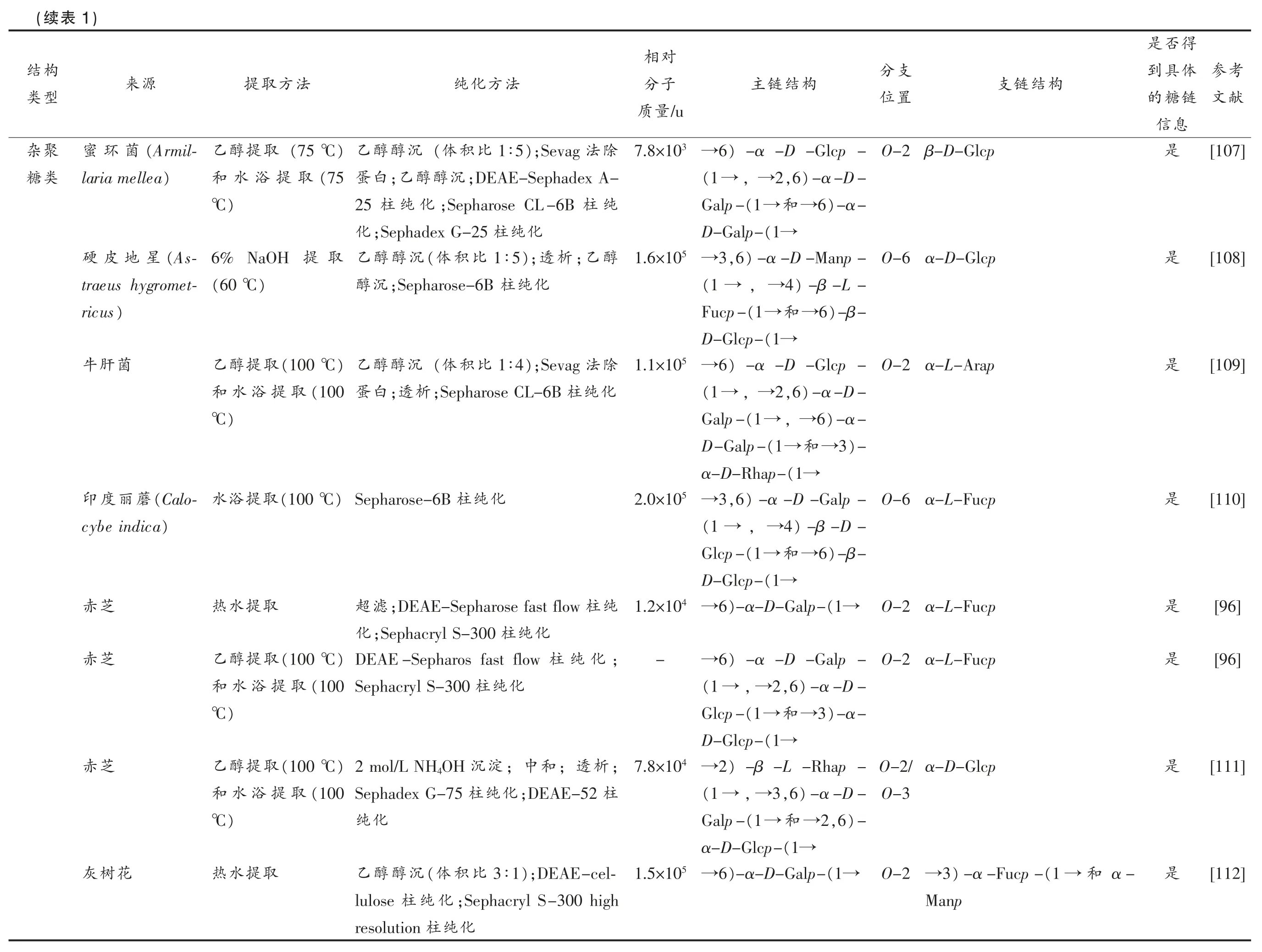
考参献文[107][108][109][110][96][96][111][112]得否是体具到链糖的息信是是是是是是是是α-和支分构结链支置位O-2 β-D-Glcp O-6 α-D-Glcp O-2 α-L-Arap O-6 α-L-Fucp O-2 α-L-Fucp O-2 α-L-Fucp α-D-Glcp O-2/O-3(1 →)-α-Fucp-3 O-2 →Manp对相构结链主子分/u量质6)-α-D-Glcp-3 →7.8×10)-α-D-,6 2,→(1 →)-α-6→和(1→Galp-(1→D-Galp-)-α-D-Manp-,6 3 5 →1.6×10)-β-L-4,→(1 →)-β-6→和(1→Fucp-(1→D-Glcp-)-α-D-Glcp-6 5 →1.1×10)-α-D-,6 2,→(1 →)-α-6,→(1→Galp-)-3→和(1→D-Galp-(1→α-D-Rhap-)-α-D-Galp-,6 3 5 →2.0×10)-β-D-4,→(1 →6)-β-→和Glcp-(1→D-Glcp-(1→(1→)-α-D-Galp-6 4 →1.2×10 6)-α-D-Galp-→-2,6)-α-D-,→(1 →3)-α-→和Glcp-(1→D-Glcp-(1→)-β-L-Rhap-2 4 →7.8×10)-α-D-,6 3,→(1 →)-,6 2→和(1→Galp-(1→α-D-Glcp-(1→)-α-D-Galp-6 5 →1.5×10法方化纯法方取提除1∶5);Sevag 法比积(体沉醇醇乙(75 ℃)取提醇乙;DEAE-Sephadex A-沉醇醇;乙白蛋(75取提浴水和纯柱;Sepharose CL-6B化纯柱25)℃化纯柱;Sephadex G-25化醇;乙析);透1∶5比积(体沉醇醇乙取提6% NaOH化纯柱;Sepharose-6B沉醇)(60 ℃除法);Sevag 1∶4比积(体沉醇醇乙)(100 ℃取提醇乙化纯柱;Sepharose CL-6B析;透白蛋(100取提浴水和)℃化纯柱)Sepharose-6B(100 ℃取提浴水纯柱;DEAE-Sepharose fast flow滤超取提水热化纯柱;Sephacryl S-300化;化纯DEAE-Sepharos fast flow 柱(100 ℃)取提醇乙化纯Sephacryl S-300 柱(100取提浴水和℃);析;透和;中淀沉OH 2 mol/L NH4)(100 ℃取提醇乙柱;DEAE-52化纯柱Sephadex G-75(100取提浴水和化纯)℃);DEAE-cel-3∶1比积(体沉醇醇乙取提水热;Sephacryl S-300 high化纯柱lulose化纯柱resolution)1源来(Armil-菌环蜜)laria mellea(As-星地皮硬traeus hygromet-)ricus菌肝牛(Calo-蘑丽度印)cybe indica芝赤芝赤芝赤花树灰表(续构结型类聚杂类糖

考参献文[113][114][115][116][117][118][119]得否是体具到链糖的息信是是是是是是是支分构结链支置位O-2 α-D-Galp O-2 β-D-Manp β-D-Manp O-2 α-D-Glcp 和α-L-Fucp和O-4 α-D-Glcp O-2 β-D-Manp O-2 β-D-Manp O-6 β-D-Glcp对相构结链主子分/u量质(1→)-α-D-Glcp-6 4 →4.0×10)-α-D-Glcp-,6 2→和,(1→(1→)-α-D-Galp-6 4 →1.3×10)-3-O-Me-α-6→和(1→D-Galp-6)-α-D-Galp-(1→4 →4.8×10)-α-D-Galp-6 5 →2.0×10)-α-D-,6 4,→(1 →)-β-6,→(1 →Galp-),4 3→和(1→D-Glcp-(1→-β-D-Glcp-6)-α-D-Galp-(1→4 →2.4×10 6)-3-O-Me-α-→和D-Galp-(1→(1→)-α-D-Galp-6 4 →3.5×10)-α-D-Glcp-,4 2→和(1→)-β-D-Glcp-3 6 →2.0×10)-β-D-,6 3,→(1 →)-α-6,→(1 →Glcp-)-3→和(1→D-Glcp-(1→α-L-Fucp-)1表法方化纯法方取提源来;融冻复);反3∶1比积(体沉醇醇乙)(85 ℃取提醇乙(Lep-蘑香脸花;白蛋除合结法Sevag和酶白蛋(80取提浴水和)ista sordida Sepharose CL-6B和DEAE cellulose)℃化纯柱纯柱;DEAE-Sepharose fast flow滤超和取提醇乙95%(Pleuro-菇珍秀;Sephacryl S-300 high resolution化)(100 ℃取提浴水)tus geesteranus化纯柱(体沉醇醇;乙析(80%);透沉醇级分取提水热耳侧达里罗佛比 5∶1);Gel-permeation chro-积化纯柱matography用次依淀);沉1∶5比积(体沉醇醇)乙(100 ℃取提浴水耳侧达里罗佛;析;透涤洗行进酮、丙醇乙蘑丽度印和属化纯柱Sepharose-6B(Leurotus florida Calocybe and var.indica APK2)复;反析3∶1);透比积(体沉醇醇乙取提醇-甲仿氯(Pleuro-菇珍秀滤;超理处化纯剂试林;斐融冻室3:1)和比积(体tus pulmonarius)取提温);Sepharose-1∶5比积(体沉醇醇)乙(100 ℃取提浴水菇尾凤化纯柱6B);4% NaOH 1∶5比积(体沉醇醇)乙(100 ℃取提浴水菌枞鸡柄粗;Sepharose-6B析;透沉醇醇;乙理处(Termitomyces化纯柱)robustus(续构型结类

考参献文[120][121][122])1表得否是对相体具到支分构结链支构结链主子分法方化纯法方取提源来链糖的置位/u量质息信是O-6 α-D-Galp)-β-D-Glcp-6→5 1.6×10);Sepharose-1∶5比积(体沉醇醇)乙(100 ℃取提浴水(Volvariella菇草4,6)-α-D-,→(1 →化纯6B 柱bombycina)6)-α-→和Manp-(1→D-Glcp-(1→是O-2 β-D-Manp(1 →)-α-D-Galp-→6 5 1.3×10);Sepharose-1∶5比积(体沉醇醇)乙(100 ℃取提浴水菇草(1→)-α-D-Glcp-,6和→2化纯柱6B是β-D-Glcp和O-6 α-L-Fucp)-β-D-Glcp-6→5 2.0×10化纯柱;Sepharose-6B沉醇醇)乙(100 ℃取提浴水菇草)-β-D-3,→(1 →)-β-,6 3,→(1→Glcp-),6 2→和(1→D-Glcp-(1→-α-D-Galp-(续构结型类
从已有的研究结果结合大量文献报道显示,绝大多数食用菌多糖中确实存在一定的规律性,且本团队已将“多糖结构的相对有序性”理论成功应用于一系列食用菌多糖的分离纯化[42,49]。结合手动文献检索对上述文献计量学的统计分析结果进行进一步佐证,将食用菌多糖的结构类型和规律归纳总结如下:分为葡聚糖类(homoglucan)及杂葡聚糖类(heteroglucan)、半乳聚糖类(heterogalactan)及甘露聚糖类(heteromannan)[50]。
2.1 葡聚糖类
这一类葡聚糖仅由葡萄糖残基构成,根据葡萄糖残基中异头碳的构型不同,可分为3 种:α-D-葡聚糖,β-D-葡聚糖和mixed α,β-D-葡聚糖[51]。在许多高等真菌及酵母中发现大量线性连接的α-D-葡聚糖结构,且连接方式具有多样性,即存在(1→3)-、(1→4)-或(1→6)-连接的α-D-Glcp残基构成的葡聚糖结构[52-53]。β-D-葡聚糖作为目前研究最多的真菌多糖活性组分,广泛分布于大量的食药用真菌中。研究发现,食用菌多糖中存在线性连接的β-D-(1→3)-葡聚糖[54]、β-D-(1→6)-葡聚糖[55]或带有分支的β-D-(1→3,6)-Glcp[56]结构。此外,还有研究显示,食用菌多糖中可能同时存在α-D-Glcp 和β-D-Glcp 的葡聚糖结构,如α-D-葡聚糖主链带有β-D-Glcp 侧链的葡聚糖结构,或β-D-葡聚糖主链带有α-D-Glcp 侧链的葡聚糖结构[57-60]。
2.2 杂葡聚糖类
食用菌多糖中除α-D-葡聚糖和β-D-葡聚糖外,还存在一类杂葡聚糖类,这类葡聚糖以DGlcp 残基构成主链,可能被β-D-Glcp、α-DGalp、α-L-Fucp 或α-D-Glcp 等不同的单糖残基取代构成侧链。根据多糖中除葡萄糖外,其它单糖种类和含量的不同,又可以分为以下5 种:
1)甘露葡聚糖(Mannoglucan)单糖组成除葡萄糖外,还含有部分甘露糖;
2)半乳葡聚糖(Galactoglucan)单糖组成除葡萄糖外,还含有部分半乳糖;
3)甘露半乳葡聚糖(Mannogalactoglucan)单糖组成除葡萄糖外,还含有部分半乳糖和甘露糖[61];
4)岩藻半乳葡聚糖(Fucogalactoglucan)单糖组成除葡萄糖外,还含有部分半乳糖和果糖[62];
5)木葡聚糖(Xyloglucan)单糖组成除葡萄糖外,还含有少部分木糖[63]。
现已证明从灵芝、香菇、金针菇、灰树花、黑木耳等天然食用菌中提取到的β-葡聚糖具有很好的免疫调节、抗肿瘤、抗氧化、抗衰老、抗病毒和改善糖尿病的作用[64-65]。而从冬虫夏草、双孢菇、姬松茸、竹荪等食用菌中得到的α-葡聚糖也具有一定的免疫调节活性[66-67]。
2.3 半乳聚糖类
从担子菌真菌(Basidiomycetes)中分离得到的结构多样的半乳聚糖类是目前报道较多的一类食用菌多糖。这类食用菌多糖以α-(1→6)-D-Galp构成主链,在Galp 的O-2 位连有分支,存在LFucp、D-Manp、D-Galp 或3-O-α-D-Manp-α-LFucp 等侧链结构。根据侧链结构和单糖种类的不同,可分为岩藻半乳聚糖(Fucogalactan)[68]、岩藻甘露半乳聚糖(Fucomannogalactan)[69]、甘露半乳聚糖(Mannogalactan)[70]及葡岩藻半乳聚糖(Glucofucogalactan)[71]等。
2.4 甘露聚糖类
子囊菌(Ascomycota)中较为常见的一类多糖为甘露聚糖类,它能与蛋白结合形成复合物,为食用菌细胞壁结构的组成部分。一般以(1→3)-或(1→6)-和(1→2)-连接的α-D-Manp 为主链,侧链连接有不同糖残基构成的分支,如α-D-Galp、α-D-Manp、α-D-Galf、α-D-GlcpA、β-D-Galf 或T-β-D-Xylp/β-D-(1→3)-Xylp。根据侧链结构的差异可分为半乳甘露聚糖(Galactomannan)[72]、葡半乳甘露聚糖(Glucogalactomannan)[73]、葡糖醛酸甘露聚糖(Glucuronomannan)[74]和木甘露聚糖(Xylomannan)[75]等。
3 结论与展望
食用菌多糖结构虽存在一定的相对有序性,但不可否认的是,也有相当一部分文献报道食用菌多糖结构不属于上述任何一类(如表2所示)。然而,从化学结构角度来看,多糖结构的“无序性”并非是绝对的,可能受限于以下几个方面而无法得到准确的多糖结构分析结果:1)食用菌多糖的单糖组成较为复杂,难以具体分析单个单糖之间的连接顺序;2)均一多糖中某种单糖或糖苷键含量过低,而无法通过常规的化学和仪器手段进行定量分析;3)多糖纯化不完全,或者仅通过测试其分子质量分布是否单一来评价其均一性,将“不均一的混合物”误当作“均一多糖”来进行结构解析;4)对多糖的结构类型认识不到位,无法科学地判断不同单糖、糖苷键的类型及其存在方式。需要提升现有方法技术和理论知识,不能简单“照抄”蛋白质和DNA 的分析手段,而是要扩展更多的多糖结构分析工具并结合其它先进技术[76-77]。例如:Amicucci 等[78]成功建立了一种灵敏、准确的化学方法,可将结构复杂的多糖氧化分解为低聚糖后进行液相色谱-质谱(LC-MS)检测,更适用于分析复杂混合物或非均一结构多糖。

考参献文[123][124][125]况概究研构结链糖本基及法方化纯离分取提的糖多杂菌用食2表,separation and purification methods and basic sugar chain structure of edible fungi heteropolysaccharide Overview of extraction Table 2 得否是体具到子分对相征特构结成组糖单法方离分取提源链糖的/u量质息信否)-α-D-3→由链,支糖聚葡)-3(1→α-D-为链主-糖露甘和糖萄葡;1 mol/L HCl取提温室液溶1 mol/L NaOH成构(1→Manp-淀沉醇;乙和中否)-,6 2,→(1→)-α-D-Galp-6:→型类键苷糖的有含4 1.1×10糖萄、葡糖露、甘糖乳半和(30%沉醇级);分(100 ℃取提浴水)-β-6,→(1→)-α-D-Manp-,6 2,→(1→α-D-Galp-糖伯拉阿和;High-化纯柱);DEAE-Sepharose 60%(1→)-β-D-4-O-Me-Glcp-6→和(1→D-Glcp-化纯柱Resolution Sephacryl S-300否)-β-4,→(1→)-β-D-Glcp-3:→型类键苷糖的有含5 2.0×10糖乳、半糖露、甘糖萄葡化纯柱Superdex G-200)-α-D-2,→(1→)-β-D-Glcp-6,→(1→D-Glcp-酸醛糖乳半和)-α-D-4,→(1 →)-α-D-Manp-4,→(1 →Manp-)-β-D-,6 4,→(1→)-β-D-Glcp-,6 3,→(1→Galp-,β-(1→)-α-D-Galp-4(1→,β-D-Glcp-(1→Glcp-)3(1→β-D-Glcp-和(1→)-β-D-Glcp-3(1→D-Glcp-(1→)-β-D-Glcp-6(1→β-D-Glcp-(1→-β-D-Glcp-(1→)-α-D-Galp-4(1→)-α-D-GalpA-4来菌肝牛菌肝牛芝赤
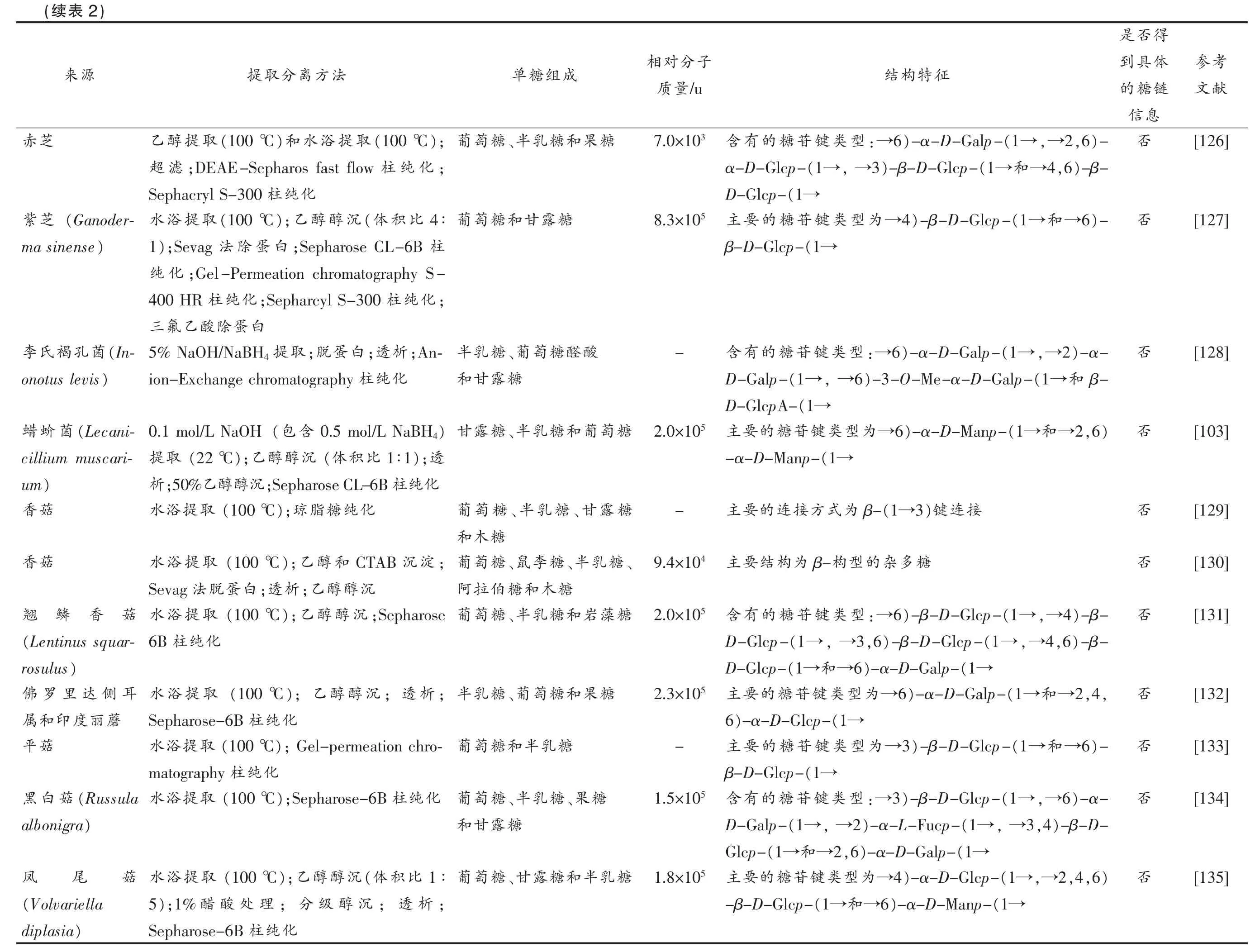
考参献文[126][127][128][103][129][130][131][132][133][134][135]得否是体具到链糖的息信否否否否否否否否否否否征特构结)-,6 2,→(1→)-α-D-Galp-6:→型类键苷糖的有含)-β-,6 4→和(1→)-β-D-Glcp-3,→(1→α-D-Glcp-(1→D-Glcp-)-6→和(1→)-β-D-Glcp-4→为型类键苷糖的要主(1→β-D-Glcp-2)-α-,→6)-α-D-Galp-(1→:→型类键苷糖的有含β-和6)-3-O-Me-α-D-Galp-(1→,→D-Galp-(1→D-GlcpA-(1→),6 2→和(1→)-α-D-Manp-6→为型类键苷糖的要主(1→-α-D-Manp-接连)键3(1→β-为式方接连的要主糖多杂的型构β-为构结要主)-β-4,→(1→)-β-D-Glcp-6:→型类键苷糖的有含)-β-,6 4,→(1→)-β-D-Glcp-,6 3,→(1→D-Glcp-(1→)-α-D-Galp-6和→(1→D-Glcp-,,4 2→和(1→)-α-D-Galp-6→为型类键苷糖的要主(1→)-α-D-Glcp-6)-6→和(1→)-β-D-Glcp-3→为型类键苷糖的要主(1→β-D-Glcp-)-α-6,→(1→)-β-D-Glcp-3:→型类键苷糖的有含)-β-D-,4 3,→(1→)-α-L-Fucp-2,→(1→D-Galp-(1→)-α-D-Galp-,6 2→和(1→Glcp-),6,4 2,→(1→)-α-D-Glcp-4→为型类键苷糖的要主(1→)-α-D-Manp-6和→(1→-β-D-Glcp-子分对相/u量质3 7.0×10 5 8.3×10-5 2.0×10-4 9.4×10 5 2.0×10 5 2.3×10-5 1.5×10 5 1.8×10成组糖单糖果和糖乳、半糖糖露甘和糖酸醛糖萄、葡糖糖露糖萄葡和糖乳、半糖糖露、甘糖乳、半糖糖、糖乳、半糖李、鼠糖糖木和糖伯糖藻岩和糖乳、半糖糖果和糖萄、葡糖糖乳半和糖糖、果糖乳、半糖糖露糖乳半和糖露、甘糖萄萄乳甘露萄木萄拉萄乳萄萄甘萄法方离分取提葡);(100 ℃取提浴水)和(100 ℃取提醇乙;化纯柱;DEAE-Sepharos fast flow滤超化纯柱Sephacryl S-300葡4∶比积(体沉醇醇);乙(100 ℃取提浴水柱;Sepharose CL-6B白蛋除法);Sevag 1;Gel-Permeation chromatography S-化纯;化纯柱;Sepharcyl S-300化纯柱400 HR白蛋除酸乙氟三半;An-析;透白蛋;脱取提5% NaOH/NaBH4和化纯ion-Exchange chromatography 柱甘)0.5 mol/L NaBH4含(包0.1 mol/L NaOH);透1∶1比积(体沉醇醇);乙(22 ℃取提化纯柱;Sepharose CL-6B沉醇醇乙;50%析葡化纯糖脂);琼(100 ℃取提浴水和葡;淀沉CTAB和醇);乙(100 ℃取提浴水阿沉醇醇;乙析;透白蛋脱法Sevag葡;Sepharose沉醇醇);乙(100 ℃取提浴水化纯柱6B半;析;透沉醇醇);乙(100 ℃取提浴水化纯柱Sepharose-6B葡);Gel-permeation chro-(100 ℃取提浴水化纯柱matography葡化纯柱);Sepharose-6B(100 ℃取提浴水和葡1∶比积(体沉醇醇);乙(100 ℃取提浴水;析;透沉醇级;分理处酸醋);1%5化纯柱Sepharose-6B 2)表(续源来芝赤(Ganoder-芝紫)ma sinense(In-菌孔褐氏李onotus levis)(Lecani-菌蚧蜡cillium muscari-)um菇香菇香菇香鳞翘(Lentinus squar-)rosulus耳侧达里罗佛蘑丽度印和属菇平(Russula菇白黑)albonigra菇尾凤(Volvariella)diplasia
此外,食用菌多糖结构的相对有序性在生物合成方面也存在很大争议。现代生物学研究已证实,糖基转移酶(GTs)在细胞中的合成和转运可导致多糖复杂性和结构多样性[79]。在过去10年中,研究人员在揭示和克隆编码多糖合成GTs 的基因方面取得显著进展,认为多糖合成过程中最关键的酶为具有高度底物专一性的GTs[80]。然而,绝大多数GT 在结构和功能上仍不明确,它们在高尔基体膜中如何高效地合成复杂的高度分支的多糖结构仍未知。目前涉及多糖生物合成方面的研究还有所欠缺,应进一步探究食用菌多糖是否存在结构精确合成的可能性,为整个理论提供更加完整、确凿的证据。尽管以目前的技术手段还无法解决这个问题,但已有人提出:多糖的延长和分化均由质膜上糖磷脂酰肌醇锚定蛋白所锚定的聚糖转移酶实现[81]。一旦这种特异性的聚糖转移酶在食用菌中发现,将进一步揭开多糖在生物体内合成机理的神秘面纱。
综上,进一步探索和辩证看待食用菌多糖结构的相对有序性规律,有助于提升食用菌多糖结构的正确认识,为今后食用菌乃至其它天然产物来源的多糖的技术研究和产品开发提供新思路。未来对食用菌多糖生物合成途径和调控机制的深入研究,将有望让人们认识到“故事”的真相。
