玉米和花生间作体系光合碳同化酶活性对CO2浓度升高的响应特征
2021-09-04薛华龙孙增光焦念元
李 雪,刘 娟,薛华龙,孙增光,王 飞,尹 飞,刘 领,焦念元*
(1 河南科技大学 农学院/河南省旱地农业工程技术研究中心,河南洛阳 471023;2 河南省农业科学院经济作物研究所,郑州 450002)
联合国政府间气候变化专门委员会(IPCC)第五次评估报告指出,如不加以控制则21世纪末全球大气CO2浓度将超过700 μmol·mol-1[1]。CO2是作物生长最重要的影响因子之一,其浓度升高可提前作物生育期、改善根冠比、增加生物量及产量[2],并且作为光合碳同化底物可直接影响作物气孔导度、蒸腾速率以及碳同化进程,进而影响光合作用[3]。目前全球正面临CO2浓度升高所带来的农业生产挑战,研究作物光合碳同化酶活性对CO2浓度升高的响应特点,对应对全球气候变化,确保粮食安全至关重要。
已有研究表明,短时间的供应CO2可以促进单作C4作物丙酮酸磷酸双激酶(PPDK)基因的表达及活性[4],增加其光合速率;C3植物叶片气孔导度会随着CO2浓度升高而下降,对CO2响应的敏感度随着温度增高而增加,在高CO2浓度下,Rubisco酶活性得到促进,光合最适温度提高,进而提高叶片光合速率[5]。还有研究表明CO2浓度升高能提高单作黄瓜叶片抗氧化酶活性以及叶绿素含量,增加“午休”时段的光合速率,显著缓解高温对光合作用的胁迫伤害[6];在干旱条件下,高CO2浓度还可以改善由干旱胁迫引起的氧化损伤,提高植物的抗逆能力[7]。高CO2浓度环境使单作水稻和玉米的叶绿素含量提高,净光合速率增加,但气孔导度及蒸腾速率下降[8]。叶子飘等认为大豆叶片的净光合速率以及电子利用效率随着CO2浓度增高而增大,但不同浓度下羧化效率差异并不显著[9-10]。而惠俊爱等认为CO2浓度增高促进了玉米淀粉积累,但Rubisco酶活性变化不显著,且叶片中叶绿素含量下降[11]。但也有研究表明,当长时间给单作作物供应CO2时会导致光适应现象,即由CO2带来的促进效用消失[12],这可能与CO2浓度升高导致作物碳的源库失衡有关[13]。而在玉米、花生间作复合体系中,CO2浓度升高能改善复合群体冠层结构,提高玉米对强光、花生对弱光的利用,实现光能的分层立体利用[14],进一步改善间作玉米、花生的光合生理特性[15-16]。那么,升高CO2浓度对间作玉米、花生的光合碳同化酶活性产生了哪些影响?
磷素作为光合磷酸化底物,产生光合碳同化所需要的能量ATP[17],施用磷肥能促进作物光合磷酸化生产ATP[18],提高单作玉米净光合速率、叶绿素含量[19]、蒸腾速率和气孔导度[20]。同时,施磷还能在一定程度上缓解由干旱造成的气孔导度降低,提高单作玉米水分利用效率[21]。缺磷使作物叶片光合性能减弱,导致减产[21],而CO2浓度升高能够有效弥补缺磷带来的减产效应[22]。在间作体系中,于CO2浓度升高条件下,磷肥对间作玉米和间作大豆的生长优势有显著促进作用[23],改善间作玉米的光合特性[15]。施磷能显著提高间作玉米和花生的叶绿素含量,增强其功能叶对光的捕获及转化能力[24-25]。那么,CO2浓度升高条件下,磷肥对间作玉米、花生的光合碳同化酶活性及净光合速率有哪些调控效应?为了探究以上问题,在升高CO2浓度条件下,本研究考察了间作玉米、花生功能叶的净光合速率和Rubisco、Ru5PK等光合碳同化关键酶活性的变化特点,从酶水平揭示间作玉米、花生功能叶光合碳同化特点对CO2浓度升高的响应机理,为将来CO2浓度升高环境下间作玉米、花生绿色高产高效栽培提供理论基础。
1 材料和方法
1.1 试验地概况
试验于2018-2019年在河南科技大学试验农场(33°35′N,111°8′E)进行。试验点地处温带,属于半湿润、半干旱大陆性季风气候,年平均气温12.1~14.6 ℃,年平均降雨量约610 mm,年平均蒸发量约2 116 mm,年日照时2 300~2 600 h,无霜期216~223 d,年平均辐射量约492 kJ·cm-2。试验地土壤为黄潮土,质地为中壤;0~20 cm土层土壤理化性质分别为:土壤容重1.35 g·cm-3、碱解氮33.96 mg·kg-1、速效磷6.84 mg·kg-1、有机质10.74 g·kg-1和pH 7.65。
1.2 试验设计
供试材料为玉米杂交种‘豫单9953’,花生品种‘花育16’,以玉米、花生间作2∶4模式为研究对象。设置CO2浓度和施磷量双因素完全随机区组试验,CO2浓度设环境CO2浓度(390±50 μmol·mol-1,Ca)和升高CO2浓度 (700±50 μmol·mol-1, Ce)2个水平,施磷量设不施磷 (P0)和施磷 (P180,180 kg P2O5·hm-2) 2个水平,共组成4个处理组合CaP0、CeP0、CaP180和CeP180,均在开顶式气室(open-top-chamber,OTC)内进行。玉米、花生间作采用2∶4 (2行玉米间作4行花生)模式,玉米宽窄行种植,宽行行距160 cm,窄行行距40 cm,株距20 cm,花生播种于宽行之中,行距30 cm,株距20 cm,玉米花生之间行距35 cm,间作带宽2 m。南北向种植,每个小区宽6 m,长8 m,包含3个带宽,每个处理重复3次。每个小区均基施90 kg N·hm-2(肥源为尿素),再于玉米大喇叭口期追施90 kg N·hm-2;施磷处理每个小区均基施180 kg P2O5·hm-2(肥源为磷酸二氢铵),其他管理同大田生产。各处理在2018年于6月4日播种、9月16~22日收获,在2019年于6月18日播种、9月28日~10月9日收获。
CO2浓度升高采用开顶式气室(open-top-chamber,OTC),以罐装液态CO2为CO2气源。试验期间,采用德国LOE公司生产的减压阀控制CO2气室内CO2浓度在(700±50) μmol·mol-1。OTC为长方体钢架结构,宽6 m,长8 m,高2.5 m,面积48 m2,在玉米苗期开始供CO2,为减少内部气体外溢,上部框架边缘增加0.5 m宽45度收缩口,安装电风扇确保气室内空气流动、CO2浓度均匀。室壁采用阳光板,透光率达90%以上。
1.3 测定项目与方法
1.3.1 功能叶净光合速率2018年苗后62 d和2019年苗后37、58、68和95 d使用LI-6400XT型光合仪(LI-COR, 美国),选择晴天在9:30~12:00自然光强下测量功能叶净光合速率。测量时均测定方向相同的玉米穗位叶以及花生主茎倒三叶,每个小区重复3次。
1.3.2 功能叶中光合碳同化酶活性2018年玉米、花生苗后43、59和72 d时取玉米穗位叶、花生主茎倒三叶,置于液氮中暂时保存,后转移至-80 ℃冰箱保存。在低温环境下取玉米、花生功能叶样0.25 g,重复3次,用植物组织破碎仪将其打碎,加入1.5 mL经过预冷处理的Tris-HCl提取缓冲液(用50 mL的0.1 mol·L-1Tris和21.9 mL的0.1 mmol·L-1HCl,配置成pH 8.2提取缓冲液),震荡均匀后将其放入冷冻离心机中在4 ℃、15 000 r/min离心20 min,上清液即为待测的酶提取液,用于1, 5-二磷酸核酮糖羧化酶(Rubisco)、磷酸烯醇式丙酮酸羧化酶(PEPC)、磷酸丙酮酸双激酶(PPDK)、3-磷酸甘油醛脱氢酶(GAPDH)、NADP-苹果酸脱氢酶(NADP-MDH)、5-磷酸核酮糖激酶(Ru5PK)、果糖-1, 6-二磷酸酯酶(FBPase)活性的测定。酶活性测定采用酶联免疫法(ELISA),按酶联免疫试剂盒(上海酶联生物科技有限公司)说明进行反应,用酶标仪(Thermo Multiskan GO,美国)在450 nm波长下测定吸光度值计算酶活性(U·g-1)。
1.3.3 籽粒产量在收获期,于各小区选取代表性2 m双行的玉米果穗、花生植株,风干后称取玉米籽粒、花生荚果质量,并计算其产量(kg·hm-2)。
1.4 数据处理与分析
采用Excel 2016和SPSS 22.0软件对数据进行整理、统计分析与作图,采用Duncan法进行显著性检验及方差分析。
2 结果与分析
2.1 CO2浓度升高和施磷对间作体系中玉米功能叶光合碳同化酶活性的影响
2.1.1 叶肉细胞内光合碳同化酶活性叶肉细胞内的光合碳同化酶主要包括磷酸烯醇式丙酮酸羧化酶(PEPC)、磷酸丙酮酸双激酶(PPDK)和NADP-苹果酸脱氢酶(NADP-MDH),在C4作物光合碳同化过程中起到CO2初次羧化固定的作用。图1显示,间作体系中玉米功能叶叶肉细胞内光合碳同化酶PEPC、PPDK和NADP-MDH活性在各处理下的变化趋势相似。随着出苗后天数的增加,各处理组合各酶活性均先升后降,并均在苗后59 d时最高;在同一时期,各酶活性在同一施磷水平下均表现为CO2浓度升高处理(Ce)不同程度高于环境CO2浓度处理(Ca),在同一CO2浓度水平下大多表现为施磷处理(P180)不同程度高于不施磷处理(P0)。
其中,Ce玉米功能叶PEPC活性在P180水平下比同期Ca提高了14.3%~34.8%,在P0水平下比同期Ca提高了25.0%~30.0%,且均在苗后43 d和59 d时增幅达到显著水平(P< 0.05),均在72 d时增幅不显著(P> 0.05);P180处理玉米功能叶PEPC活性在Ca、Ce水平下分别比P0处理提高了15.0%~20.0%和6.7%~19.2%,但增幅仅分别在苗后43 d和59 d时达到显著水平(图1,A)。
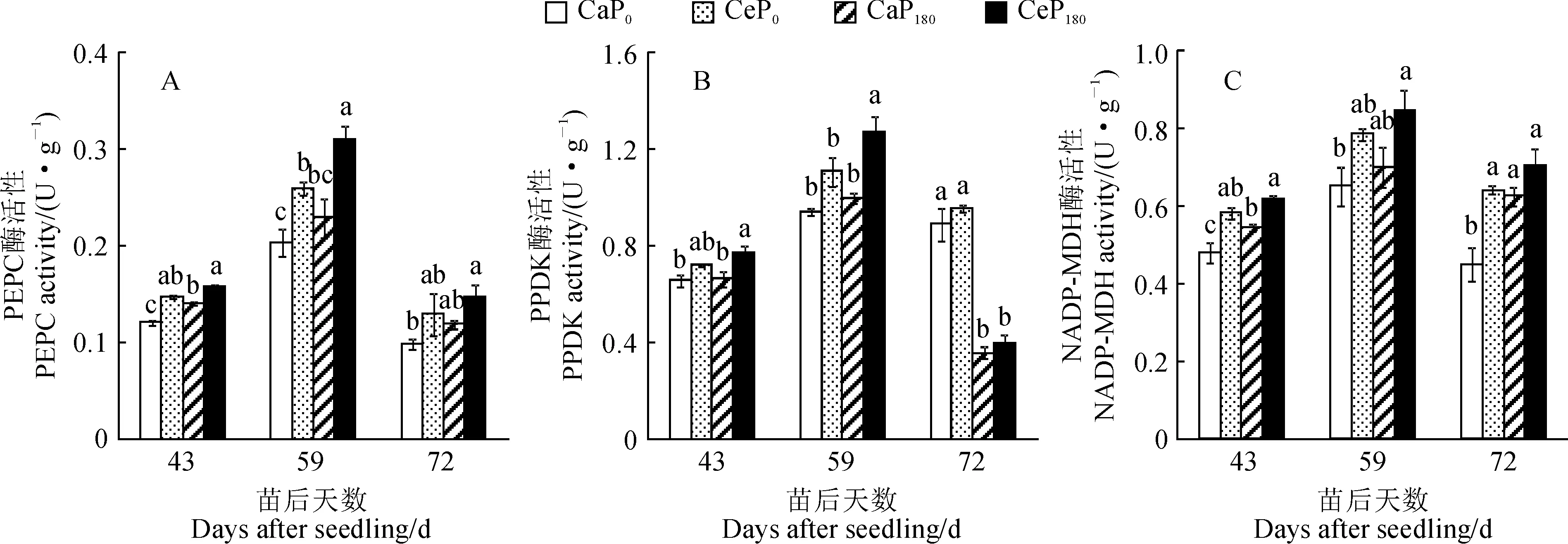
P0. 0 kg P2O5·hm-2;P180. 180 kg P2O5·hm-2;Ca.环境CO2浓度(390±50 μmol·mol-1);Ce.增加CO2浓度 (700±50 μmol·mol-1);PEPC.磷酸烯醇式丙酮酸羧化酶;PPDK.磷酸丙酮酸双激酶;NADP-MDH.NADP-苹果酸脱氢酶; 不同小写字母表示同一时期处理间在0.05水平差异显著;下同图1 CO2浓度升高和施磷对间作玉米功能叶PEPC、PPDK和NADP-MDH活性的影响P0. 0 kg P2O5·hm-2; P180.180 kg P2O5·hm-2; Ca. Ambient CO2 concentration(390±50 μmol·mol-1); Ce. Elevated CO2concentration (700±50 μmol·mol-1). PEPC. Phosphoenolpyruvate carboxylase; PPDK. Phosphopyruvate dikinase; NADP-MDH. NADP-malate dehydrogenase; Different lowercase letters mean significant difference among treatments during the same stage at 0.05 level. The same as belowFig.1 Effects of elevated CO2 concentration and phosphorus application on activities of PEPC, PPDK, and NADP-MDH in functional leaves of intercropping maize
同时,Ce 玉米功能叶的PPDK活性在P180水平下比Ca提高了11.1%~27.0%,并在苗后43和59 d达到显著水平,在P0水平下比Ca提高了7.9%~18.1%,但均未达到显著水平。在苗后43和59 d,P180处理玉米功能叶PPDK活性在Ca 和Ce水平下分别比P0处理提高了1.5%~6.4% 和6.9%~14.4%,而仅在苗后59 d的Ce水平下达到显著水平。但在苗后72 d,玉米功能叶PPDK活性施磷处理反而呈显著下降趋势(图1,B)。说明在间作玉米生育前期 CO2浓度升高以及施磷对PPDK活性都具有正调控效应,而随着生育进程施磷带来的正调控效应逐渐削弱至消失。
另外,Ce处理间作玉米功能叶NADP-MDH活性在P180水平下比Ca处理提高了12.7%~21.4%,但仅在苗后43 d达到显著水平,在P0水平下比Ca处理提高了20.8%~42.2%,在苗后43和72 d均达到显著水平;在同一CO2浓度下,玉米功能叶NADP-MDH活性均表现为P180显著大于 P0(P< 0.05),在Ca和Ce水平下分别提高了7.7%~40%和6.9%~10.9%,但仅在苗后43和72 d的Ca水平下达到显著水平(图1,C)。这表明CO2浓度升高可增加间作玉米功能叶叶肉细胞内碳同化酶活性,促进对CO2的初次羧化,施磷在生育前中期对此有正调控作用。
2.1.2 维管束鞘细胞内光合碳同化酶活性维管束鞘细胞内的光合碳同化酶主要包括1, 5-二磷酸核酮糖羧化酶(Rubisco)、3-磷酸甘油醛脱氢酶(GAPDH)和5-磷酸核酮糖激酶(Ru5PK),其功能是对CO2进行二次羧化固定,进行卡尔文循环,完成后续碳同化过程。间作玉米功能叶维管束鞘细胞内光合碳同化酶Rubisco、GAPDH和Ru5PK活性随生育时期和处理的变化趋势与其叶肉细胞内光合碳同化酶相似,即均在苗后59 d时达到最大值,Ce处理均大体高于Ca处理,P180处理不同程度高于P0处理(图2)。
其中,玉米功能叶Rubisco活性于苗后43和72 d时在各处理组合间无显著差异;在苗后59 d时,Ce处理玉米功能叶Rubisco活性在P0和P180水平下分别比Ca提高了21.1%和37.1%,但仅后者达到差异显著水平(P< 0.05),此时P180处理玉米功能叶Rubisco活性在Ca和Ce水平下分别比P0处理增加18.2%和4.4%,但均未达到显著水平(图2,A)。同时,间作玉米功能叶的GAPDH活性于苗后43 d时在各处理间差异不显著;在苗后59 和72 d时,Ce处理GAPDH活性在P0和P180水平下分别比Ca显著提高30.6%~45.0%和26.5%~37.2%,P180处理GAPDH活性在Ce和Ca水平下分别比P0处理提高了4.9%~9.7%和4.9%~20.3%,但仅后者达到显著水平(图2,B)。另外,间作玉米功能叶Ru5PK活性于苗后43 d时在各处理间差异也不显著,但表现为Ce> Ca;在苗后59 和72 d时,Ce处理Ru5PK活性在P0和P180水平下分别比Ca显著提高10.6%~29.7%和15.3%~28.6%,P180处理Ru5PK活性在Ce和Ca水平下分别比P0处理增加6.6%~24.4%和7.5%~19.3%(图2,C)。以上结果表明CO2浓度增高能促进间作玉米功能叶维管束鞘细胞对CO2的二次羧化固定,施磷对其具有正调控作用。
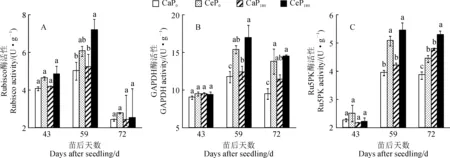
Rubisco.1,5-二磷酸核酮糖羧化酶;GAPDH.3-磷酸甘油醛脱氢酶;Ru5PK.5-磷酸核酮糖激酶。下同图2 CO2浓度升高和施磷对间作玉米功能叶维管束鞘细胞内光合碳同化酶活性的影响Rubisco. 1, 5-ribulose bisphosphate carboxylase; GAPDH. 3-glyceraldehyde phosphate dehydrogenase; Ru5PK. Ribulokinase 5-phosphate. The same as belowFig.2 Effects of elevated CO2 concentration and phosphorus application on photosynthetic carbon assimilase activity in the tube bundle sheath cells of intercropped maize functional leaves
2.2 CO2浓度升高和施磷对间作体系中花生功能叶光合碳同化关键酶活性的影响
表1显示,间作体系中花生功能叶的Rubisco、GAPDH、Ru5PK、果糖-1, 6-二磷酸酯酶(FBPase)活性在各处理下的变化趋势相似。即随着苗后天数的增加,各处理组合功能叶各酶活性均先升后降,并均在苗后59 d时最高,Ce处理均大体高于Ca处理,P180处理不同程度高于P0处理。其中,花生功能叶Rubisco活性于苗后72 d时在各处理组合间差异不显著;在苗后43和59 d时,Ce处理Rubisco活性在P0和P180水平下分别比Ca显著提高13.9%~39.4%和13.4%~40.9%,P180处理Rubisco活性在Ce和Ca水平下分别比P0处理提高了11.9%~30%和12%~13.9%,但仅后者达到显著水平。同时,间作花生功能叶的GAPDH活性各处理间均存在显著差异;Ce处理GAPDH活性在P0和P180水平下分别比Ca显著提高26.5%~56.3%和17.7%~51.8%,P180处理GAPDH活性在Ce和Ca水平下分别比P0处理提高5.2%~9.5%和0.3%~33.9%。另外,间作花生功能叶的Ru5PK活性各处理间也存在显著差异。Ce处理Ru5PK活性在P0和P180水平下分别比Ca显著提高17.8%~51.8%和19.5%~61.4%,P180处理Ru5PK活性在Ce和Ca水平下分别比P0处理提高13.8%~29.7%和8.1%~43.9%,但仅后者达到显著水平。此外,间作花生功能叶的FBPase活性在各处理间叶存在显著差异,并表现为Ce>Ca、P180>P0;Ce处理FBPase活性在P0和P180水平下分别比Ca显著提高31.5%~43.4%和22.0%~74.2%,P180处理FBPase活性在Ce和Ca水平下分别比P0处理提高15.5%~62.5%和24.4%~30.9%。以上结果表明CO2浓度增高能促进花生功能叶暗反应中对CO2羧化固定,施磷对其有着正调控作用。
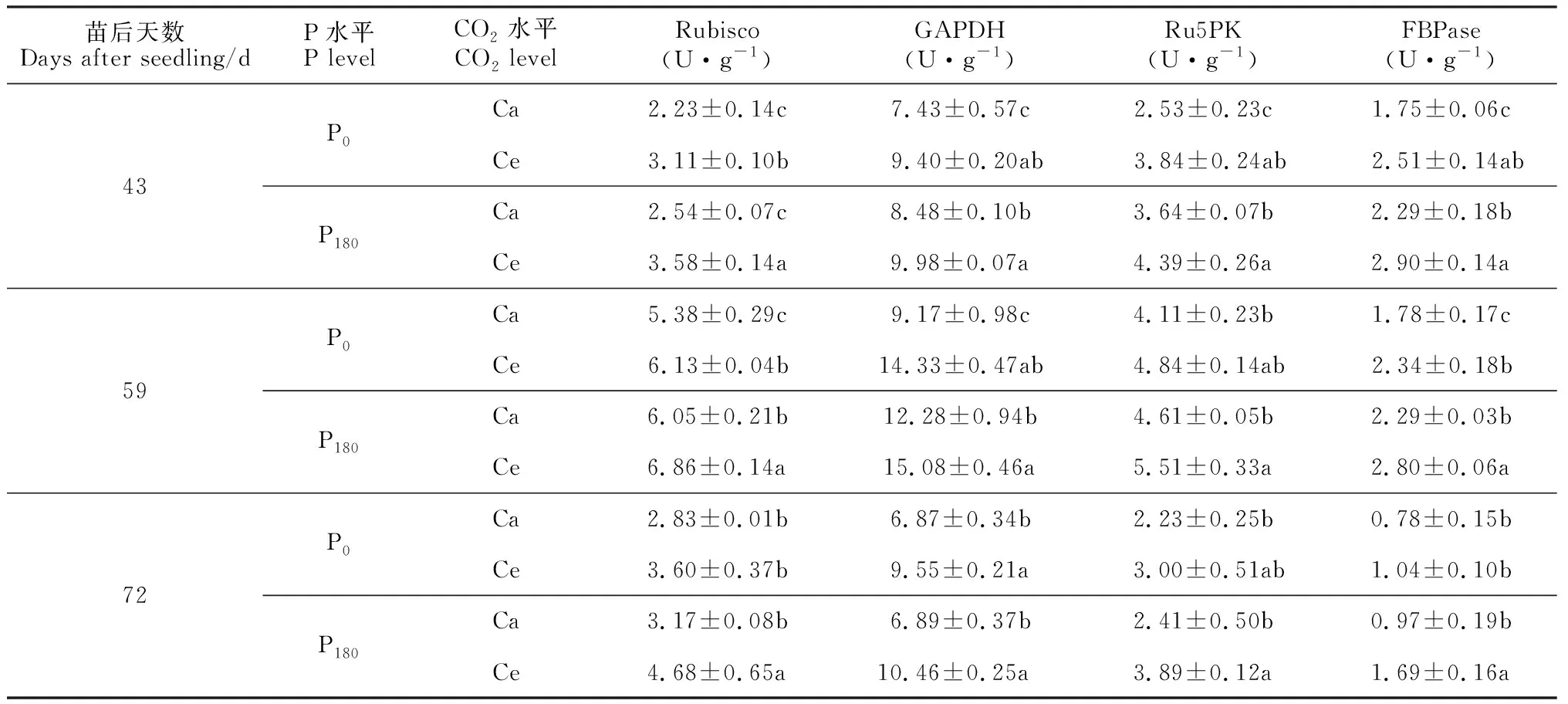
表1 CO2浓度升高对间作花生功能叶光合碳同化酶活性的影响
2.3 CO2浓度升高和施磷对间作体系中玉米和花生功能叶净光合速率的影响
图3显示,间作体系中玉米和花生功能叶净光合速率在各处理下的变化趋势相似,仍表现为Ce处理均大体高于Ca处理,施磷处理(P180)不同程度高于不施磷处理(P0)。
首先,在2018年,与Ca相比,二氧化碳浓度升高(Ce)和施磷(P180)均显著提高了间作玉米苗后62 d的穗位叶净光合速率(P< 0.05),并表现为CeP180>CaP180>CeP0>CaP0(图3,A)。在2019年,同一磷水平下,Ce处理间作玉米穗位叶净光合速率在苗后37、58和68 d均显著高于 Ca,增幅分别为14.8%~18.8%(P0)和18.1%~18.4%(P180),在苗后95 d时则比Ca降低了6.4%~8.0%,但仅在P180处理下达到显著水平;在同一CO2浓度下,P180处理间作玉米功能叶净光合速率均高于P0处理,增幅在Ca和Ce处理下分别达到8.8%~28.2%和8.5%~26.1%。
其次,2018年苗后50 d和70 d时,二氧化碳浓度升高(Ce)和施磷(P180)均显著提高了花生功能叶净光合速率(图3,B)。在2019年,同一磷水平下,花生功能叶净光合速率在苗后30 d、50 d和70 d均表现为Ce 显著高于 Ca,增幅分别为24.2%~32.3%(P0)和8.1%~14.4%(P180);在苗后95 d时则表现为Ce低于Ca,降幅为5.8%~15.3%;在同一CO2浓度下,间作花生功能叶净光合速率各时期均表现为P180高于 P0,Ca和Ce的增幅分别为15.0%~21.7%和11.6%~22.4%,且大多达到显著水平。可见,CO2浓度升高对间作花生功能叶净光合速率有显著的促进作用,施磷对其有正向调控作用。
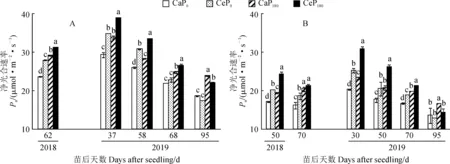
图3 CO2浓度升高对间作玉米(A)和间作花生(B)功能叶净光合速率的影响Fig.3 Effects of elevated CO2 concentration on net photosynthetic rate in functional leaves of intercropped maize (A) and intercropped peanut (B)
2.4 CO2浓度升高和施磷对间作体系中玉米和花生籽粒产量的影响
Ce处理都不同程度提高了间作玉米、间作花生和间作体系的籽粒产量,且双因素方差分析结果显示,施磷和升高CO2浓度对间作玉米、间作花生和间作体系产量有显著影响,磷水平与CO2浓度对间作玉米和间作体系产量具有显著的正向交互作用(表2)。其中,Ce处理下间作玉米产量在P0和P180水平下分别比Ca显著提高4.4%和52.0%,施磷处理间作玉米产量在Ce和Ca水平下分别比不施磷处理提高24.2%和67.2%。同时,Ce处理间作花生产量在P0和P180水平下比Ca处理分别提高了10.3%和24.0%,施磷处理下间作花生产量比不施磷处理提高了17.1%~19.3%。此外,Ce处理间作体系产量相比Ca提高了5.7%~47.0%,施磷处理下间作体系的产量与不施磷相比分别提高了23.3%~58.4%,且均达到显著差异(P< 0.05)。这说明CO2浓度升高以及施磷均能促进间作玉米、间作花生和间作体系产量。另外,相关分析结果显示,玉米、花生间作体系籽粒产量与Rubisco等碳同化关键酶活性和净光合速率均呈显著性正相关(表3,表4),即CO2浓度升高能显著提高碳同化关键酶活性,促进间作玉米、间作花生的净光合速率,显著提高了其玉米、花生间作体系的产量。
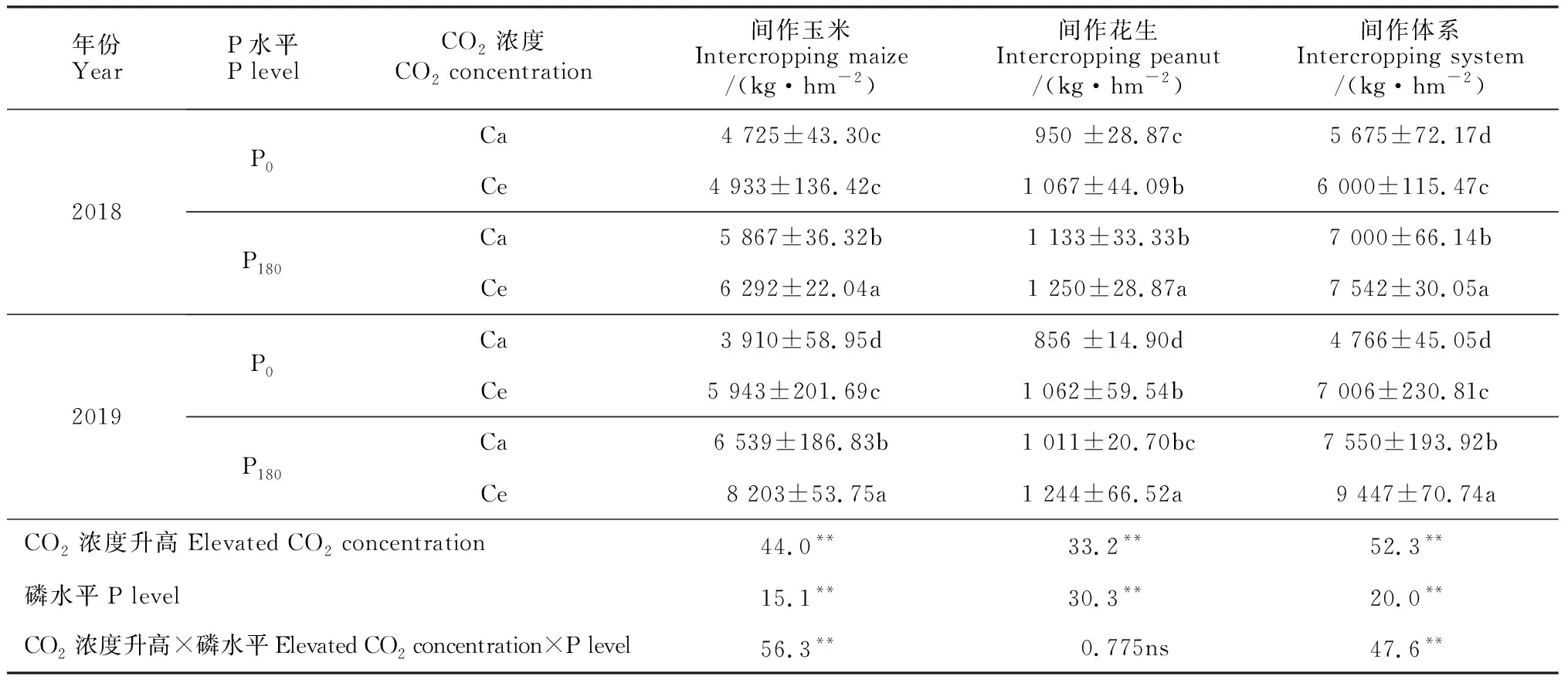
表2 CO2浓度升高对玉米、花生间作体系籽粒产量的影响
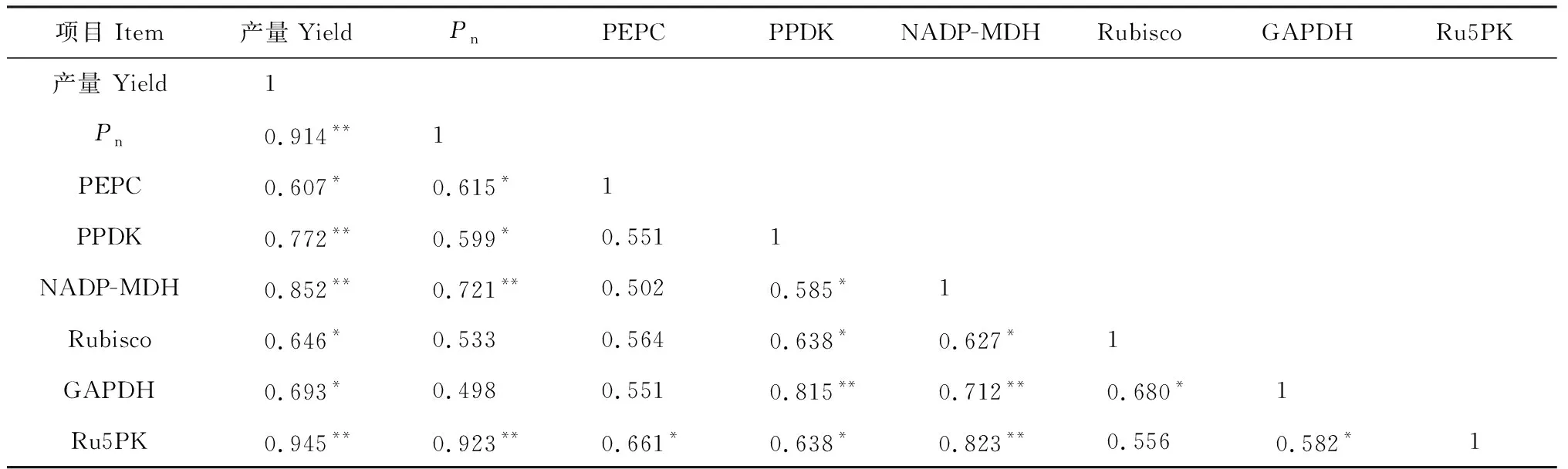
表3 间作玉米的产量、净光合速率和光合碳同化酶活性间相关性分析
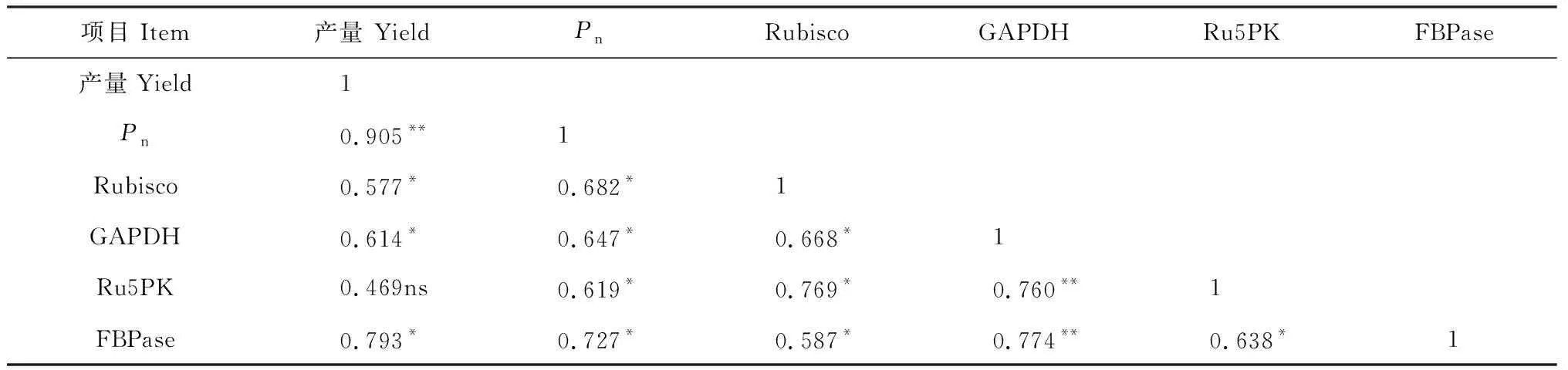
表4 间作花生产量、净光合速率和光合碳同化酶活性间相关性分析
3 讨 论
3.1 CO2浓度升高能提高间作体系中玉米和花生的光合碳同化酶活性
光合碳同化过程中,CO2作为底物,其浓度对作物光合有一定的影响。CO2浓度升高能使水稻和玉米的叶绿素值增高,净光合速率增加[8],玉米叶面积指数增大[26]。有研究表明Rubisco,PPDK,FBPase等均为光敏感酶[17, 27],受光调节,而王飞等认为CO2浓度升高能进一步提高间作玉米对强光、间作花生对弱光的利用[16]。本研究结果表明CO2浓度升高能增加间作玉米功能叶PEPC、PPDK和NADP-MDH酶活性,这说明CO2浓度升高能提高间作玉米叶肉细胞内对光合底物的初步羧化固定能力;而在CO2浓度升高条件下,Rubisco、GAPDH和Ru5PK等3个酶活性显著增加,说明CO2浓度升高促进了间作玉米维管束鞘细胞中对CO2的二次羧化固定。通过对间作玉米净光合速率与光合碳同化关键酶活性的相关性分析可知,净光合速率与PEPC、PPDK、 NADP-MDH和Ru5PK等酶活性呈显著正相关。CO2浓度升高能进一步增强间作玉米对强光的利用,提高净光合速率,是由于其PEPC、NADP-MDH、PPDK、Rubisco、GAPDH和Ru5PK等光合碳同化关键酶活性的增加。叶子飘等认为大豆叶片的净光合速率以及电子利用效率随着CO2浓度增高而增大,但不同浓度下羧化效率差异并不显著[9]。周宁等认为高CO2浓度条件能提高水稻的净光合速率和实际光量子产量[28]。周晓旭研究表明冬小麦的光能转换率随CO2浓度升高而增加[29]。本研究表明CO2浓度升高能提高间作花生的Rubisco、GAPDH、FBPase和Ru5PK酶活性,这说明CO2浓度升高能促进间作花生功能叶Rubisco对CO2的羧化,1,3-PGA向3-磷酸甘油醛的转化,果糖-6-磷酸(F-6-P)的生成,及Ru5PK催化RUBP的再生,加速碳同化。通过相关性分析可知间作花生净光合速率与Rubisco、GAPDH、FBPase和Ru5PK等碳同化关键酶活性显著相关,所以CO2浓度升高能进一步提高间作花生对弱光的利用,提高净光合速率,关键在于间作花生功能叶的Rubisco、GAPDH、FBPase和Ru5PK等碳同化关键酶活性的提高。
3.2 CO2浓度升高条件下,施磷对间作玉米和花生光合碳同化酶活性具有明显正调控效应
磷素是植物生长发育所必需的元素之一,在光合磷酸化过程中起着至关重要的作用,碳同化反应过程依赖于光合磷酸化产生的能量而进行[30-31],磷肥的施用可促进植株的光合磷酸化反应[18, 32-33],进而促进碳同化过程。徐明睿等认为施磷可提高玉米叶面积指数以及SPAD值,改善叶片的光合性能[34]。本研究结果表明,CO2浓度升高后,增施磷肥比不施磷能提高间作玉米功能叶的PEPC、PPDK、NADP-MDH、Rubisco、GAPDH和Ru5PK和间作花生Rubisco、GAPDH、FBPase和Ru5PK等光合碳同化关键酶活性,这可能是因为施磷能促进光合磷酸化产生更多ATP,为光合碳同化提供更多的同化力,进而激发光合碳同化关键酶活性,增强玉米、花生间作功能叶对CO2的羧化固定。因此,在CO2浓度升高条件下,施磷可显著提高玉米、花生间作功能叶的净光合速率,从而促进光合作用。
3.3 CO2浓度升高和施磷可以提高间作玉米和花生的籽粒产量
CO2浓度升高对单作玉米的干物质积累及产量有促进作用[35]。同时有研究认为在高CO2浓度环境下小麦、玉米产量显著提高,与氮肥具有显著的互作效应[36, 37]。通过相关分析结果显示产量与Rubisco等碳同化关键酶活性和净光合速率均呈显著性正相关,即CO2浓度升高能显著提高碳同化关键酶活性,促进间作玉米、间作花生的净光合速率,显著提高了其玉米、花生间作的产量,这可能与CO2浓度升缩短间作玉米营养生长期,延长籽粒灌浆期,增加穗粒数和粒重有密切关系[16]。有研究认为施磷可以显著提高玉米净光合速率、促进作物光合磷酸化生产ATP[18],本研究结果表明,增施磷肥可促进玉米花生间作体系的光合作用,促进其生长发育,进而提高产量。施磷提高单作施磷双因素结果表明CO2浓度升高与磷肥有明显的正互作效应,这说明CO2浓度升高和施磷对玉米、花生间作产量具有正向协调作用。
4 结 论
CO2浓度升高通过提高间作玉米功能叶的PEPC、PPDK、NADP-MDH、Rubisco、GAPDH和Ru5PK和间作花生Rubisco、GAPDH、FBPase和Ru5PK等光合碳同化关键酶活性,促进光合碳同化,提高净光合速率,增加间作体系的产量;在CO2浓度升高条件下,施磷对玉米、花生间作光合碳同化关键酶活性、净光合速率及产量均具有正调控作用。
