GABA和CaCl2复配对盐碱胁迫下番茄幼苗生长、叶绿素荧光参数和活性氧的影响
2021-07-26陈光鑫陈迎雪王叶宁吕晨希高洪波吴晓蕾
陈光鑫 丁 栋 李 倩 陈迎雪 王叶宁 吕晨希 高洪波 吴晓蕾
(河北农业大学园艺学院,河北保定 071001)
土壤盐碱化现象是世界性生态和资源问题,严重限制了农业的发展。据不完全统计,我国盐碱化土壤面积约为990万hm2(张体彬 等,2017),特别是在我国北方地区,盐碱胁迫已成为制约农作物产量和品质的主要非生物胁迫因素之一(姜伟,2010)。土壤盐碱化主要受自然和人为因素共同影响(Li et al.,2015),设施栽培环境由于相对封闭,复种指数高,土壤更易受化肥过量施用及灌溉不足影响而引发次生盐碱化(Li et al.,2015)。不同于中性盐胁迫,盐碱胁迫除引起植物失水萎蔫、营养失衡及离子毒害外,还会造成高pH胁迫(Yang et al.,2007),从而对植物造成更大的伤害。因此,如何有效缓解盐碱胁迫已成为备受关注的课题。
番茄(Solanum lycopersicumL.)是我国广泛栽培的一种重要园艺作物。作为番茄生产大国,2017年我国番茄种植面积为132万hm2,其中近50%为设施栽培。番茄属中度盐敏感型园艺作物(Katerji et al.,2000),随着设施土壤盐碱化的日益加重,对番茄产量和品质形成了较大的威胁。以往的研究大多集中于盐胁迫对植物产生的影响(罗达 等,2019;苏兰茜 等,2019),针对植物盐碱胁迫的响应机制研究还较少。在缓解盐碱胁迫方面,外源物质的添加是提高盐碱胁迫下植物耐性和作物产量的一种简单有效的方法(Jin et al.,2019;赵颖 等,2020)。γ-氨基丁酸(GABA)是动物、植物中广泛存在的一种天然的非蛋白氨基酸,在植物中,GABA作为一种代谢物或信号分子在胁迫条件下的许多生理过程中发挥重要作用(Lang et al.,2016;Zhen et al.,2018),内源GABA可受盐刺激和外源GABA诱导的影响,提高植物抗盐胁迫能力(Shelp et al.,2012;Faës et al.,2015)。钙作为植物必需的大量营养元素之一以及胞内重要的第二信使,对于植物逆境响应非常重要(陈立松和刘星辉,1997)。有研究表明施用外源钙可以降低植物吸收Na+的能力、改善细胞对盐的适应能力(朱义等,2007)。谷氨酸脱羧酶(GAD)是GABA支路中合成GABA的限速酶(Inatomi &Slaughter,1971),在其C-末端有与钙调素结合区域(Yin et al.,2015),而逆境胁迫会造成植物胞内或胞外Ca2+浓度积累从而激活GAD,诱导GABA大量积累(Kinnersley &Turano,2000)。但Ca2+在GABA诱导植物抗逆中的具体作用机制尚不清楚。
为此,在前期针对GABA提高番茄耐盐性研究的基础上(罗黄颖 等,2011),本试验以盐敏感型番茄品种中杂9号为试材(Hu et al.,2009),采用表型差异显著的盐碱胁迫浓度(60 mmol ·L-1NaCl+60 mmol·L-1NaHCO3)(梁杨杨 等,2016),研究单独添加GABA和CaCl2及两者复配组合对盐碱胁迫下番茄幼苗生长、叶绿素荧光和活性氧等指标的影响,探讨外源GABA和钙在提高番茄耐盐碱性过程中相互作用的生理调节机制,为生产中外源物质施用提高番茄耐盐碱性及相关制剂开发应用提供理论和实践依据。
1 材料与方法
1.1 试材培养与盐碱胁迫处理
本试验在河北农业大学西校区园艺学院试验温室内进行,供试番茄品种为中杂9号,购自中国农业科学院蔬菜花卉研究所。GABA购自美国Sigma-Adrich公 司,CaCl2、NaCl、NaHCO3均 为分析纯试剂。前期预备试验已筛选出差异显著的盐碱胁迫处理浓度。2018年5月23日,将番茄种子催芽后,采用72孔穴盘,使用草炭、珍珠岩混配基质进行育苗,浇灌Hoagland’s营养液(pH值6.5,EC值1.8 mS·cm-1)。6月20日,选取整齐一致且长势良好的四叶一心幼苗,洗去幼苗根部基质,定植于容积20 L的灰色塑料盆中进行水培,用充氧泵及时补充氧气,保证番茄幼苗正常生长。用Hoagland’s营养液预培养3 d后,设置如下10个配方进行水培(表1)。每处理3盆,每盆定植50株番茄幼苗。处理后不同时间内取样测定幼苗的生长及生理指标,生长指标每处理取样10株,生理指标取样6株,3次重复。
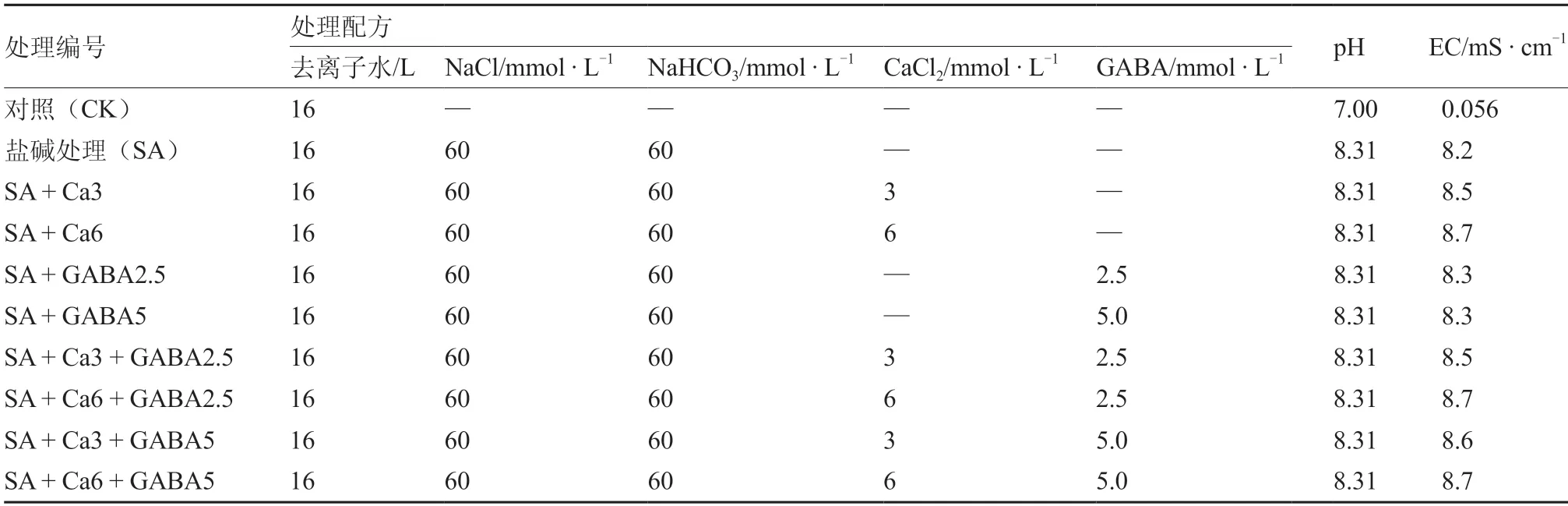
表1 处理编号及具体处理配方
1.2 测定指标和方法
在处理0、2、4、6 d,每处理固定选取10株幼苗测定株高、茎粗,株高测量部位为幼苗茎基部至顶部生长点,株高生长速率按公式〔(测定株高-前一次测定株高)/前一次测定株高×100%〕计算;茎粗测量部位为茎基部以上2 cm,茎粗生长速率按〔(测定茎粗-前一次测定茎粗)/前一次测定茎粗×100%〕计算。在处理0、2、4、6 d,每处理选取均匀一致的10株幼苗,采用LA-S根系扫描仪测定根长和根数,生长速率计算公式同上。于处理6 d后,每处理选取10株幼苗,称量全株鲜质量,在烘箱内105 ℃下杀青10 min后在80 ℃烘干至恒重,测定植株干质量。于处理6 d后,每处理随机选取6株幼苗,采用叶色指数仪(SPAD-502 Plus,日本)测定从上向下数第3片展开叶片SPAD值(叶色指数),叶绿素荧光仪(FluorCam开放式叶绿素荧光成像系统,捷克)测定从上向下数第3片展开叶片叶绿素荧光参数。于处理2、4、6 d,每处理随机选取6株幼苗从上向下数第3、4片展开叶片,参照Thordal-Christensen等(1997)的方法测定超氧阴离子产生速率和H2O2含量。
1.3 数据分析
试验数据使用Excel 2007和SAS 8.1软件进行处理和统计分析,各测定数据均源于3次测试重复结果的平均值。采用单因素和Duncan’s法进行方差分析和差异显著性比较(P<0.05),利用Excel软件绘图,图表中数据为平均值 ± 标准误。
2 结果与分析
2.1 不同GABA和CaCl2 复配组合对盐碱胁迫下番茄幼苗生长的影响
2.1.1 不同GABA和CaCl2复配组合对盐碱胁迫下番茄幼苗株高和茎粗生长速率的影响 由图1-A可知,在整个处理过程中,SA处理的幼苗株高生长速率均显著低于对照,降低幅度随胁迫时间延长而逐渐增加,分别达到39.3%、58.3%、67.8%。外源CaCl2和GABA单独处理和复配处理均可有效缓解盐碱胁迫对番茄幼苗生长的抑制作用,促进株高生长速率增加。其中CaCl2处理中以SA +Ca6处理的株高生长速率较高,较SA处理提高了24.1%~50.2%,差异显著;GABA处理中以SA +GABA5处理的株高生长速率较高,较SA处理提高了23.0%~55.3%,差异显著;复配组合中以SA +Ca6+GABA5的株高生长速率最高,较SA处理提高幅度呈随胁迫时间延长而上升的趋势,在处理6 d时达到79.6%。
如图1-B所示,与对照相比,盐碱胁迫显著抑制了番茄幼苗茎粗的增加,处理2、4、6 d时降低幅度分别达到67.8%、77.8%和80.4%,呈现出随胁迫时间延长降低幅度逐渐增大的趋势。外源CaCl2和GABA单独处理和复配处理均可促进幼苗茎粗的增加,但提高幅度差异较大。其中以SA+Ca3+GABA5、SA+Ca6+GABA5复 配组合效果较好,在整个处理过程中茎粗生长速率均高于其他单独组合和复配组合,相较于SA处理茎粗生长速率分别提高了100.9%~138.6%和103.9%~148.4%,并且相较于其他组合,在胁迫6 d时依然保持了较高的茎粗生长速率。
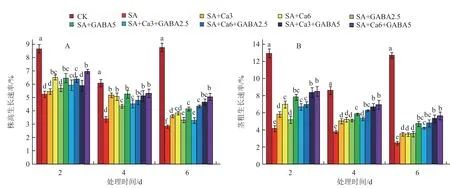
图1 盐碱胁迫下不同GABA和CaCl2 复配组合的株高和茎粗生长速率
2.1.2 不同GABA和CaCl2复配组合对盐碱胁迫下番茄幼苗根长和根数生长速率的影响 由图2-A可以看出,SA处理对于幼苗根系伸长有显著的抑制作用,表现为根长生长速率随处理时间延长而逐渐下降,整个胁迫过程中较对照降低86.3%、91.9%和94.8%。与SA处理相比,在胁迫6 d时,2个浓度的外源GABA单独处理对于根长生长速率的提升达到4倍以上,而外源CaCl2单独处理对于根长生长率的提升仅为2.8倍,表明外源GABA对于根长的缓解作用更好。单独处理的2个浓度相比,较高浓度的CaCl2和GABA效果均好于较低浓度。复配组合中以SA+Ca6+GABA5的根长生长速率最高,在胁迫6 d时依然保持了较高的增长速率,达到SA处理的6.9倍,且在处理4 d和6 d时,显著高于其他外源添加处理。
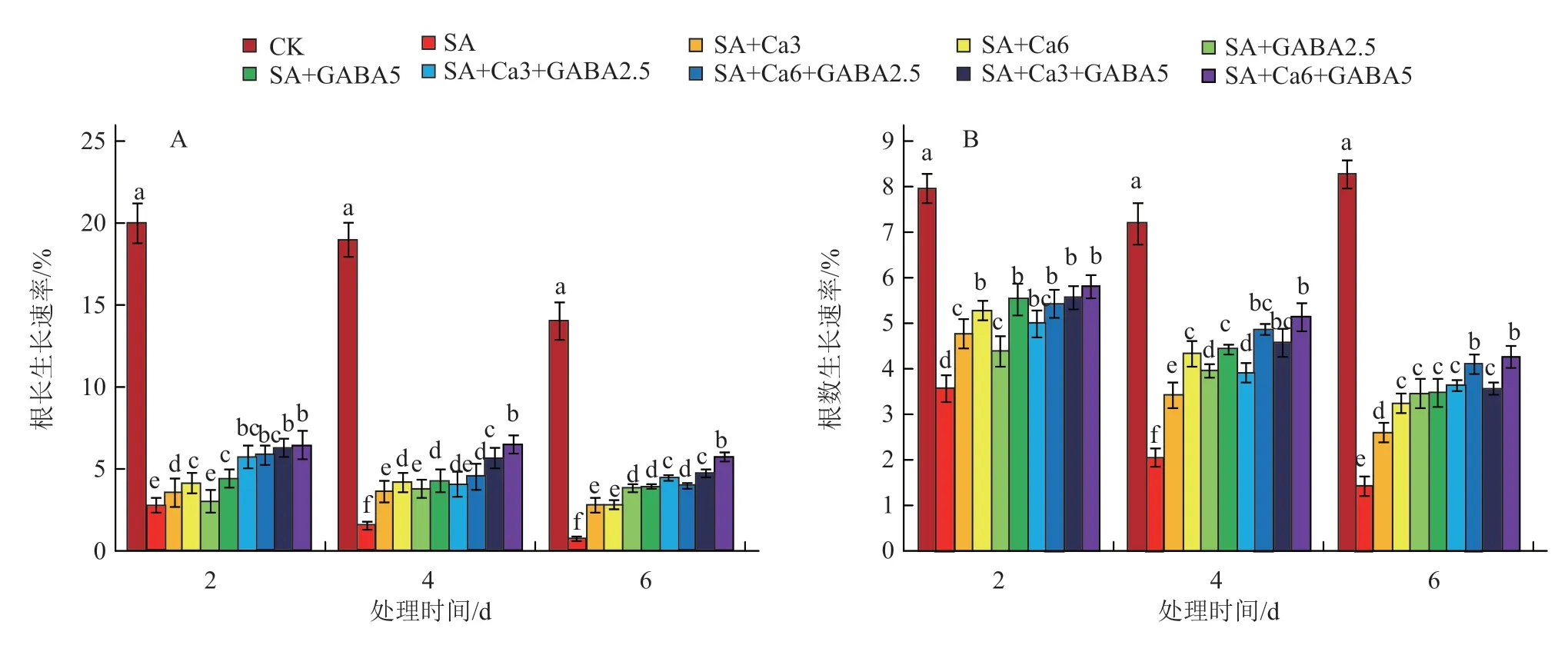
图2 盐碱胁迫下不同GABA和CaCl2 复配组合的根长和根数生长速率
SA处理对于幼苗根数的增加也有显著的抑制作用,与对照相比,根数生长速率的降低幅度分别达到55.0%、71.4%和82.6%(图2-B)。处理6 d时复配组合对于根数的增加作用好于外源GABA单独处理,而外源GABA单独处理好于外源CaCl2单独处理,最优复配组合为SA+Ca6+GABA5,整个处理期间的根数生长速率最高,其次为SA+Ca6+GABA2.5处理,且2个处理间差异不显著。
2.1.3 不同GABA和CaCl2复配组合对盐碱胁迫下番茄幼苗干、鲜质量的影响 由表2可知,SA处理的番茄幼苗鲜质量和干质量与对照存在显著差异,其中鲜质量较对照降低了32.9%,干质量则下降了13.2%,说明盐碱胁迫对于植株鲜质量的影响大于对干质量的影响。与SA处理相比,无论是外源GABA单独处理、外源CaCl2单独处理,还是复配处理,均显著增加了幼苗的鲜质量;除SA +Ca3+GABA2.5处理外,各外源添加处理的干质量均显著高于SA处理。复配处理中以SA+Ca6 +GABA5效果最好,较SA处理鲜质量提高27.7%,干质量提高12.1%。
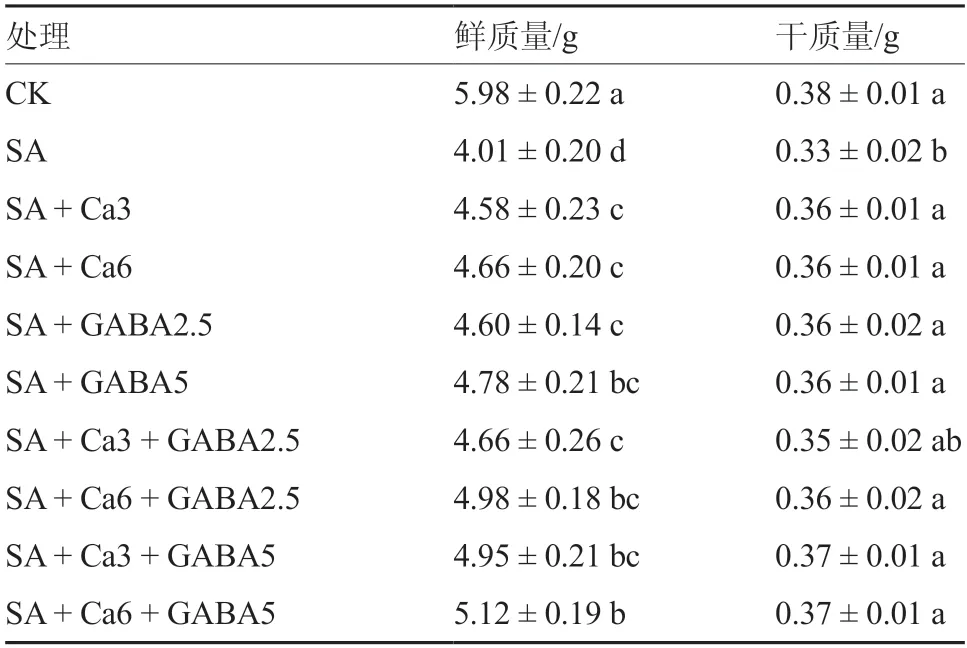
表2 不同GABA和CaCl2 复配组合对盐碱胁迫下番茄幼苗干、鲜质量的影响
2.2 不同GABA和CaCl2 复配组合对盐碱胁迫下番茄幼苗叶色指数和叶绿素荧光参数的影响
由表3可以看出,SA处理的幼苗叶片叶色指数显著低于对照和其他外源添加处理,其中较对照降低了30.8%。外源GABA单独处理中SA +GABA5的叶色指数较SA+GABA2.5显著提高,外源CaCl2单独处理中SA+Ca6效果好于SA +Ca3。复配处理中SA+Ca3+GABA5叶色指数最高,其次为SA+Ca6+GABA5处理,且两者差异不显著。
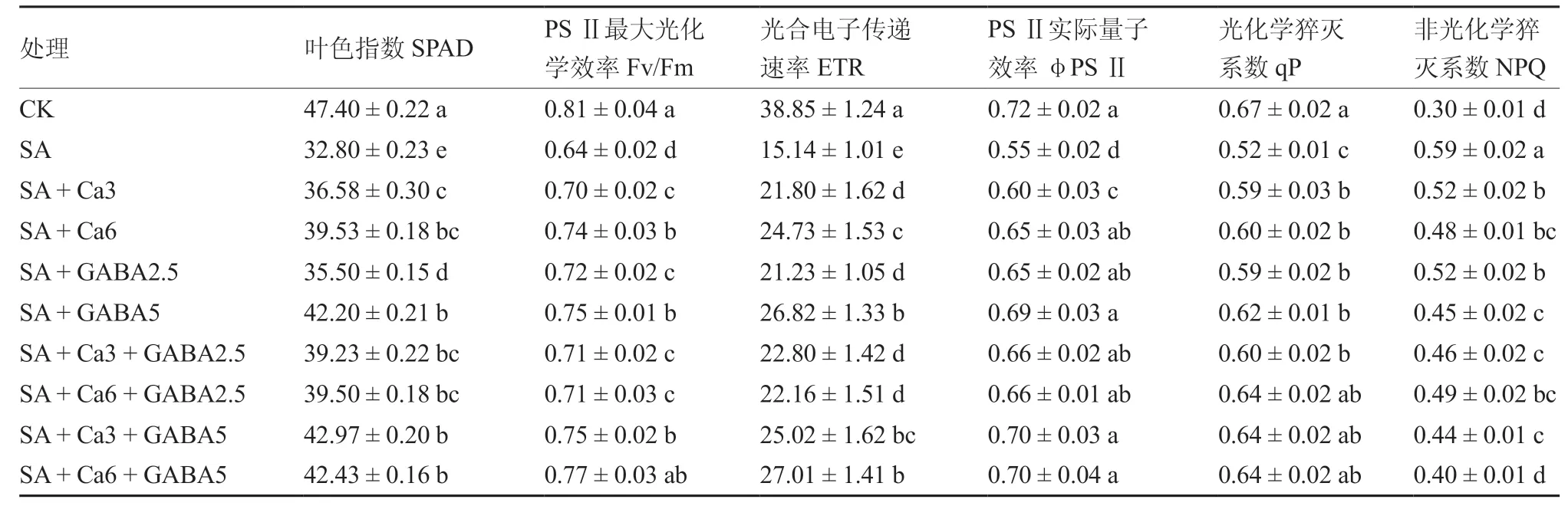
表3 不同GABA和CaCl2 复配组合对盐碱胁迫下番茄幼苗叶色指数和叶绿素荧光参数的影响
SA处理的幼苗叶片叶绿素荧光参数与对照差异显著,表明盐碱胁迫对幼苗叶片光合系统造成了伤害,主要表现为Fv/Fm、ETR、φPS Ⅱ、qP的显著降低,而NPQ则显著上升。各外源添加处理均可显著影响盐碱胁迫下幼苗叶片的叶绿素荧光参数,SA+GABA5处理对于叶绿素荧光系统伤害的缓解效果好于2个外源CaCl2单独处理,表现为ETR显著高于2个外源CaCl2单独处理,Fv/Fm、φPS Ⅱ、qP在各单独处理中最高。复配处理中以SA+Ca6+GABA5和SA+Ca3+GABA5效 果较好,除NPQ外,两者之间没有显著差异。
2.3 不同GABA和CaCl2 复配组合对盐碱胁迫下番茄幼苗活性氧代谢的影响
如图3-A所示,盐碱胁迫处理显著提高了番茄叶片H2O2含量,整个处理过程中SA处理较对照H2O2含量增加94.5%~127.1%,说明盐碱胁迫对植株叶片细胞造成了严重伤害。与SA处理相比,外源添加GABA和CaCl2均可显著降低叶片中H2O2含量。与CaCl2单独处理相比,GABA单独处理对于H2O2的清除具有更强的持续性,整个处理期间H2O2含量降低幅度均超过20%。复配处理中SA+Ca6+GABA5对于H2O2的清除作用最强,H2O2含量降低幅度达到32.5%~39.5%。
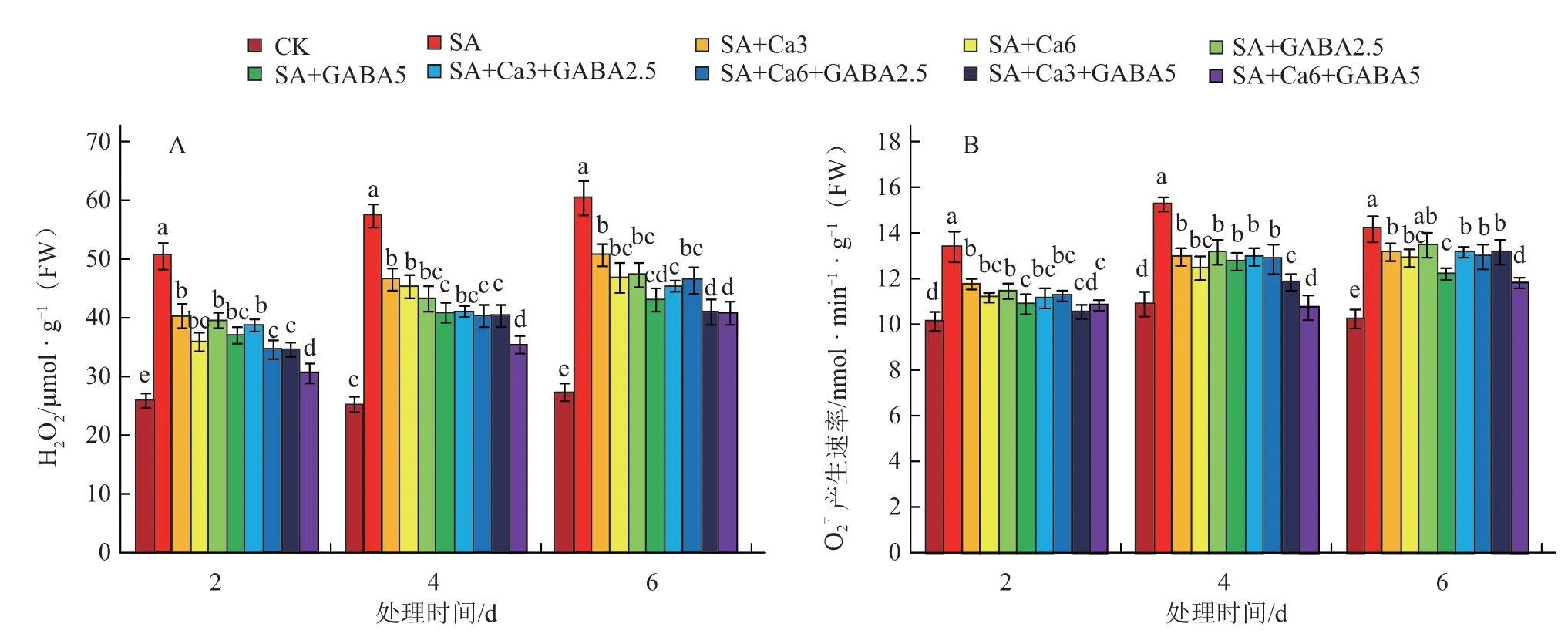
图3 盐碱胁迫下不同GABA和CaCl2复配组合的过氧化氢含量和超氧阴离子产生速率
3 讨论与结论
生物量降低是盐胁迫下植物最明显的表型体现(王素平 等,2006)。盐碱胁迫诱导渗透胁迫和氧化胁迫,干扰植物代谢,进而限制植物生长发育。已有报道指出外源CaCl2可显著提高盐胁迫下大蒜幼苗的株高和根长(Chen et al.,2011)。河北农业大学设施蔬菜课题组前期研究表明,外源GABA施用可提高盐碱胁迫下番茄幼苗地上部和地下部鲜质量(梁杨杨 等,2016)。本试验中,GABA和CaCl2单独使用均有利于盐碱胁迫下番茄幼苗生长,表现为株高、茎粗、根长和根数生长速率以及干鲜质量的提升。GABA和CaCl2存在浓度效应,整体表现为高浓度(6 mmol·L-1CaCl2、5 mmol ·L-1GABA)好于低浓度(3 mmol·L-1CaCl2、2.5 mmol·L-1GABA)。总体来看,添加5 mmol·L-1GABA对于生物量的提升效果好于添加6 mmol ·L-1CaCl2的处理,说明对于上述浓度来说,GABA对于盐碱胁迫的缓解作用大于CaCl2,可能的原因是选用的CaCl2单独处理浓度还未达最适浓度。与单独添加处理相比,4个复配组合在不同程度上缓解了盐碱胁迫对幼苗生长的抑制,其中表现最好的是SA+Ca6+GABA5处理,在外源添加处理中幼苗生长指标均达最高,SA+Ca3+GABA5处理表现次之,且在多个时间点与SA+Ca6+GABA5处理无显著差异。由此可以看出,合理浓度的复配处理整体较单独处理提高了番茄幼苗的耐盐碱性,并且缓解效果主要归功于GABA,当选用较高浓度GABA与不同浓度CaCl2复配时,其组合效果均较好,CaCl2浓度的高低对于复配组合的影响较小。
叶绿素对光合作用至关重要。光合能力下降是盐碱胁迫下植物生长受到抑制的主要原因。盐碱胁迫会降低叶片的光合作用(Xiang et al.,2016;Radanielson et al.,2018)。植物中叶绿素的生物合成过程中任何环节出现问题,叶绿素含量就会受到影响(Min et al.,2006;Tanaka &Tanaka,2006)。已有研究表明,GABA处理减轻了胁迫引起的光合作用降低,恢复了叶绿体结构损伤(Xiang et al.,2016)。本试验发现外源CaCl2、GABA单独处理和复配处理均可以达到显著改善叶绿素荧光参数的效果,虽然复配处理组合SA+Ca6+GABA5仅在NPQ指标上与其他外源添加处理有显著差异,但其参数在各处理间均为最优,说明GABA和CaCl2复配较单独处理可以有效提高番茄叶片光合能力,主要原因在于GABA和CaCl2均可缓解盐碱胁迫造成的离子毒害,维持良好的营养元素吸收,从而维持细胞氧化还原平衡,促进叶绿素的正常合成。
活性氧(ROS)是调节植物生长、代谢和应激反应的重要信号分子(Baxter et al.,2014;Liu et al.,2018)。逆境引发的过多活性氧产生与细胞膜损伤和电解质渗漏密切相关(Demidchik et al.,2014)。外源GABA处理能有效地缓解盐碱胁迫引起的生长限制和膜脂氧化(Jin et al.,2019),但还没有直接证据表明GABA直接清除ROS来缓解胁迫伤害。因此猜测GABA增强胁迫耐受力可能是通过抗氧化系统介导的。本试验结果表明,外源GABA可明显降低盐碱胁迫下叶片H2O2含量和产生速率。此外,本试验更加关注的是GABA和CaCl2的复配效果,其中SA+Ca6+GABA5处理的H2O2含量和产生速率最低,说明CaCl2的添加可进一步增强GABA对番茄叶片清除活性氧能力的提高。GABA诱导植物逆境耐性增强与Ca2+调节通路密切相关(Yu et al.,2014)。与盐胁迫不同的是,盐碱胁迫造成的高pH环境会加剧根系离子吸收的障碍,迫使胞内环境酸化,胁迫条件下胞质中Ca2+/CaM和H+浓度上升,均可促进GAD的活性使GABA的含量增加(Aurisano et al.,1995)。这也可能是复配处理效果好于单独处理的原因。
本试验结果表明,与单独添加处理相比,复配处理SA+Ca6+GABA5对于盐碱胁迫伤害的缓解效果更好,说明外源CaCl2的添加对于GABA增强植物耐盐碱能力有促进作用,但其对于盐碱胁迫伤害的缓解效应并没有显著高于其他复配处理和单独处理,因此认为复配处理表现的是协同作用而并不是简单的叠加关系,是代谢调控的结果。CaCl2可能通过代谢调控GABA来诱导植物提高耐盐碱能力。因此,外源GABA和CaCl2在抗逆过程中如何影响内源GABA和Ca2+的变化,从而更好地发挥协同效应还有待进一步明确,此外,本试验结果还为GABA与钙复配制剂在盐碱土生产上的应用提供了参考。
