隆林山羊CIDEc基因的克隆及组织表达分析
2021-08-03邹菊红江雨航张叁保高小童申玉建韦英明黄大安黄艳娜蒋钦杨
邹菊红 江雨航 张叁保 高小童 申玉建 韦英明 黄大安 黄艳娜 蒋钦杨

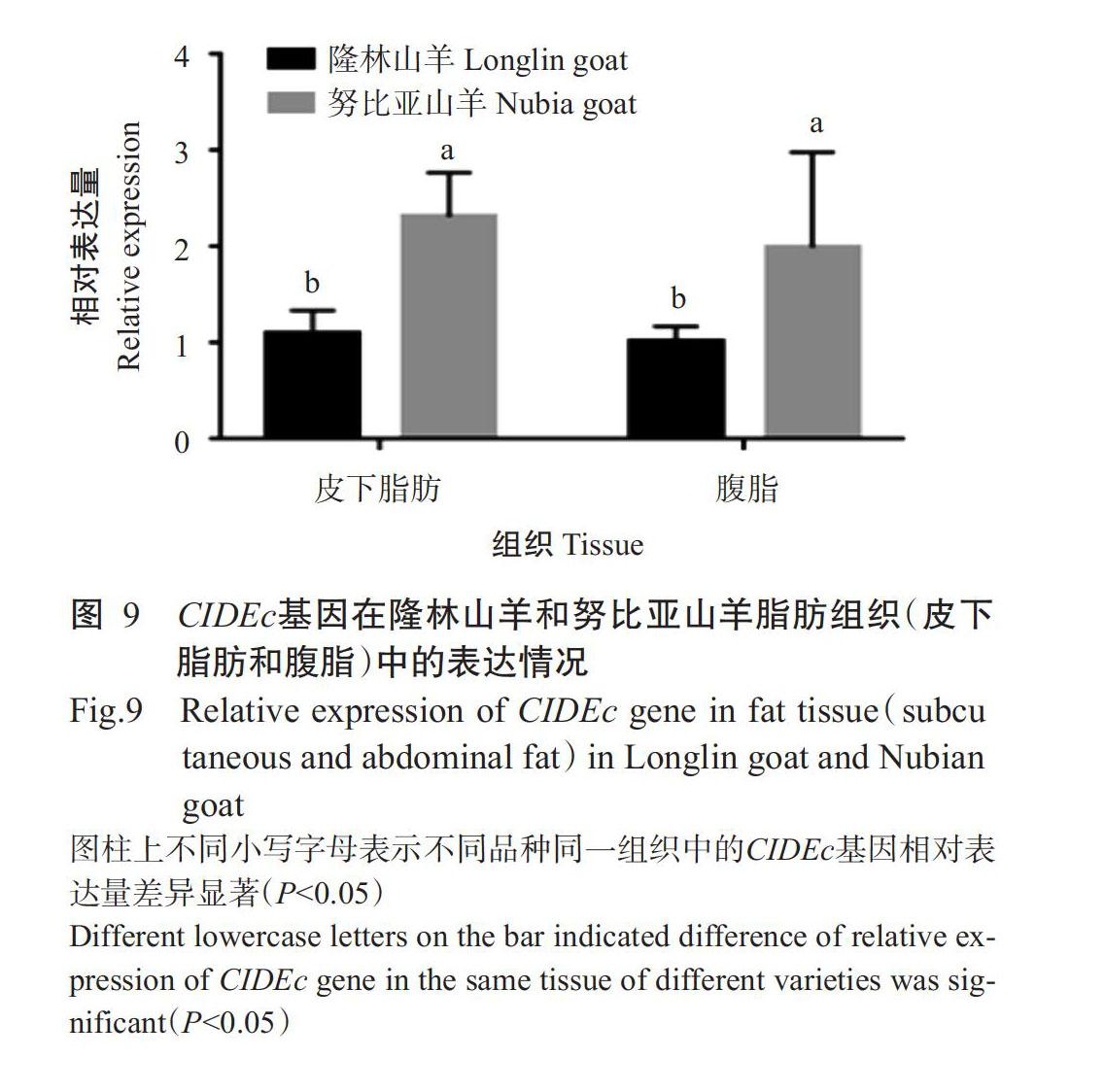
摘要:【目的】明確隆林山羊诱导细胞凋亡DFF45样效应因子c基因(CIDEc)的生物学特性及其表达规律,为揭示CIDEc基因对山羊脂肪代谢的调控机制提供理论依据。【方法】提取隆林山羊背最长肌、心脏、肝脏、脾脏、肾脏、腹脂和皮下脂肪及努比亚山羊腹脂和皮下脂肪的总RNA,PCR扩增隆林山羊CIDEc基因编码区(CDS)序列,使用ExPASy、TMHMM Server v.2.0、ProtScale、NPS@SOPMA和SWISS-MODEL等在线软件进行生物信息学分析,并利用实时荧光定量PCR检测CIDEc基因在隆林山羊和努比亚山羊不同组织器官中的表达情况。【结果】隆林山羊CIDEc基因CDS序列全长为741 bp,共编码244个氨基酸残基,其编码蛋白分子量26.09 kD,不稳定系数48.44,脂肪指数100.7,理论等电点(pI)5.28,属于酸性蛋白,不存在跨膜结构,亲水性较强。隆林山羊CIDEc基因CDS序列与NCBI已公布的山羊CIDEc基因(XM_018038446.1)CDS序列相对比仅有1处碱基发生突变,即第560位点G突变为T,属于同义突变。隆林山羊与绵羊的CIDEc基因CDS序列相似性最高(99.0%),与小鼠的相似性较低(79.6%);基于CIDEc基因CDS序列相似性构建的系统发育进化树也显示隆林山羊与绵羊的亲缘关系最近,与小鼠的亲缘关系较远。在隆林山羊CIDEc蛋白的二级结构中:α-螺旋占32.38%,β-转角占12.70%,延伸链占13.11%,无规则卷曲占41.80%。CIDEc基因在隆林山羊7个组织器官中均有表达,且在腹脂和皮下脂肪中高表达,极显著高于在其他器官组织的相对表达量(P<0.01);CIDEc基因在努比亚山羊脂肪组织(皮下脂肪和腹脂)中的相对表达量显著高于在隆林山羊脂肪组织中的相对表达量(P<0.05)。【结论】CIDEc基因在隆林山羊各器官组织中均有表达,以在脂肪组织中的表达水平较高,且其在努比亚山羊脂肪组织(皮下脂肪和腹脂)中的表达水平显著高于在隆林山羊中的表达水平。可见,CIDEc蛋白是脂肪代谢的重要调节剂,与动物体内脂质存储密切相关。
关键词: 隆林山羊;CIDEc基因;克隆;生物信息学分析;脂肪代谢;调控机制
中图分类号: S826.89 文献标志码: A 文章编号:2095-1191(2021)04-1082-08
Cloning and tissue expression analysis of CIDEc gene
in Longlin goat
ZOU Ju-hong1, JIANG Yu-hang1, ZHANG San-bao1, GAO Xiao-tong1, SHEN Yu-jian1,
WEI Ying-ming2, HUANG Da-an3, HUANG Yan-nan1, JIANG Qin-yang1*
(1College of Animal Science and Technology, Guangxi University, Nanning 530004, China; 2Agriculture and Animal Husbandry Development Institute, Guangxi University, Nanning 530004, China; 3Campus Cooperation
Office, Guangxi University, Nanning 530004, China)
Abstract:【Objective】To investigate the biological characteristics and expression pattern of DFF45-like effector C gene(CIDEc) in Longlin goat, and to provide theoretical basis for revealing the regulation mechanism of CIDEc gene on goat fat. 【Method】Total RNA was extracted from longissimus dorsi muscle, heart, liver, spleen, kidney, abdominal fat and subcutaneous fat of Longlin goat, and abdominal fat and subcutaneous fat of Nubia goat. The sequence of CIDEc gene coding region(CDS) of Longlin goats was amplified by PCR. Online softwares such as ExPASy, TMHMM Server v.2.0, ProtScale, NPS@SOPMA and SSWISS-Model were used for bioinformatics analysis, and real-time fluorescence quantitative PCR was used to detect the expression of CIDEc gene in different tissues and organs of Longlin goat and Nubia goat. 【Result】The CDS sequence of Longlin goat CIDEc gene was 741 bp, encoding 244 amino acid residues. The molecular weight of the encoded protein was 26.09 kD, the instability coefficient was 48.44, the fat index was 100.7, and the theoretical isoelectric point(pI) was 5.28. It was an acidic protein with no transmembrane structure and strong hydrophilicity. The CDS sequence of Longlin goat CIDEc gene had only one mutation compared with the CDS sequence of common goat CIDEc gene(XM_018038444.6) published by NCBI, that was, the mutation at site 560 G was changed to T, which belonged to the synonym mutation. The CDS sequence of CIDEc gene of Longlin goat was the highest in similarity with that of sheep(99.0%), and the lowest in similarity with that of mice(79.6%). The phylogenetic tree constructed based on the similarity of CIDEc gene CDS sequence also showed that Longlin goat was the most closely related to sheep, and closely related to mice. In the secondary structure of Longlin goat CIDEc protein, there were 32.38% of equenxes, 12.70% of quturns, 13.11% of extended chains and 41.80% of irregular coils. The CIDEc gene was expressed in all 7 tissues and organs of Longlin goats, and the CIDEc gene was highly expressed in abdominal fat and subcutaneous fat, which was extremely significantly higher than that in other organs and tissues(P<0.01). The relative expression level of CIDEc gene in adipose tissue(subcutaneous adipose tissue and abdominal adipose tissue) of Nubia goat was significantly higher than that of Longlin goat(P<0.05). 【Conclusion】CIDEc gene is expressed in all organs and tissues of Longlin goat, especially in adipose tissue, and its expression level in adipose tissue(subcutaneous fat and abdominal fat) of Nubian goat is significantly higher than that in Longlin goat. CIDEc protein is an important regulator of fat metabolism and is closely related to lipid storage in animals.
Key words: Longlin goat; CIDEc gene; cloning; bioinformatics analysis; fat metabolism; regulatory mechanism
Foundation item: Guangxi Innovation-driven Development Special Project(Guike AA17204052); Guangxi Key Research and Development Project(Guike AB18221067); Central Leading Local Science and Technology Development Special Project(Guike ZY18076011); Science and Technology Project of Chongzuo City(Chongke FC2018006)
0 引言
【研究意义】隆林山羊是广西地方优良的肉用山羊品种,具有生长发育快、耐粗饲、适应能力强及繁殖率高等特点(吴海,2011),其肉质高蛋白、低胆固醇,具有很好的营养保健功能,而深受人们喜爱。肌间脂肪含量是评价羊肉品质的重要指标,而诱导细胞凋亡DFF45样效应因子c基因(CIDEc)是调节肌间脂肪的候选基因。CIDEc基因高表达于白色脂肪组织,其次是褐色脂肪组织(Inohara et al.,1998)。在脂肪细胞中敲低CIDEc基因将导致脂解明显增加,大脂滴分解成小脂滴(Puri and Czech,2008),即CIDEc基因在脂质代谢平衡中发挥重要作用(Goodman,2008)。因此,加强隆林山羊CIDEc基因功能研究对改善其羊肉品质具有重要意义。【前人研究进展】在诱导细胞凋亡的DFF45(Cell death-inducing DNA fragmentationfactor 45-like effectors,CIDE)蛋白家族中已发现3个成员(CIDEa、CIDEb和CIDEc),且均属于脂滴蛋白(李艳华,2008;Li et al.,2009)。CIDEc在小鼠上又称为鼠脂肪特异蛋白(Fat-specificprotein of 27 kD,FSP27)。CIDEc基因不仅具有诱导细胞调亡的功能,还与动物体内脂肪的分解和形成有关。CIDEc基因在啮齿动物中可定位于脂肪细胞的脂滴表面,且特异富集在脂滴—脂滴接触部位(LDCSs),在脂肪细胞中促进甘油三酯聚集、增大脂滴体积,进而调节脂肪代谢和机体能量平衡(Barneda et al.,2015;Langhi and Baldán,2015;Zhou et al.,2015)。此外,CIDEc在脂肪肝的脂解、胰岛素敏感性和中性脂质(TAG)积累过程中发挥重要作用(Liangpunsakul and Gao,2016;Xu et al.,2016)。如在高脂喂养的小鼠和OB小鼠中使CIDEc基因沉默,均会导致内脏脂肪减少,全身血糖得到有效控制(Langhi et al.,2017),且CIDEc基因敲除小鼠对高脂饮食诱导的体重增加有一定抵抗力(Tanaka et al.,2015)。在脂肪组织中,CIDEc能刺激TAG液滴形成,而抑制非酯化脂肪酸的β-氧化反应(Liang-punsakul and Gao,2016)。Danesch等(1992)研究发现CIDEc基因仅在成熟的脂肪组织中表达,但不在生长或汇合的前脂肪细胞和非脂防细胞中表达,且其表达受转录因子C/EBP的调控(Wu et al.,2002)。Qian等(2017)研究发现,组蛋白脱乙酰基酶6(HDAC6)能促使CIDEc发生脱乙酰基作用,导致其乙酰化水平降低及快速降解,而抑制脂滴融合生长和脂质储存。CIDEc蛋白主要表达于脂肪组织中,与机体的能量代谢密切相关,在动物机体中敲除CIDEc基因最明显的变化就是脂肪组织中脂质明显减少或动物发展成脂肪营养不良表型(Zhou et al.,2015)。缺失CIDEc基因的哺乳动物通常表现为瘦表型、高耗能和胰岛素拮抗等特点(高磊等,2015)。如CIDEc基因敲除小鼠并未呈现出与细胞凋亡相关的表型,而是表现为脂代谢过程的改变(Zhou et al.,2003;Li et al.,2007;Nishino et al.,2008;Toh et al.,2008),即CIDEc基因敲除小鼠體型偏瘦,整体代谢率明显提高,线粒体和小脂滴数量明显增加,葡萄糖吸收及胰岛素敏感性显著增强(Danesch et al.,1992;Wu et al.,2002),以抵抗由食物诱导的肥胖和糖尿病(Zhou et al.,2003)。【本研究切入点】CIDEc基因作为一种与脂肪代谢有关的基因,在脂质贮积、脂滴形成及脂肪分解方面发挥关键作用,目前在人类、小鼠和猪体内已有较多报道(李艳华,2008;Barneda et al.,2015;Langhi and Baldán,2015;Zhou et al.,2015),但鲜见针对隆林山羊CIDEc基因的相关研究。【拟解决的关键问题】以隆林山羊为研究对象,克隆CIDEc基因并对其编码区(CDS)进行序列分析及蛋白理化性质分析等,采用实时荧光定量PCR检测CIDEc基因在不同品种山羊不同组织器官中的表达差异,为揭示CIDEc基因对山羊脂肪代谢的调控机制提供理论依据。
1 材料与方法
1. 1 试验材料
试验动物为相同饲养条件下健康、生长良好的6月龄隆林山羊和努比亚山羊各3头,均由广西大学动物科学技术学院教学实习基地提供。放血宰杀后迅速采集山羊的背最长肌、心脏、肝脏、脾脏、肾脏、腹脂和皮下脂肪等7种组织样品,迅速置于液氮中,稍后转移至-80 ℃冰箱保存备用。Premix Taq DNA聚合酶、pMD18-T载体、反转录试剂盒、定量试剂盒及大肠杆菌Trans5α感受态细胞购自TaKaRa公司;DS5000 DNA Marker、乙醇、氯仿、异丙醇、Solution 1、RNA提取剂TRIzol及胶回收试剂盒购自生工生物工程(上海)股份有限公司;琼脂糖购自法国Biowest公司。
1. 2 试验方法
1. 2. 1 引物设计与合成 根据NCBI已公布的山羊CIDEc基因序列(XM_018038446.1),利用Oligo 7设计特异性引物(表1),即PCR引物和实时荧光定量PCR引物。委托生工生物工程(上海)股份有限公司合成。
1. 2. 2 CIDEc基因克隆 采用TRIzol法提取隆林山羊背最长肌、心脏、肝脏、脾脏、肾脏、腹脂和皮下脂肪及努比亚山羊腹脂和皮下脂肪的总RNA,经紫外分光光度计测定其浓度和质量合格后,反转录合成cDNA。以反转录合成的cDNA为模板,利用引物CIDEc-F和CIDEc-R扩增隆林山羊CIDEc基因序列。PCR反应体系10.0 μL:2×Taq Master Mix 5.0 μL,上、下游引物各0.3 μL,cDNA模板0.5 μL,RNase-free H2O 3.9 μL。扩增程序:95 ℃预变性3 min;95 ℃ 15 s,56 ℃ 30 s,72 ℃ 30 s,进行32个循环;72 ℃延伸10 min。以1.0%琼脂糖凝胶电泳检测PCR扩增产物,胶回收试剂盒回收目的条带,然后将纯化后的PCR扩增产物连接到pMD18-T载体上,连接体系10.0 μL(Solution 1 5.0 μL,PCR扩增产物4.0 μL,pMD18-T载体1.0 μL)。4 ℃连接过夜后转化Trans5α感受态细胞,挑取单克隆菌落培养,筛选出阳性菌液送至生工生物工程(上海)股份有限公司进行测序分析。
1. 2. 3 实时荧光定量PCR检测CIDEc基因组织表达谱 采用实时荧光定量PCR检测CIDEc基因在隆林山羊7种组织和努比亚山羊2种组织中的表达情况,以TRIzol法提取组织RNA反转录合成的cDNA为模板,GAPDH为内参基因,使用CIDEc-F1和CIDEc-R1进行实时荧光定量PCR扩增。每个组织样品进行3次重复。实时荧光定量PCR反应体系10.0 μL:TB GreenTM Premix Ex TaqTM II 5.0 μL,上、下游引物各0.4 μL,cDNA模板2.5 μL,RNase free H2O 1.7 μL。扩增程序:95 ℃预变性30 s;95 ℃ 5 s,60 ℃ 30 s,进行39个循环。采用2-△△Ct法换算CIDEc基因的相对表达量,并以SPSS 18.0中的单因素方差分析(One-way ANOVA)进行显著性检验。
1. 2. 4 隆林山羊CIDEc基因生物信息学分析 使用DNAStar 7.1中的SeqMAN和EditSeq对隆林山羊CIDEc基因序列进行拼接;运用NCBI中的BLAST对隆林山羊CIDEc基因进行同源性比对;采用Meg-Align对隆林山羊CIDEc基因进行物种同源性比对及构建系统发育进化树,参考物种来自NCBI已公布的绵羊(Ovis aries:KM_199684.1)、人类(Homo sapiens:NM_001321043.1)、猕猴(Macaca mulatta:XM_028844410.1)、大鼠(Rattus norvegicus:NM_001170467.1)、小鼠(Mus musculus:NM_178373.4)和猪(Sus scrofa:NM_001112689)。使用ExPASy(http://web.expasy.org/protparam/)在线分析隆林山羊CIDEc基因编码蛋白的氨基酸组成、分子质量及理论等电点等;运用TMHMM Server v.2.0(http://www.cbs.dtu.dk/services/TMHMM/)预测隆林山羊CIDEc蛋白质跨膜区;采用ProtScale(http://web.expasy.org/protscale/)预测隆林山羊CIDEc蛋白亲/疏水性;并分别以NPS@SOPMA和SWISS-MODEL预测隆林山羊CIDEc蛋白的二、三级结构。
2 结果与分析
2. 1 隆林山羊CIDEc基因扩增结果
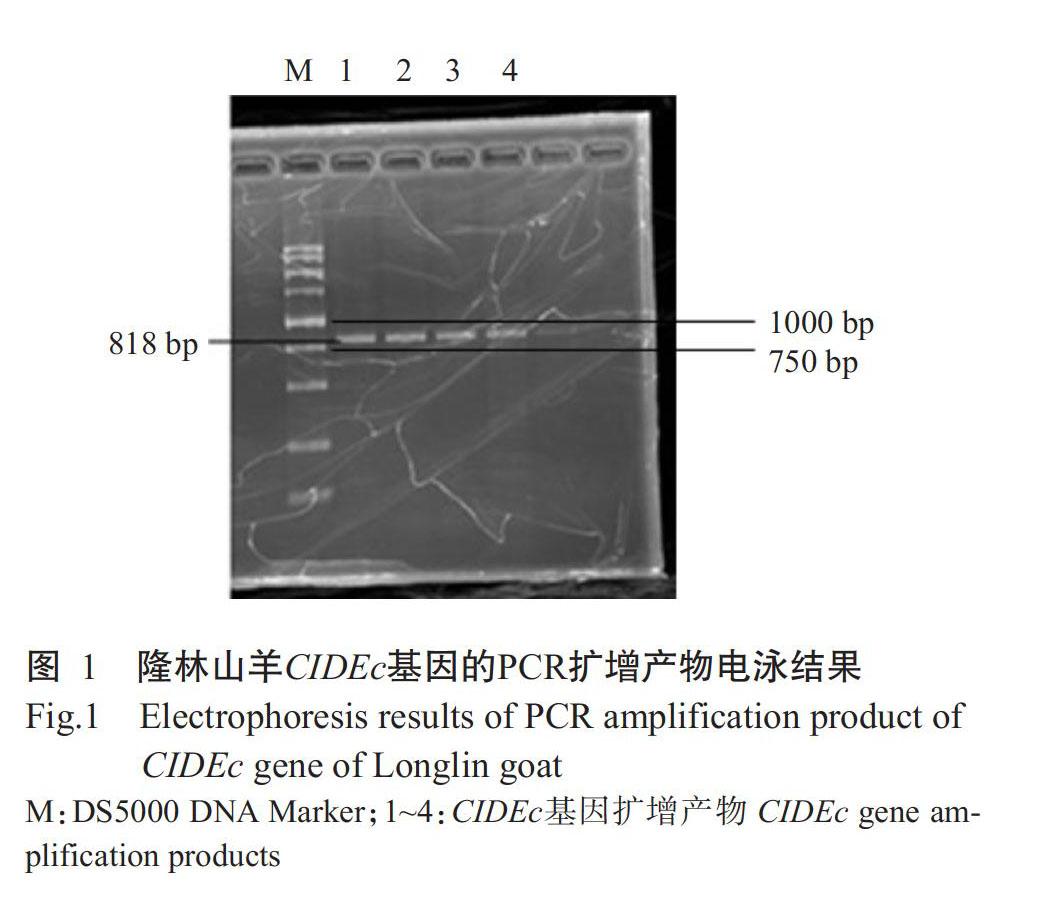
PCR扩增获得的隆林山羊CIDEc基因经1.0%琼脂糖凝胶电泳检测,结果获得单一、明亮清晰、大小约800 bp的特异性条带(图1),与预期结果相符。
2. 2 隆林山羊CIDEc基因序列分析结果
序列分析结果表明,隆林山羊CIDEc基因CDS序列全长741 bp,共编码244个氨基酸残基。将该序列与NCBI已公布的山羊CIDEc基因序列(XM_018038446.1)进行比对分析,结果显示二者的相似性为99.9%,仅有1处碱基发生突变(图2),即在第560位点G突变为T,属于同义突变。
2. 3 隆林山羊CIDEc基因同源性比对分析及系统发育进化树的构建
使用DNAStar 7.1中的MegAlign对隆林山羊CIDEc基因CDS序列与绵羊、人类、猕猴、大鼠、小鼠和猪的CIDEc基因CDS序列进行同源性比对分析,结果显示其相似性分别为99.0%、85.2%、85.8%、80.6%、79.6%和87.2%,即隆林山羊与绵羊的CIDEc基因CDS序列相似性最高,与小鼠的相似性较低。采用MegAlign构建基于CIDEc基因CDS序列相似性的系统发育进化树,结果(圖3)也显示隆林山羊与绵羊的亲缘关系最近,与小鼠的亲缘关系较远。
2. 4 隆林山羊CIDEc蛋白生物信息学分析结果
2. 4. 1 隆林山羊CIDEc蛋白理化性质预测结果
由ExPASy在线分析结果可知,隆林山羊CIDEc蛋白分子式为C1164H1797N323O339S11,分子量为26.09 kD,不稳定系数为48.44,即该蛋白为不稳定蛋白。在编码的244个氨基酸残基中,以甘氨酸(Gly)和亮氨酸(Leu)的含量最高,分别占14.8%;天冬氨酸(Asp)含量最低,仅占0.4%;序列N端残基为丝氨酸(Ser)。隆林山羊CIDEc蛋白的脂肪指数为100.7,理论等电点(pI)为5.28,属于酸性蛋白。
2. 4. 2 隆林山羊CIDEc蛋白跨膜区预测结果 使用TMHMM Server v.2.0对隆林山羊CIDEc蛋白进行跨膜区预测,结果(图4)显示隆林山羊CIDEc蛋白不存在跨膜结构,属于膜外蛋白。
2. 4. 3 隆林山羊CIDEc蛋白亲/疏水性预测结果
采用ProtScale预测隆林山羊CIDEc蛋白亲/疏水性,图中的正值表示疏水性,负值表示亲水性。由图5可看出,其最大值为2.258,最小值为-2.013,综合可知隆林山羊CIDEc蛋白的亲水性较强,属于亲水蛋白。
2. 4. 4 隆林山羊CIDEc蛋白二、三级结构预测结果 使用NPS@SOPMA对隆林山羊CIDEc蛋白二级结构进行预测,结果(图6)显示,在隆林山羊CIDEc蛋白二级结构中,α-螺旋(蓝色小写字母h)占32.38%,β-转角(绿色小写字母t)占12.70%,延伸链(红色小写字母e)占13.11%,无规则卷曲(黄色小写字母c)占41.80%。利用SWISS-MODEL对隆林山羊CIDEc蛋白三级结构进行预测,其预测结果(图7)暗示隆林山羊CIDEc蛋白存在某些功能。
2. 5 山羊CIDEc基因组织表达差异分析结果
2. 5. 1 隆林山羊CIDEc基因组织表达差异 实时荧光定量PCR检测结果(图8)表明,CIDEc基因在隆林山羊背最长肌、心脏、肝脏、脾脏、肾脏、腹脂和皮下脂肪等组织中均有表达,且在腹脂和皮下脂肪中高表达,极显著高于在其他器官组织的表达水平(P<0.01)。CIDEc基因在隆林山羊腹脂中的相对表达量最高,在皮下脂肪中的相对表达量次之,而在心脏、肝脏和背最长肌中的表达量相对较低。
2. 5. 2 隆林山羊和努比亚山羊脂肪中的CIDEc基因表达差异 对比隆林山羊与努比亚山羊CIDEc基因在脂肪(皮下脂肪和腹脂)中的相对表达量,结果(图9)显示CIDEc基因在努比亚山羊脂肪(皮下脂肪和腹脂)中的相对表达量显著高于在隆林山羊脂肪中的相对表达量(P<0.05),即CIDEc基因在努比亚山羊皮下脂肪和腹脂中的表达量均高于隆林山羊。
3 讨论
在正常情况下,脂肪在动物机体内均处于能量代谢稳定状态,一旦这个状态被打破,即有可能引起一系列代谢紊乱疾病,如糖高血压、肥胖和脂防肝等(黄柳梅,2016)。CIDEc基因作为细胞凋亡因子,在3T3-L1细胞中已证实其过表达可诱导细胞凋亡(Keller et al.,2008;李烦繁,2010)。近年来,关于CIDE蛋白的研究主要集中在脂质代谢功能方面(Xu et al.,2012),且越来越多研究证明CIDEc脂滴特异性蛋白高表达于脂肪组织,可促进细胞内脂肪积累(李红强等,2017),在调控动物体脂质代谢平衡方面发挥重要作用(向梳瑕等,2010)。因此,了解CIDEc基因对隆林山羊脂肪细胞分化和脂肪沉积的调控机制,对改善其羊肉品质具有重要意义。本研究从隆林山羊组织中克隆出CIDEc基因CDS序列并进行生物信息學分析,通过实时荧光定量PCR检测CIDEc基因在隆林山羊和努比亚山羊脂肪组织(皮下脂肪和腹脂)中的表达差异,以期为进一步研究CIDEc基因的功能及其在脂质代谢中的作用提供理论依据。
He等(2018)研究表明,在脂多糖(LPS)诱导的炎症反应中CIDEc可通过与腺苷5'-单磷酸激活蛋白激酶(AMPK)α亚基相互作用,直接下调AMPK活性,AMPK可抑制LPS诱导的核因子κB(NF-κB)活化,而产生抗炎作用。沉默CIDEc基因还可减少LPS诱导的上皮细胞凋亡,即CIDEc/AMPK信号通路在LPS诱导的炎症和上皮细胞凋亡中发挥重要作用。Tan等(2016)研究发现,肿瘤坏死因子α(TNF-α)可下调CIDEc基因表达水平以增强机体脂解作用。CIDEc广泛参与动物体内信号传导、生化反应等,尤其更偏向于参与对机体脂肪的调控。本研究结果表明,山羊CIDEc基因在脂肪组织中的相对表达量极显著高于在其他器官组织的相对表达量,与Wang等(2013)研究发现牛CIDEc基因在脂肪组织中高表达的结论一致。山羊CIDEc基因CDS序列与人类CIDEc基因CDS序列的相似性为85.2%,而与绵羊、猕猴、大鼠、小鼠和猪CIDEc基因CDS序列的相似性为99.0%、87.2%、79.6%、80.6%和85.8%,说明CIDEc基因在不同哺乳动物中的保守性较高。基于CIDEc基因CDS序列相似性构建的系统发育进化树也显示,隆林山羊与绵羊和猪等哺乳动物及人类的遗传距离相对较近,而与大鼠和小鼠相距较远,与物种分类学保持较高的一致性。李红强等(2020)研究发现CIDEc能招募其他调控蛋白而促进脂滴的融合和生长。本研究对隆林山羊CIDEc蛋白的二、三级结构进行预测,结果发现CIDEc蛋白确实能形成二级结构乃至更高的三级结构,预示着其存在重要的生理作用与功能。
本研究还发现,CIDEc基因在努比亚山羊脂肪组织(皮下脂肪和腹脂)中的相对表达量显著高于其在隆林山羊中的相对表达量。曹艳红(2019)的研究显示,努比亚山羊和隆林山羊在生长速度方面差异极显著,2种羊肉中的不饱和脂肪酸含量也差异显著,究其原因可能与努比亚山羊生长速度快,而隆林山羊生长速度较慢有关。此外,Liang等(2003)研究发现CIDEc作为FSP27的人类同源物,与FSP27有60%的相似性。FSP27仅在成熟脂肪细胞中表达(Danesch et al.,1992),与脂肪细胞分化密切相关,但由于存在种属间差异,因此有关山羊CIDEc基因对脂肪相关代谢机制的调控作用尚有待进一步探究。
4 结论
CIDEc基因在隆林山羊各器官组织中均有表达,以在脂肪组织中的表达量较丰富,且其在努比亚山羊脂肪组织(皮下脂肪和腹脂)中的表达水平显著高于在隆林山羊中的表达水平。可见,CIDEc蛋白是脂肪代谢的重要调节剂,与动物体内脂质存储密切相关。
参考文献:
曹艳红. 2019. 利用高通量测序解析隆林山羊与努比亚山羊的遗传差异及表达差异[D]. 杨凌:西北农林科技大学. doi:10.27409/d.cnki.gxbnu.2019.000029. [Cao Y H. 2019. Analysis of genetic and expression differences between Longlin goat and Nubia goat by high throughput sequen-cing[D]. Yangling:Northwest A & F University.]
高磊,许瑞霞,赵伟利,杨井泉,梁耀伟,刘守仁,沈敏,王新华. 2015. 绵羊诱导细胞凋亡的DFF45样效应因子c基因(CIDEC)克隆及其在持续饥饿条件下阿勒泰羊尾脂组织中的差异表达[J]. 农业生物技术学报,23(2):227-235. doi:10.3969/j.issn.1674-7968.2015.02.010. [Gao L,Xu R X,Zhao W L,Yang J Q,Liang Y W,Liu S R,Shen M,Wang X H. 2015. Cloning of sheep cell death- inducing DFFA-like effector c(CIDEC) cDNA and its differential expression in tail fat tissue of Altay sheep (Ovis aries) in persistent starvation[J]. Journal of Agricultural Biotechnology,23(2):227-235.]
黄柳梅. 2016. CIDEa和CIDEc基因在肉鸡中的表达分析[D]. 南昌:江西农业大学. [Huang L M. 2016. Expression analysis of CIDEa and CIDEc genes in broiler chickens[D]. Nanchang:Jiangxi Agricultural University.]
李烦繁. 2010. CIDEC在人脂肪细胞分化中的作用及其机制研究[D]. 西安:第四军医大学. doi:10.7666/d.d219379. [Li F F. 2010. The role and possible mechanism of CIDEC in human adipocyte differentiation[D]. Xi?an:Fourth Mi-litary Medical University.]
李红强,郭振清,杜金友. 2020. 诱导细胞凋亡DFF45样效应因子C的结构与表达调控及其功能[J]. 中国生物化学与分子生物学报,36(5):512-518. doi:10.13865/j.cnki.cjbmb.2020.02.1532. [Li H Q,Guo Z Q,Du J Y. 2020. Structural,expression and functional analysis of cell dea-thinducing DNA fragmentation factor 45-like effector C(CIDEC)[J]. Chinese Journal of Biochemistry and Molecular Biology,36(5):512-518.]
李紅强,李海玉,李婧实,高飞飞,李鑫波,郭振清. 2017. CIDEC的亚细胞定位及其功能初步研究[J]. 核农学报,31(8):1494-1499. doi:10.11869/j.issn.100-8551.2017.08. 1494. [Li H Q,Li H Y,Li J S,Gong F F,Li X B,Guo Z Q. 2017. Localization and preliminarily functional analysis of CIDEC[J]. Journal of Nuclear Agricultural Scien-ces,31(8):1494-1499.]
李艳华. 2008. 猪CIDE家族基因的克隆及功能研究[J]. 武汉:华中农业大学. doi:10.7666/d.y1394601. [Li Y H. 2008. Cloning and functional research of porcine CIDEs family[J]. Wuhan:Huazhong Agricultural University.]
吴海. 2011. 隆林县发展隆林山羊现状及前景分析[J]. 广西畜牧善医,27(4):235-236. doi:10.3969/j.issn.1002-5235. 2011.04.019. [Wu H. 2011. Analysis of present situation and prospect of developing Longlin goats in Longlin County[J]. Guangxi Journal of Animal Husbandry & Ve-terinary Medicine,27(4):235-236.]
向梳瑕,潘志雄,李亮,王继文. 2010. DNA断裂因子相似蛋白(CIDE)家族在甘油三酯代谢中的作用[J]. 生命科学,22(6):575-578. doi:10.13376/j.cbls/2010.06.004. [Xiang S X,Pan Z X,Li L,Wang J W. 2010. The role of cell-death-inducing DNA-fragmentation-factor-like effector (CIDE) family in TG metabolism[J]. Chinese Bulletin of Life Sciences,22(6):575-578.]
Barneda D,Planas-Iglesias J,Gaspar M L,Mohammadyani D,Prasannan S,Dormann D,Han G S,Jesch S A,Carman G M,Kagan V,Parker M G,Ktistakis N T,Klein-Seetha-raman J,Dixon A M,Henry S A,Christian M. 2015. The brown adipocyte protein CIDEA promotes lipid droplet fusion via a phosphatidic acid-binding amphipathic helix[J]. eLife,4:e07485. doi:10.7554/eLife.07485.
Danesch U,Hoeck W,Ringold G M. 1992. Cloning and transcriptional regulation of a novel adipocyte-specific gene,FSP27. CAAT-enhancer-binding protein(C/EBP) and C/EBP-like proteins interact with sequences required for differentiation-dependent expression[J]. The Journal of Biological Chemistry,267(10):7185-7193.
Goodman J M. 2008. The gregarious lipid droplet[J]. The Journal of Biological Chemistry,283(42):28005-28009. doi:10.1074/jbc.R800042200.
He J,Zhang B,Gan H. 2018. CIDEC is involved in LPS-induced inflammation and apoptosis in renal tubular epithelial cells[J]. Inflammation,41(5):1912-1921. doi:10.1007/ s10753-018-0834-3.
Inohara N,Koseki T,Chen S,Wu X,Nú?ez G. 1998. CIDE,a novel family of cell death activators with homology to the 45 kDa subunit of the DNA fragmentation factor[J]. The EMBO Journal,17(9):2526-2533. doi:10.1093/emboj/17.9.2526.
Keller P,Petrie J T,de Rose P,Gerin I,Wright W S,Chiang S H,Nielsen A R,Fischer C P,Pedersen B K,MacDougald O A. 2008. Fat-specific protein 27 regulates storage of triacylglycerol[J]. The Journal of Biological Chemistry,283(21):14355-14365. doi:10.1074/jbc.M708323200.
Langhi C,Arias N,Rajamoorthi A,Basta J,Lee R G,Baldán ?. 2017. Therapeutic silencing of fat-specific protein 27 improves glycemic control in mouse models of obesity and insulin resistance[J]. Journal of Lipid Research,58(1):81-91. doi:10.1194/jlr.M069799.
Langhi C,Baldán A. 2015. CIDEC/FSP27 is regulated by pero-xisome proliferator-activated receptor alpha and plays a critical role in fasting- and diet-induced hepatosteatosis[J]. Hepatology,61(4):1227-1238. doi:10.1002/hep. 27607.
Li J Z,Ye J,Xue B F,Qi J Z,Zhang J,Zhou Z H,Li Q,Wen Z L,Li P. 2007. Cideb regulates diet-induced obesity,li-ver steatosis,and insulin sensitivity by controlling lipogenesis and fatty acid oxidation[J]. Diabetes,56(10):2523-2532. doi:10.2337/db07-0040.
Li Y H,Lei T,Chen X D,Xia T,Peng Y,Long Q Q,Zhang J,Feng S Q,Zhou L,Yang Z Q. 2009. Molecular cloning,chromosomal location and expression pattern of porcine CIDEa and CIDEc[J]. Molecular Biology Report,36(3):575-582. doi:10.1007/s11033-008-9216-5.
Liang L,Zhao M J,Xu Z H,Yokoyama K K,Li T. 2003. Molecular cloning and characterization of CIDE-3,a novel member of the cell-death-inducing DNA-fragmentation-factor (DFF45)-like effector family[J]. The Biochemical Journal,370(1):195-203. doi:10.1042/BJ20020656.
Liangpunsakul S,Gao B. 2016. Alcohol and fat promote stea-tohepatitis:A critical role for fat-specific protein 27/CIDEC[J]. Journal of Investigative Medicine,64(6):1078-1081. doi: 10.1136/jim-2016-000204.
Nishino N,Tamori Y,Tateya S,Kawaguchi T,Shibakusa T,Mizunoya W,Inoue K,Kitazawa R,Kitazawa S,Matsuki Y,Hiramatsu R,Masubuchi S,Omachi A,Kimura K,Saito M,Amo T,Ohta S,Yamaguchi T,Osumi T,Cheng J L,Fujimoto T,Nakao H,Nakao K,Aiba A,Okamura H,Fushiki T,Kasuga M. 2008. Fsp27 contributes to efficient energy storage in murine white adipocytes by promoting the formation of unilocular lipid droplets[J]. The Journal of Clinical Investigation,118(8):2808-2821. doi:10.1172/JCI34090.
Puri V,Czech M P. 2008. Lipid droplets:FSP27 knockout enhances their sizzle[J]. The Journal of Clinical Investigation,118(8):2693-2696. doi:10.1172/JCI36554.
Qian H,Chen Y,Nian Z,Su L,Yu H Y,Chen F J,Zhang X Q,Xu W Y,Zhou L K,Liu J M,Yu J H,Yu L X,Gao Y,Zhang H C,Zhang H H,Zhao S M,Yu L,Xiao R P,Bao Y Q,Hou S C,Li P P,Li J D,Deng H T,Jia W P,Li P. 2017. HDAC6-mediated acetylation of lipid droplet-bin-ding protein CIDEC regulates fat-induced lipid storage[J]. The Journal of Clinical Investigation,127(4):1353-1369. doi:10.1172/JCI85963.
Tan X R,Cao Z Z,Li M,Xu E,Wang J J,Xiao Y F. 2016. TNF-α downregulates CIDEC via MEK/ERK pathway in human adipocytes[J]. Obesity(Silver Spring),24(5):1070-1080. doi:10.1002/oby.21436.
Tanaka N,Takahashi S,Matsubara T,Jiang C,Sakamoto W,Chanturiya T,Teng R,Gavrilova O,Gonzalez F J. 2015. Adipocyte-specific disruption of fat-specific protein 27 causes hepatosteatosis and insulin resistance in high-fat diet-fed mice[J]. The Journal of Biological Chemistry,290(5):3092-3105. doi:10.1074/jbc.M114.605980.
Toh S Y,Gong J Y,Du G L,Li J Z,Yang S Q,Ye J,Yao H L,Zhang Y X,Xue B F,Li Q,Yang H Y,Wen Z L,Li P. 2008. Up-regulation of mitochondrial activity and acquirement of brown adipose tissue-like property in the white adipose tissue of fsp27 deficient mice[J]. PLoS One,3(8):e2890. doi:10.1371/journal.pone.0002890.
Wang J,Cao X K,Pan H,Hua L S,Yang M J,Lei C Z,Lan X Y,Chen H. 2013. Cell death-inducing DFFA-like effector c(CIDEC/Fsp27) gene:Molecular cloning,sequence characterization,tissue distribution and polymorphisms in Chinese cattles[J]. Molecular Biololoy Reports,40(12):6765-6774. doi:10.1007/s11033-013-2793-y.
Wu X L,Shi Q S,Liu X C,Peng Y L,Jiang J,Yang S L,Xiao B N. 2002. A search for chromosome 13 QTL in a complex pig family[J]. Acta Genetics Sinica,29(1):1-6.
Xu L,Zhou L K,Li P. 2012. CIDE proteins and lipid metabolism[J]. Arteriosclerosis Thrombosis & Vascular Biology,32(5):1094-1098.
Xu W,Wu L,Yu M,Chen F J,Arshad M,Xia X Y,Ren H,Yu J H,Xu L,Xu D J,Li J Z,Li P,Zhou L K. 2016. Differential roles of cell death-inducing DNA fragmentation factor-α-like effector(CIDE) proteins in promoting lipid droplet fusion and growth in subpopulations of hepatocytes[J]. The Journal of Biological Chemistry,291(9):4282-4293. doi:10.1074/jbc.M115.701094.
Zhou L K,Park S Y,Xu L,Xia X Y,Ye J,Su L,Jeong K H,Hur J H,Oh H,Tamori Y,Zingaretti C M,Cinti S,Argente J,Yu M,Wu L Z,Ju S H,Guan F F,Yang H Y,Choi C S,Savage D B,Li P. 2015. Insulin resistance and white adipose tissue inflammation are uncoupled in energetically challenged Fsp27-deficient mice[J]. Nature Communications,6:5949. doi:10.1038/ncomms6949.
Zhou Z H,Toh Toh S,Chen Z M,Guo K,Ng C P,Ponniah S,Lin S C,Hong W J,Li P. 2003. Cidea-deficient mice have lean phenotype and are resistant to obesity[J]. Nature Genetics,35(1):49-56. doi:10.1038/ng1225.
(責任编辑 兰宗宝)
