蛋氨酸和半胱氨酸对丝尾鳠生长性能、蛋氨酸代谢和蛋白质代谢的影响
2021-09-08张新党李光彬王秋梅苏林海李必为邓君明
张新党,李光彬,王秋梅,孙 彦,黄 汉,苏林海,李必为,邓君明
(云南农业大学动物科学技术学院,昆明 650201)
0 引言
蛋氨酸(Met)又名甲硫氨酸,属中性氨基酸,与生物体内各种含硫化合物的代谢有重要关系,是许多养殖鱼类的第一限制性氨基酸[1],在鱼类生长发育过程中具有重要的生理生化功能。研究表明,在鱼类配合饲料中添加适量的Met不仅可以有效地提高其饲料利用率、生长速率以及其免疫能力,还能降低其养殖成本[2-4]。Met 在动物体内大部分直接参与蛋白质的合成,当饲料中半胱氨酸(Cys)的量不足以满足动物合成蛋白所需时,Met 便会转化为合成蛋白质所需的Cys[5]。但由于胱硫醚合成酶的不可逆反应阻止了Cys重新转化为高半胱氨酸与丝氨酸,因此Cys 又不能完全满足动物对于Met的需求量[6]。因此,饲料中Cys的含量也会影响鱼类对Met的需求量。
丝尾鳠(Hemibagrus wyckioides)隶属于鲇形目、鲿科、鳠属,又名长胡子鱼,原产于老挝、越南、泰国等东南亚各国河流水域,在中国则主要分布在云南省境内澜沧江下游支流[7-8]。近年来,随着丝尾鳠人工繁殖和育苗技术的突破以及多种养殖规模的推广,极大地推动了丝尾鳠养殖业的兴起,其养殖规模不断扩大,现已在云南文山、河口、景洪等地进行推广养殖,养殖面积大于667 hm2,而且取得了良好的经济效益和社会效益[9-10]。然而,迄今有关丝尾鳠营养生理鱼营养需求方面的研究还比较少,目前也仅有丝尾鳠蛋白质需求的研究报道[11]。本课题组前期的两批试验结果表明:于基础饲料(Met 含量0.5%)中添加不同水平Met(0、0.5%、0.7%、0.9%、1.1%、1.3%、1.5%)对丝尾鳠生长性能和大部分生理生化指标没有显著影响。然而,传统水产动物营养学认为Met 是鱼类的必需氨基酸(EAA),Met 缺乏会显著降低鱼类的生长性能。同时,本课题组前期以纯化原料配制4种饲料(对照组、蛋氨酸缺乏组、半胱氨酸缺乏组、蛋氨酸+半胱氨酸缺乏组)饲喂丝尾鳠幼鱼,但丝尾鳠幼鱼对于这四种饲料并无摄食现象。本试验采用低完整蛋白高晶体氨基酸基础饲料(对照组)中分别扣除Met、Cys、Met+Cys 的形式,配制四组等氮等能饲料[Con、-Met、-Cys、-(Met+Cys)]饲喂丝尾鳠,分析其生长性能和生理生化指标,证实Met 是否为丝尾鳠的EAA,并研究丝尾鳠对Met的代谢特点和利用能力。
1 材料与方法
1.1 试验饲料
本试验采用低蛋白高晶体氨基酸基础饲料(Con)中分别扣除Met、Cys、Met+Cys 的形式,以鱼粉、豆粕和晶体氨基酸作为饲料蛋白源,鱼油、豆油和大豆卵磷脂作为脂肪源,饲料原料中的晶体氨基酸用谷氨酸和天冬氨酸进行等氮调节,除Met和Cys水平不同外,其余的EAA和非必需氨基酸(NEAA)按照丝尾鳠幼鱼全鱼氨基酸组成模式,用6 mol/L NaOH 溶液调节饲料pH 至中性,配制4 种等氮等能饲料,分别为Con、(-Met)、(-Cys)、-(Met+Cys),以确定Met 是否为丝尾鳠EAA。试验饲料组成及营养成分和饲料氨基酸组成见表1和表2。
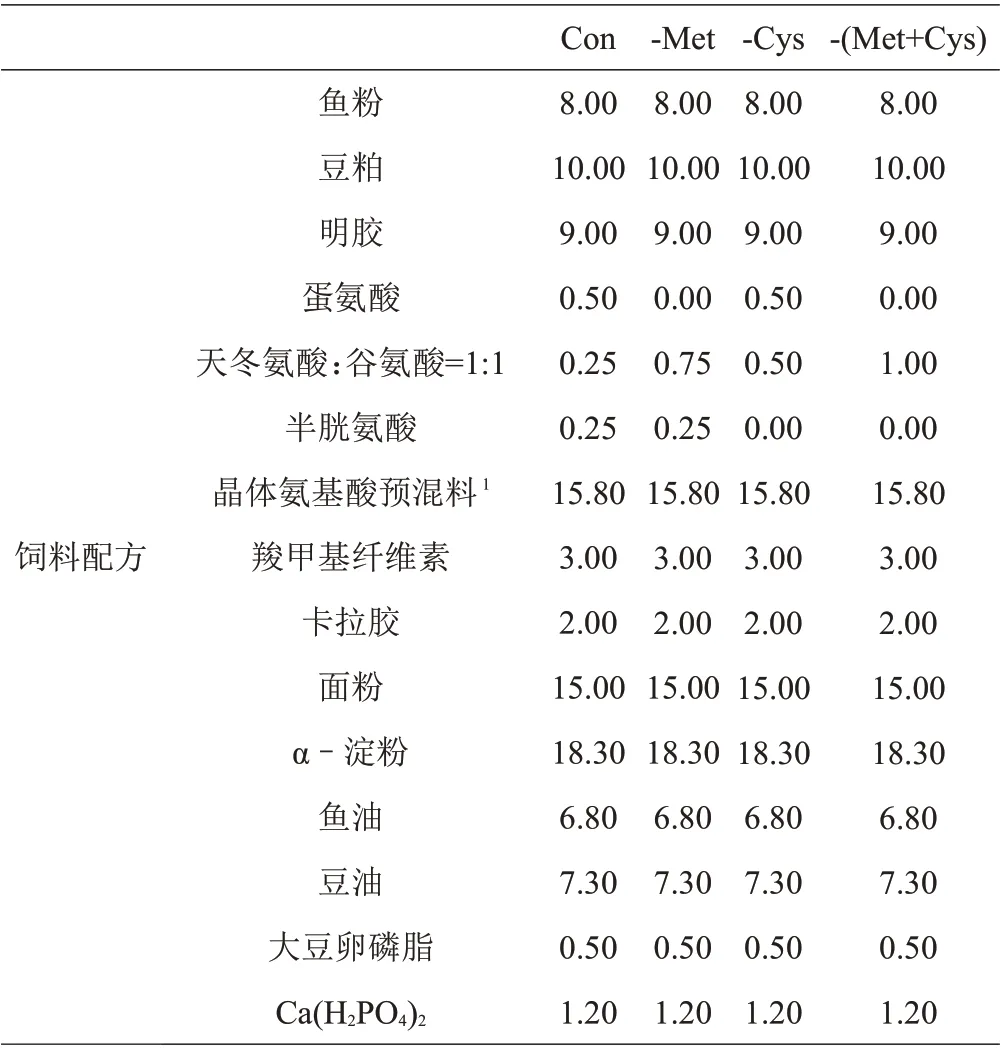
表1 试验饲料配方和营养组成(干物质基础) %
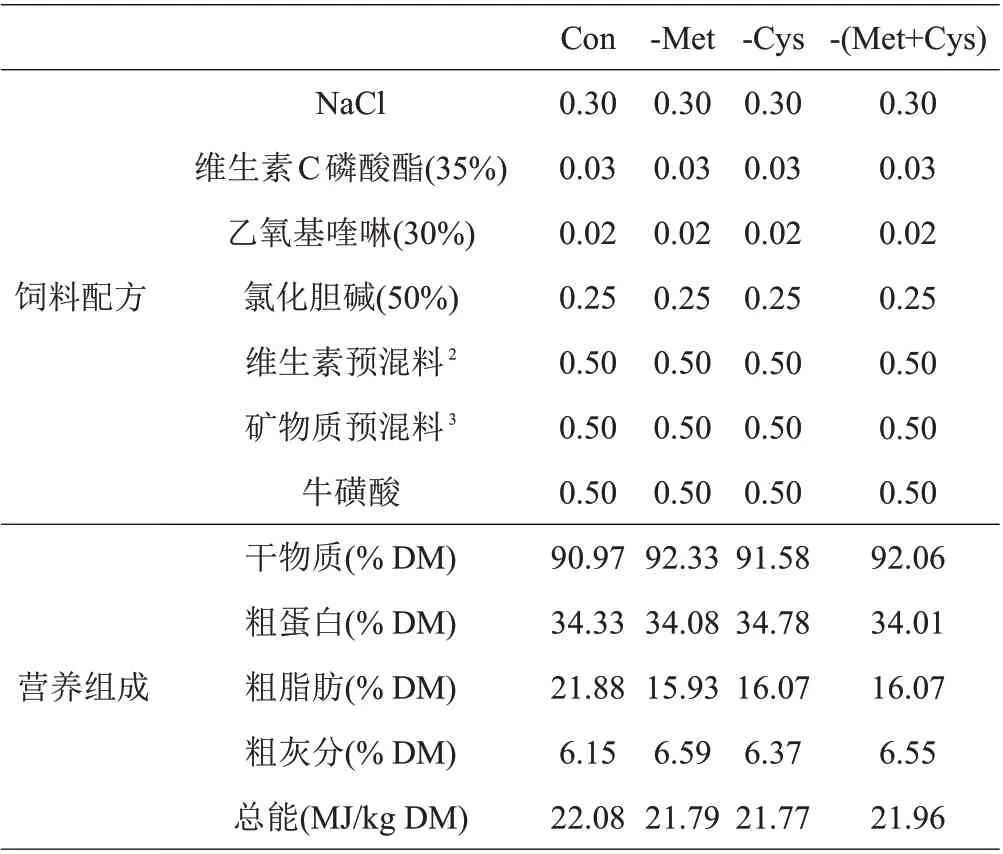
续表1
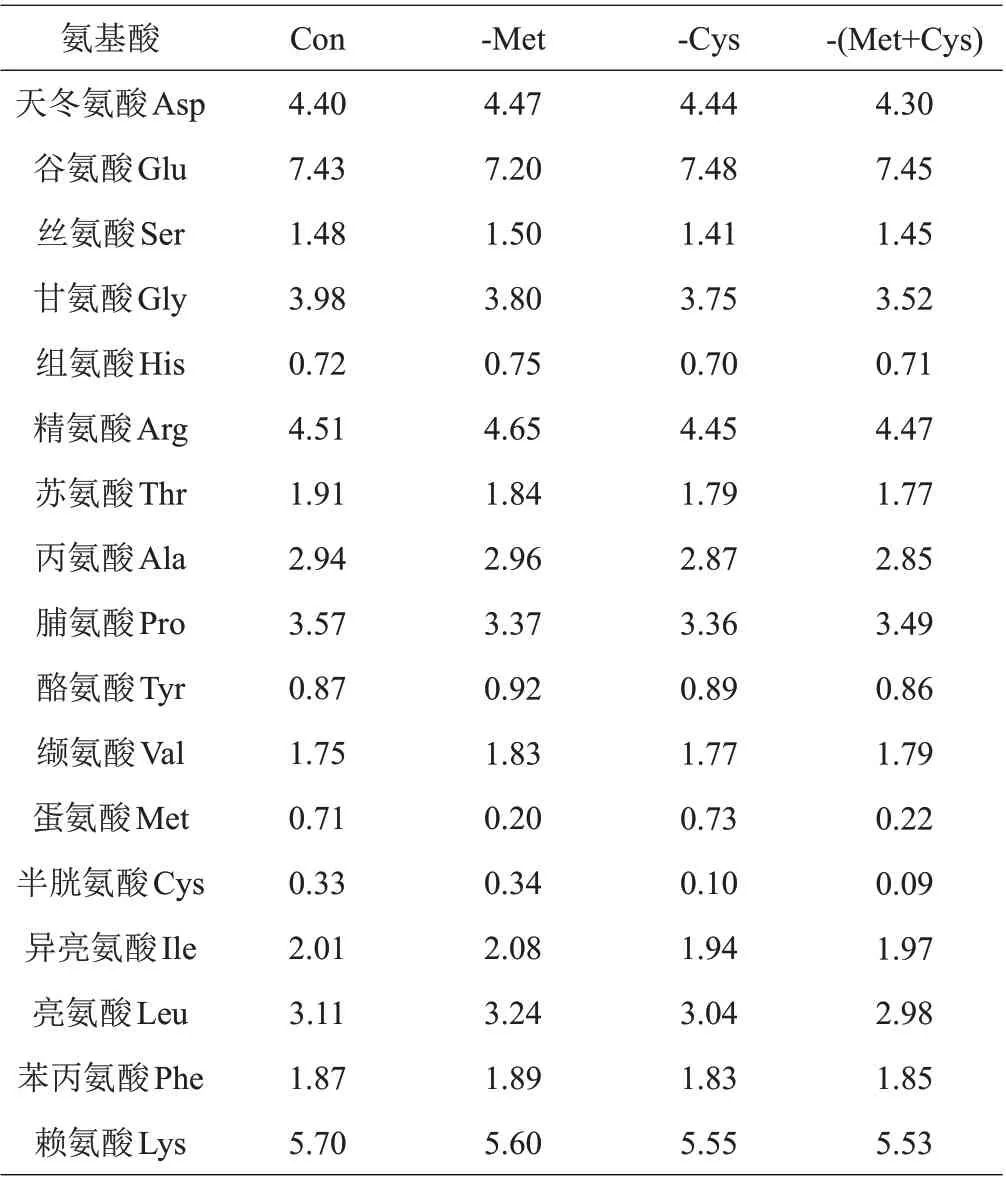
表2 试验饲料氨基酸组成 %
配制饲料前,所有原料必需经过粉碎机(SFSP 系列,昆明华明粮油饲料设备厂生产)粉碎,且全部过60目筛。将所有粉碎好的饲料原料按表1饲料配方混合均匀,然后再加入鱼油、豆油和大豆卵磷脂(先溶于鱼油和豆油中),手工将油脂微小颗粒搓散,再加入30%左右蒸馏水使粉状饲料形成硬团,最好于颗粒饲料机(KS-180,江苏晶谷米机有限公司生产)中将饲料挤压成直径1.0 mm的条状,并在40℃鼓风烘箱中干燥12 h,置于-20℃冰箱保存备用。
1.2 试验动物及饲养管理
试验用丝尾鳠(Hemibagrus wyckioides)幼鱼为当年人工培育的同一批苗种,由云南省西双版纳州水产技术推广站提供。试验于云南农业大学动物科学技术学院水产养殖学实验室中进行。试验前,试验幼鱼暂养2周适应环境,每天饲喂2次。养殖用水为曝气自来水,采用循环流水系统,循环系统采用机械和生物介质过滤。
试验开始前禁食24 h,选择大小均匀(初始体重约2.70 g)、体格健壮的丝尾鳠鱼苗480尾,随机分配于12个玻璃缸(4个试验处理组,每组3个重复),每缸(0.5 m× 0.5 m × 1.0 m)40 尾鱼。每天早晚投饵2 次(07:00,17:00),达饱食水平,早上投喂结束后30 min,吸出残饵及粪便。试验共进行8 周,整个养殖试验期间均采用24 h 连续充氧,自然光照,水温始终保持在27~29℃。
1.3 样品收集
养殖试验结束后,禁食24 h,于每缸随机取6尾鱼作为全鱼样品,分析测定鱼体常规组成。此外,每缸随机选取3 尾鱼,解剖后将完整的肝脏样品置于-80℃下保存;每缸随机选取3 尾鱼,解剖后将肠取出,剥去脂肪,去除其内容物后装鱼密封袋中,放于-80℃保存。
1.4 分析测试
试验持续养殖8周,结束前一天停止投喂,统计每个玻璃缸鱼体总重和数量,记录每个玻璃缸投喂饲料的总用量,计算平均代谢体重、摄食率、体增重、日增重系数以及饲料系数、蛋白质效率。各生长指标的计算分别见公式(1)~(6)。


鱼体和饲料常规成分分析参照AOAC法。其中:水分测定用恒温烘箱在105℃下烘至恒重;粗蛋白质含量的测定采用全自动凯氏定氮仪(JK9830,济南精密科学仪器仪表有限公司测定);粗脂肪的测定采用索氏提取法(以石油醚为溶剂)测定;粗灰分的测定为箱式电阻炉(SX-410,北京市永光明医疗仪器有限公司)550℃灼烧法(16 h)。总能的测定采用氧弹式能量仪(ZDHW-6,鹤壁市华泰仪器仪表有限公司)测定。饲料中氨基酸的组成分析采用Venusil AA氨基酸分析方法(酸水解法),用高效液相色谱仪(LC-20AT,日本岛津公司)测定。
肝脏、肠道的生化指标均采用南京建成生物工程研究所生产的试剂盒,按照使用说明进行操作。其中,肝脏和肠道中胰蛋白酶采用紫外比色法,淀粉酶(AMS)采用碘-淀粉比色法,脂肪酶(LPS)、二糖酶采用比色法。肝脏碱性磷酸酶(ALP)连续监测法,谷草转氨酶(AST)、谷丙转氨酶(ALT)采用赖氏法,谷胱甘肽还原酶(GR)采用紫外比色法,超氧化物歧化酶(SOD)采用黄嘌呤氧化酶法,溶菌酶(LZM)采用比浊法,谷氨酸脱氢酶(GDH)、腺苷单磷酸脱氨酶(AMPD)、蛋氨酸腺苷转移酶(MAT)、蛋氨酸合成酶(MS)、甜菜碱高半胱氨酸甲基转移酶(BHMT)和胱硫醚β合酶(CBS)均采用双抗体夹心法,丙二醛(MDA)、总抗氧化能力(TAC)、过氧化物酶(POD)、谷胱甘肽过氧化物酶(GSH-Px)、谷氨酰转肽酶(GGT)、过氧化氢酶(CAT)均采用可见光法。组织中蛋白质含量采用考马斯亮兰法测定。
1.5 统计分析
所有试验结果均表示为平均值±标准误(means±SEM)。所有百分率数据分析统计前均先经Arcsine转换。采用单因素方差分析(ANOVA),当处理组之间存在显著性差异(P<0.05)时,用Duncan's 检验进行多重比较分析。所有统计分析均采用SPSS 17.0 软件进行分析。
2 结果
2.1 生长性能
由表3可知,养殖试验期间,各饲料处理组丝尾鳠成活率为98.33%~99.17%,不同饲料处理组间成活率无显著性差异。-Cys 丝尾鳠增重率、日增重系数、摄食率和蛋白质效率均显著高于其他饲料处理组;反之,Con和-Cys饲料系数显著低于-Met和-(Met+Cys)。
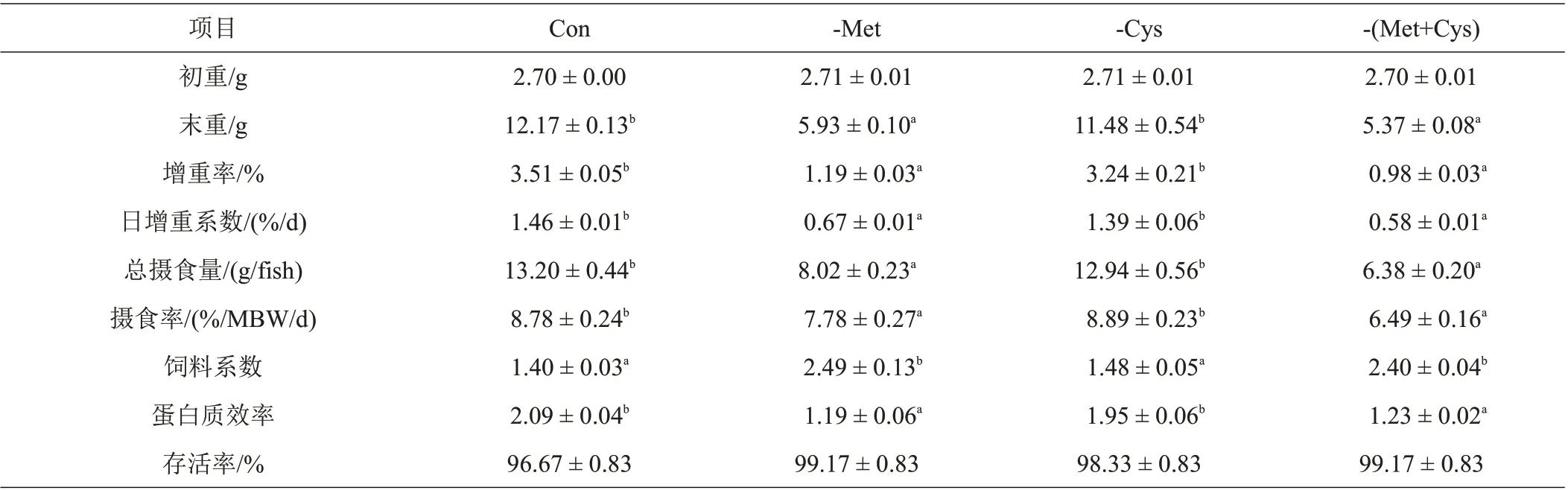
表3 不同饲料处理对丝尾鳠生长性能的影响
2.2 消化酶活性
由表4 可知,Con 和-Met 丝尾鳠胃蛋白酶活性显著高于-Cys;-Met 和-(Met+Cys)肝脏脂肪酶活性显著低于Con;-Cys肝脏二糖酶活性则显著高于Con,而与其他饲料处理组无显著性差异。不同饲料处理组间肠道胰蛋白酶、脂肪酶、淀粉酶、二糖酶的活性以及肝脏胰蛋白酶、淀粉酶的活性均无显著性差异。
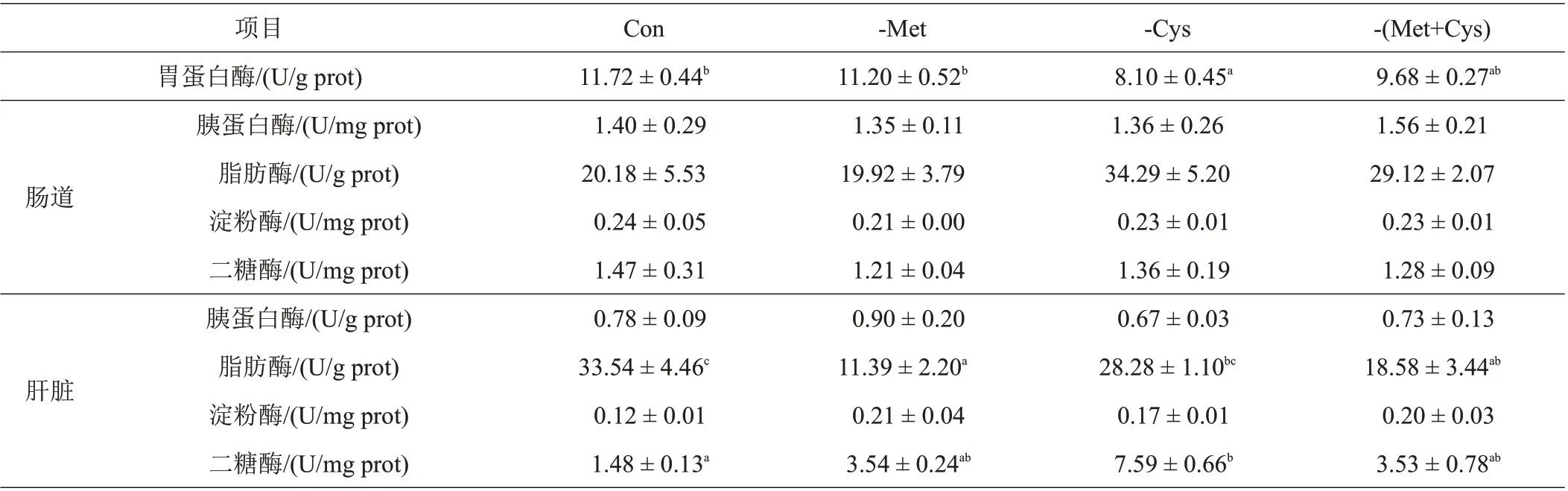
表4 不同饲料处理对丝尾鳠消化酶活性的影响
2.3 蛋白质代谢
由表5 可知,Con 丝尾鳠肝脏AMPD 显著低于-(Met+Cys);而-(Met+Cys)肝脏ALT 活性则显著高于其他饲料处理组;不同处理组间肝脏GDH 和AST 活性则没有显著性差异。-Met 肝脏MAT 活性显著高于Con 和-Cys;类似地,-Met 肝脏CBS 显著高于-Cys 和-(Met+Cys);-(Met+Cys)肝脏BHMT 活性显著高于Con 和-Cys,与-Met 无显著性差异;而Con 组MS 显著高于-Cys,与其他饲料处理组无显著性差异。

表5 不同饲料处理对丝尾鳠肝脏蛋氨酸和蛋白质代谢关键酶活性的影响
2.4 抗氧化功能
由表6 可知,Con 丝尾鳠肝脏SOD 活性显著低于-(Met+Cys);-Met肝脏POD活性和T-AOC活性显著高于-Cys 和-(Met+Cys);类似地,-Met 肝脏CAT 和GR活性均显著高于其他饲料处理组;Con 和-(Met+Cys)肝脏GSH-Px 活性显著高于-Met 和-Cys;-Cys 肝脏AKP 活性显著高于-(Met+Cys)和Con;而Con 组MDA含量则显著低于其他饲料处理组。不同饲料处理组LZM活性没有显著性差异。
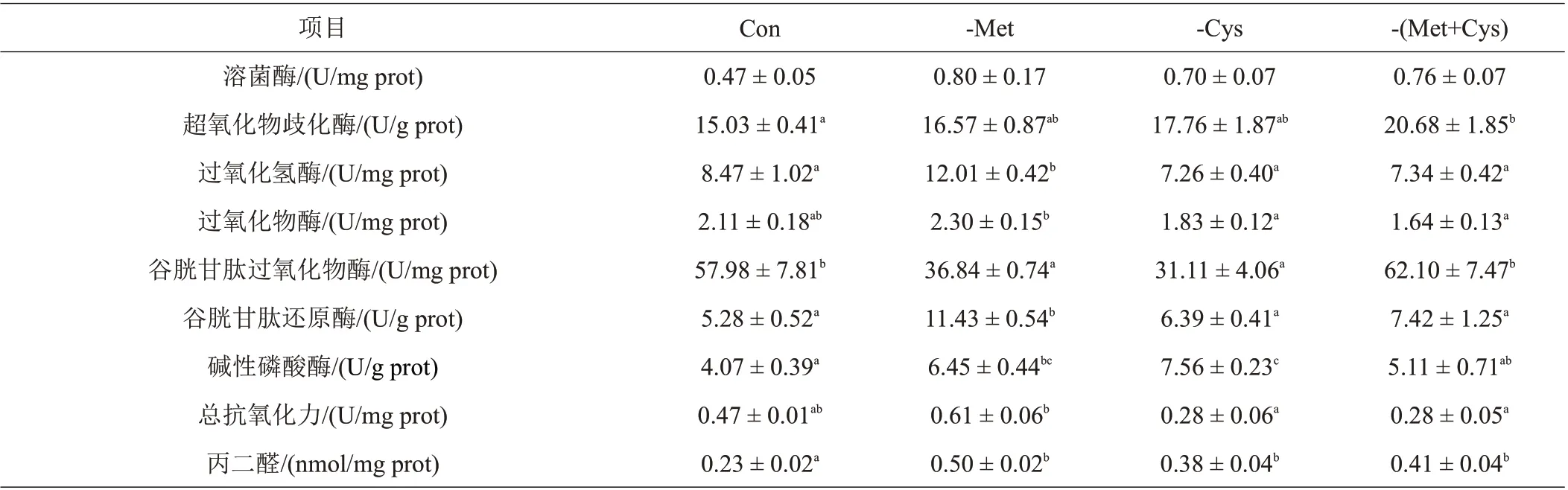
表6 不同饲料处理对丝尾鳠肝脏抗氧化能力的影响
2.5 鱼体组成
由表7 可知,-Met 和-(Met+Cys)鱼体水分显著高于Con 和-Cys,相反地,-Met 和-(Met+Cys)鱼体粗蛋白、粗脂肪和粗灰分则显著低于Con和-Cys。
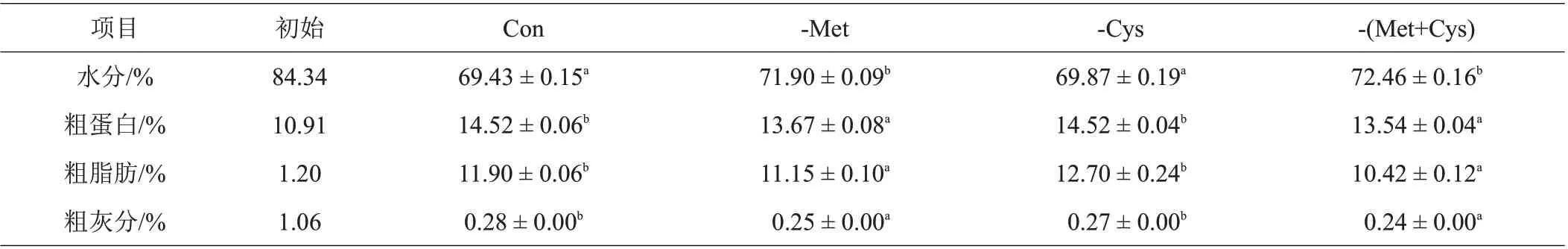
表7 不同饲料处理对丝尾鳠鱼体组成的影响
3 讨论
3.1 不同饲料处理对丝尾鳠生长性能的影响
研究表明,当饲料中Met含量不足时,会降低饲料转化率和鱼体增重率[13],造成鱼类生长速度减缓或停滞、肾脏肿大等负面影响,最终导致其肝坏死或纤维化[14]。而Cys 作为一种NEAA,可由Met 转化而成,进而参与组成蛋白质,因此鱼类需要足够的Met 以及Cys 来满足蛋白质合成及其他一些生理活动的需要。本试验中,扣除Met和Met+半胱氨酸组丝尾鳠的饲料转化率和鱼体增重率均显著低于其他饲料处理组,其原因可能是饲料中Met含量的不足造成其氨基酸不平衡,进而对其饲料利用率和生长造成影响[15]。研究表明,饲料中Met 含量不足时,显著降低了大菱鲆[16](Scophthalmus maximus)、瓦氏黄颡鱼[17](Pelteobagrus vachelli)、吉富罗非鱼[18](Oreochromis niloticus)等的生长性能和饲料利用率;类似地,周贤君等[19]研究发现,低含量Met不仅降低了异育银鲫的生长性能和特定生长率,还使其蛋白沉积率显著下降。以上结果均表明饲料中Met 的缺乏会严重抑制鱼类的生长,而补充适量的Met 则明显改善其生长性能,同时Met 还明显影响了尖齿胡鲶[20](Clarias gariepinus)、斑点叉尾鮰[21](Ietalurus Punetaus)和印度囊鳃鲶[22](Heteropneustes fossilis)的生长状态。本课题组前期两批试验结果显示添加不同水平Met并未对丝尾鳠的生长造成显著性差异,这是由于前期试验饲料中本底Met 的含量均为0.5%,而本课题组后期试验也表明丝尾鳠幼鱼的最适Met 需要量为0.5%左右,因此本底中的Met 含量已能满足丝尾鳠幼鱼的生长需求,因此造成添加不同水平Met 并未对其生长以及生理生化指标造成显著性差异。而本试验中,当扣除Met和Cys时,饲料本底中所含Met为0.2%左右,造成丝尾鳠幼鱼的生长性能和饲料利用率显著下降,可说明Met 是丝尾鳠幼鱼的EAA。
3.2 不同饲料处理对丝尾鳠消化酶活性的影响
鱼类的生长依赖于对营养物质的消化和吸收,鱼类消化道中的消化酶活性则是衡量其消化吸收能力的重要指标。而消化器官则包括肠道和肝胰脏,其中,肝胰脏是消化酶的主要分泌器官,肠道则是营养物质的消化吸收场所[23]。也有研究表明,采食量可在一定程度上反映鱼类肠道对其营养物质的消化吸收能力[24]。本试验中,扣除Met 组的肝脏脂肪酶显著下降,同时,相对于扣除Cys,扣除Met 也明显降低了丝尾鳠幼鱼肝脏二糖酶活性,这与本试验中丝尾鳠的采食量趋势相一致。唐炳荣[23]等在对草鱼(Ctenopharyngodon idellus)的研究中发现,缺乏Met 会显著降低生长中期草鱼肝脏和肠道中的胰蛋白酶和脂肪酶活性;类似地,彭艳等[25]对建鲤(Cyprinus carpiovar Jian)的研究中也得出Met 对其肠道胰蛋白酶、脂肪酶均有显著性影响。肠道消化酶活性的高低可代表水生动物消化能力的强弱,各种水生动物的消化酶活性是不同的,鱼类的健康状况也会对消化酶有一定的影响。因此,本试验扣除Met 时,不利于丝尾鳠幼鱼对营养物质的消化吸收,从而导致其消化酶活性降低以及生长性能和饲料利用率的下降。
3.3 不同饲料处理对丝尾鳠蛋白质代谢的影响
Met 在肝脏中的代谢主要由3 个代谢途径的蛋氨酸-高半胱氨酸循环组成[26-27],分别为转甲基作用、再甲基作用和转巯基作用(见图1)。其中转甲基作用和再甲基作用构成了动物体内大多数的Met 循环,而转巯基作用则仅分布于部分器官中,包括肝脏、肠道和胰腺[28]。首先,Met 在MAT 的催化下生成S-腺苷蛋氨酸(SAM),SAM 不仅是重要的甲基供体,还在体内参与多胺合成,转甲基后生成腺苷高半胱氨酸,后者再进一步进行水解成为高半胱氨酸(Hcy)。然后Hcy 作为一种重要的的中间代谢产物,既可以通过MS 催化生成Met,也可以通过BHMT 催化的酶促反应进行再甲基化生成Met,或者Hcy 在CBS 的作用下发生不可逆反应生成胱硫醚[29]。由此可知,Met 在动物体内的代谢是一个复杂的生物化学反应体系,其中MAT、BHMT和MS主要与甲基化合成Met及其甲基供体的循环利用有关,而CBS则主要与Met 转巯基降解及其他含硫生物活性物质的合成有关。本试验结果表明,扣除Met 后均显著影响了丝尾鳠幼鱼肝脏中MAT、CBS、BHMT 和MS 的活性。由于Hcy 是MAT、BHMT、MS和CBS酶促反应的共同底物,所以Met 代谢的重点调控点是在甲基化和转巯基途径之间的分配。本研究结果显示,当扣除丝尾鳠幼鱼饲料中的Met时,其肝脏中MAT、CBS和BHMT的活性相对于其他饲料处理组均有上升的趋势,说明丝尾鳠幼鱼对饲料中极低Met 的利用主要是通过同时提高Hcy的甲基化和转巯基的反应来实现[30]。

图1 蛋氨酸部分代谢途径
研究表明,在动物代谢过程中很多氨基酸都是转化为谷氨酸来参与体内代谢的。因此,GDH是反应蛋白质分解代谢和NEAA合成的一种关键酶,它的活性高低在一定程度上反应了鱼类体内蛋白质分解代谢的情况。在动物体蛋白质代谢过程中转氨酶起着非常重要的作用,是机体中间代谢的关键酶反应。而转氨酶的种类有很多,其中以ALT和AST活性最强。本试验结果显示,相对于其他饲料处理组,扣除Met后丝尾鳠幼鱼肝脏中ALT和AST活性明显升高,这些结果均表明了肝脏中氨基酸代谢的不平衡状况[17]。类似地,AMPD 也是反映肝脏损伤程度的敏感指标,丝尾鳠幼鱼肝脏中的AMPD活性表现出与ALT、AST相同的变化趋势,进一步说明Met 的缺乏会影响丝尾鳠幼鱼的氨基酸代谢。
3.4 不同饲料处理对丝尾鳠抗氧化功能的影响
已有研究表明,动物代谢过程中会产生一定量自由基,生物体内的自由基处于不断产生与清除的动态平衡中,而自由基含量过高则会导致细胞结构和功能的破坏[31],抗氧化功能的强弱主要通过其抗氧化体系的抗氧化酶反映,T-AOC、POD、SOD、GSH-Px 与CAT是抗氧化体系内主要的抗氧化酶。T-AOC 是鱼类抗氧化能的总体表现,其活性大小在一定程度上反应了机体抗氧化能力的强弱,POD 可与SOD、CAT 相互协调配合,清除体内过剩的自由基[32]。类似地,GR 也可清除生物体内活性氧,还能催化氧化型谷胱甘肽与还原型谷胱甘肽两者间的相互转化,是产生谷胱甘肽(GSH)的关键酶,而GSH-Px则是机体内广泛存在的一种重要的过氧化物分解酶,主要是催化GSH参与过氧化反应,清除在细胞呼吸代谢过程中产生的过氧化物,在保护细胞免受自由基损伤过程中起着关键作用[33]。MDA是脂质过氧化的产物,其含量直接反映了脂质过氧化的速率和强度,间接反映了机体清除自由基的能力。本试验结果表明,当丝尾鳠幼鱼饲料中扣除Met后,其抗氧化功能呈现出整体下降的趋势,所以Met的缺乏会造成丝尾鳠幼鱼抗氧化功能的下降。
3.5 不同饲料处理对丝尾鳠鱼体组成的影响
本试验结果显示,当扣除Met后,鱼体水分含量显著升高;相反地,扣除Met 后鱼体粗蛋白、粗脂肪和粗灰分的含量则显著降低。研究表明,饲料中Met 不足时会使中华鳖[34]的粗脂肪含量下降,曾雯娉等[35]对凡纳滨虾的研究也发现其肌肉中的粗脂肪含量随Met含量的升高呈先升后降的趋势,这与本试验结果相类似。但Simmons等[36]在北极嘉鱼(Salvelinus alpinus)上的研究表明,随着饲料Met含量的升高,其灰分沉积呈相反的趋势,这与本试验结果不同。其原因可能是由于实验鱼的种类饲料的组成不同造成的。Cai 等[37]对斑点叉尾鮰的研究也表明,当饲料中Met含量较低时,会显著降低其鱼体粗蛋白含量,这与本试验结果相似。类似地,Goff 等[38]对红拟石首鱼(Sciaenops ocellatus)、陈乃松等[39]对大口黑鲈(Micropterus salmoides)等的研究结果同样显示,在Met含量不足时会明显降低鱼体粗蛋白含量。出现这一结果的原因可能是由于饲料中Met 不足时,使饲料中氨基酸的组成不平衡,进而导致鱼类机体的蛋白质合成功能下降,表现为鱼体粗蛋白含量的降低。
4 结论
蛋氨酸缺乏时显著降低了丝尾鳠幼鱼的生长性能,并对丝尾鳠幼鱼的消化酶活性、蛋白质代谢和抗氧化功能造成明显的负面影响。因此,蛋氨酸是丝尾鳠幼鱼的必需氨基酸。
