陆地棉种子和纤维中非结构性碳水化合物含量的基因型差异及与衣分形成的关系
2021-07-22向春玲罗海华彭金剑袁长凯汤飞宇
向春玲,陈 功,罗海华,彭金剑,高 欣,袁长凯,汤飞宇
(江西农业大学 农学院/作物生理生态与遗传育种教育部重点实验室,江西 南昌 330045)
【研究意义】衣分是指子棉产量中纤维产量所占的百分比,是评价棉花品种优劣的关键指标,研究影响衣分形成的生理生态因素具有重要的意义。子棉纤维主要的成分是纤维素,比例约占90%或更高[1]。纤维素是一种结构性碳水化合物,最初的合成底物是蔗糖[1]。种皮的主要成分是淀粉,种胚则以脂肪和蛋白为主[2]。脂肪和蛋白的合成均以碳代谢为基础,依靠碳代谢的中间产物提供碳骨架和能量[3]。因此,棉铃(纤维、种胚和种皮)的发育受到碳代谢的严格控制。衣分的高低主要取决于碳同化物在棉花种子及其着生纤维之间的分配[4-5]。【前人研究进展】棉铃内的非结构性碳水化合物主要包括葡萄糖、果糖、蔗糖及淀粉等,对棉铃的生长发育具有重要的作用。其作用机制体现在以下4个方面:(1)作为底物用于细胞结构或贮藏物质的合成。如纤维细胞纤维素合成的最初底物是蔗糖,直接前体是尿苷二磷酸-D-葡萄糖(UDPG)[6-9];(2)维持细胞生理生化活动所需能量的来源,如己糖通过糖酵解产生ATP实现棉花胚珠转移细胞的物质运输功能[10];(3)作为重要的渗透调节活性物质,调控细胞的渗透势和膨压,影响细胞的伸长和扩展,如提高纤维内的己糖浓度促进了纤维的伸长[11];(4)作为信号分子调节相关基因或转录因子的表达,影响细胞的生长发育。如葡萄糖能够上调细胞周期基因的表达,刺激早期幼胚的细胞分裂[12]。蔗糖上调与贮藏相关基因的表达,启动细胞的分化与成熟[13]。棉纤维的发育和品质形成与非结构性碳水化合物的动态变化息息相关[14-15]。蔗糖和可溶性糖转化率高的棉纤维,纤维素的累积速率较高,快速累积的持续时间长,对棉纤维干物质量的增加有利[14]。可溶性糖、蔗糖和β-1,3 葡聚糖含量的动态变化与棉纤维素累积特性相关,进而影响到棉纤维强度的形成[15]。可溶性糖作为棉纤维内一类主要的渗透调节物质,是驱动棉纤维伸长的重要动力[1,16]。较长纤维的海岛棉比陆地棉在棉铃开花后10 d 具有更高的葡萄糖、果糖及蔗糖浓度[11]。在干旱条件下,干旱钝感基因型比敏感基因型在纤维伸长阶段具有更高的己糖和蔗糖水平,可能是其最终纤维长度下降幅度较轻的重要原因[16]。棉花种胚的发育受到可溶性糖的种类、数量及相互比例等的影响。如在胚珠发育早期(棉铃开花后5 d),在种皮细胞内检测到细胞壁酸性转化酶基因(GhCWIN1)转录活跃,该酶催化蔗糖分解产生的葡萄糖有利于细胞核分裂,增加种胚细胞数量[17]。在棉花种胚发育过程中,己糖浓度一直呈下降趋势,在中后期种胚维持较高的蔗糖浓度,成为主要的非结构性碳水化合物[18],为种胚贮藏物质棉子糖和水苏糖的合成提供底物[19]。高浓度的蔗糖可激活细胞分化和物质贮藏功能[13]。在蚕豆中发现,蔗糖水平上升时,也正是己糖水平明显下降,细胞分裂中止,细胞启动分化及贮藏活性[20]。在种胚发育初期活跃的细胞分裂阶段往往表现出高的己糖/蔗糖浓度比,而物质贮藏阶段则表现高的蔗糖/己糖浓度比[21]。因此葡萄糖和蔗糖除了作为营养物质外,还分别代表了诱导种胚细胞分裂和分化、贮藏相关的信号[13,21]。适度晚播的棉花种胚花后5 d 表现出更强的酸性转化酶活性,产生更高的己糖浓度,促进了种胚细胞分裂,因而导致了更大的棉子[22]。【本研究切入点】源叶光合生产的蔗糖在棉铃胚珠种皮内的维管束卸载后,一部分向外以共质体运输的方式输入纤维,另一部分以质外体运输的方式输入种胚[1]。其中分布于内种皮最内侧的转移细胞在促进碳同化物向种胚的转移发挥了关键作用[10]。由此可见,纤维和种胚对碳同化物的竞争可能影响两者干物质量的积累,进而影响衣分的形成。关于纤维和胚珠(种胚和种皮)内非结构性碳水化合物的变化状况与衣分形成的关系尚未见报道。【拟解决的关键问题】研究以2个衣分和棉子大小具有明显差异的陆地棉品系为试验材料,观察其棉铃发育过程中纤维、种胚和种皮非结构性碳水化合物(己糖、蔗糖和淀粉)含量的变化,比较不同基因型的差异,证明非结构性碳水化合物在纤维和种胚分配的差异可能是导致衣分差异形成的一个重要原因。旨在为培育高衣分棉花品种提供参考依据和技术思路。
1 材料与方法
1.1 试验材料与设计
试验材料为江西农业大学自育陆地棉品系:低衣分大子品系A201 和高衣分小子品系A705。于2016 年种植于江西农业大学农业科技园,供试土壤为红壤旱地,前茬空闲,肥力中上等。田间试验随机区组排列,4 行区,行长7.7 m,行距1.0 m,株距0.3 m,小区面积30.8 m2,3 次重复。2016 年5 月6 日播种。全生育期施纯N:276 kg/hm2、P2O5:105 kg/hm2、K2O:247.5 kg/hm2。磷肥和钾肥作基肥和花铃肥分2 次施用,比例为1∶1;氮肥作基肥、花铃肥和盖顶肥分3 次施用,比例为1∶2∶1。按常规高产栽培技术要求管理,及时灌溉、防治病虫害和杂草。
1.2 取样
于盛花期挂牌标记中部果枝(7—9)内围果节当天白花,开花后5,10,17,24,31,38,45 d 共7 个时期取发育整齐一致的棉铃带回实验室,在室内将种子、种胚和种皮分离开来。种胚与种皮的分离始于开花后24 d,由于此前的种胚尚小,难以分离。样品经105 ℃杀青30 min 后于60 ℃下烘干至恒重,烘干后用微型植物粉碎机磨碎并过32目筛网,用于己糖(葡萄糖、果糖)、蔗糖和淀粉浓度的测定。
1.3 测定项目与方法
葡萄糖、果糖、蔗糖和淀粉的提取方法参照陈功等[23]论文中所述。葡萄糖和果糖浓度的测定采用酶比色法[24],己糖浓度定义为葡萄糖和果糖浓度的总和,蔗糖浓度的测定采用间苯二酚法[25];淀粉浓度的测定采用蒽酮法[25]。
1.4 数据统计分析
采用独立样本T测验,检验A705和A201棉铃产量性状、各取样点己糖、蔗糖及淀粉浓度的差异显著性。使用Origin 8.5软件作图。
2 结果与分析
2.1 棉铃产量性状的基因型差异
由表1 可知,相对于A201,A705 具有显著更大的单铃纤维质量、衣指和衣分,显著更小的铃质量、单铃种子质量、子指。表明A201 具有更强的种子干物质积累能力,A705 则具有更高的纤维干物质积累能力。

表1 A705和A201棉铃产量性状的表现Tab.1 Performance of cotton boll yield traits in A705 and A201
2.2 A705和A201纤维非结构性碳水化合物浓度的动态变化与差异
由图1A 可知,A705 和A201 棉铃开花后,纤维蔗糖浓度的变化一致,皆呈逐渐下降的趋势。除铃龄45 d外,其余时期A705的蔗糖浓度都显著或极显著高于A201,前者比A201平均高134.3%。纤维己糖浓度基本呈下降趋势(图1B)。A705 纤维己糖浓度比A201 平均高21.3%,且在铃龄38 d 显著高于A201。综上所述,高衣分小子品系A705比低衣分大子品系A201纤维具有较高的蔗糖、己糖水平。
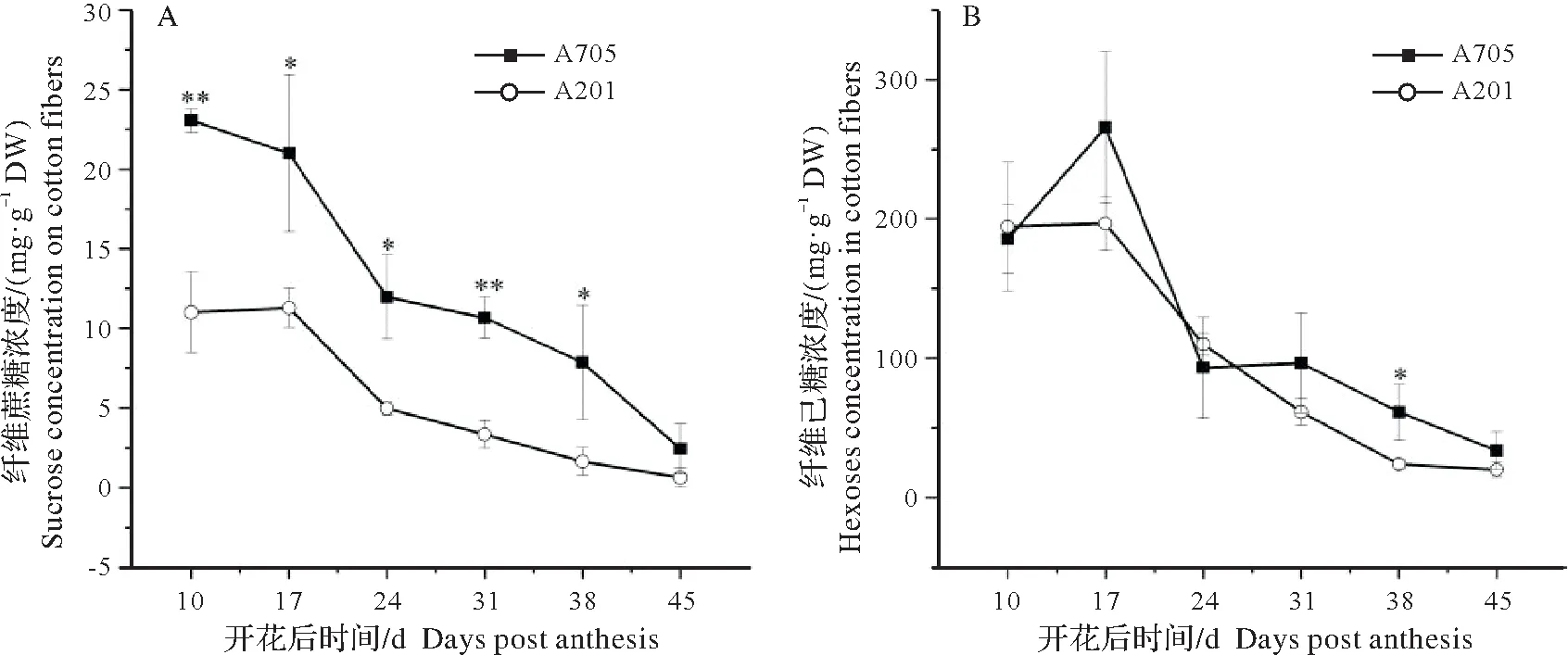
图1 棉纤维蔗糖浓度(A)和己糖浓度(B)的变化趋势Fig.1 Dynamics of sucrose(A)and hexose concentrations in cotton fibers
2.3 A705和A201胚珠/种胚非结构性碳水化合物浓度的动态变化与差异
由于铃龄17 d前(含17 d)的种胚还比较细嫩,尚不能从胚珠中分离开来,因此,图2中铃龄5~17 d代表的是胚珠的非结构性碳水化合物状况,24~45 d 代表的是种胚的非结构性碳水化合物状况。由图2A可知,A705与A201的种胚中蔗糖浓度的变化趋势一致,自开花后5~17 d呈上升趋势,24 d略有下降,31 d达到峰值,此后呈下降趋势。A201 种胚蔗糖浓度整体高于A705,平均高27.11%,且在铃龄5 d,31~45 d显著高于后者。2 个品系种胚的己糖浓度均呈下降趋势(图2B)。A201 平均水平比A705 高29.17%,且在铃龄5,10,24,38 d 显著或极显著高于后者。2 个品系种胚的淀粉浓度均呈单峰曲线,自铃龄5 d 起持续上升,至24 d达到峰值,此后下降(图2C)。A201平均水平比A705低18.56%,且在铃龄10 d,45 d显著低于后者。综上所述,低衣分大子品系A201比高衣分小子品系A705种胚具有较高的蔗糖和己糖水平,较低的淀粉水平。

图2 棉花胚珠/种胚蔗糖(A)、己糖(B)、淀粉(C)浓度变化趋势Fig.2 Dynamics of sucrose(A),hexose(B)and starch(C)concentrations in cotton ovules/embryos
2.4 A705和A201种皮非结构性碳水化合物浓度的动态变化与差异
图3 表明,2 个品系种皮的蔗糖、己糖和淀粉浓度均随棉铃发育均呈下降趋势。A201 种皮蔗糖的平均浓度比A705 高24.24%,且在铃龄31 d、38 d 均显著高于A705(图3A)。A201 种皮己糖的平均浓度比A705 高26.52%,且在铃龄24 d、31 d 均极显著高于A705(图3B)。与蔗糖和己糖的表现相反,A201 种皮淀粉的平均浓度比A705 低18.56%,且在铃龄31 d 极显著低于A705。综上所述,低衣分大子品系A201比高衣分小子品系A705种皮具有较高的蔗糖和己糖水平,较低的淀粉水平。

图3 棉花种皮蔗糖(A)、己糖(B)、淀粉(C)浓度变化趋势Fig.3 Dynamics of sucrose(A),hexose(B)and starch(C)concentrations in cotton seed coats
3 结论与讨论
3.1 非结构性碳水化合物与棉花纤维素合成的关系
纤维中的主要成分是纤维素,约占90%或更高[1]。蔗糖是纤维素合成的最初底物。蔗糖和可溶性糖转化率高的棉纤维,纤维素的累积速率较高,快速累积的持续时间长,对棉纤维干物质量的增加有利[14]。高强棉纤维相对于中、低强纤维次生壁加厚发育期(铃期24~31 d)蔗糖累积量高[15]。研究发现高衣分小子基因型A705 纤维发育期间比低衣分大子基因型A201 具有更高的蔗糖浓度,因而为纤维素合成提供了更丰富的原料,可能是A705比A201具有更高单铃纤维质量、衣指和衣分的重要原因。己糖可以通过多种生化途径转化为纤维素合成的直接前体UDPG,用于纤维素的合成。因此A705 纤维内较高的可溶性糖水平对其纤维素合成有利。
3.2 非结构性碳水化合物与棉花种子发育的关系
棉花胚珠发育初期包含相当多的己糖,而蔗糖较少[19],这与本研究的结果一致。在本研究中,由于铃龄5 d 的胚珠比较幼嫩,胚乳呈乳汁状,细胞壁尚未形成,种胚处于球形胚阶段,肉眼尚不能观察到,还不能从胚珠中分离出来。在多个物种如拟南芥、蚕豆和棉花,发现葡萄糖浓度和细胞壁转化酶表达与早期胚(心形胚之前)有丝分裂活性存在显著的相关性,表明葡萄糖可作为一个信号刺激胚胎发育过程中的细胞分裂[21]。有报道称库器官如玉米子粒和豆类种子中的己糖可刺激细胞分裂,可能是通过调控细胞周期基因如CDC2,Cyclin delta2 和3 实现[26]。在本研究中,低衣分大子基因型A201 胚珠在开
花后5~10 d比高衣分小子基因型A705具有显著更高的己糖浓度,高浓度的己糖可能促进了种胚的细胞分裂,增加了种胚细胞的数量。蚕豆幼嫩种胚的己糖浓度由种皮的酸性转化酶控制,大子基因型种皮中酸性转化酶高活性持续时间长,分解产生的大量己糖通过质外体途径释放到胚乳中,促进种胚较长时间的分裂,因而产生比小子基因型具有更多的细胞数目[20]。笔者在另一项研究中发现,适度晚播的棉花种胚花后5 d表现出更强的酸性转化酶活性,产生更高的己糖浓度,促进了种胚细胞分裂,因而导致了更大的棉子[22]。
种胚蔗糖含量从花后一周直至20 d,快速升高,然后在种子成熟阶段,保持基本稳定[19]。这与本研究略有不同,棉铃开花至花后31 d,胚珠/种胚蔗糖一直呈上升,但此后下降,表明蔗糖有较好的转化利用率,对种胚合成脂肪、蛋白质、棉子糖和水苏糖等有利。高浓度的蔗糖作为信号促进种胚从细胞增殖阶段转向物质积累阶段[12-13],另一方面为种胚贮藏物质的合成提供碳骨架。低衣分大子基因型A201 比高衣分小子基因型A705 胚珠/种胚发育阶段具有较高的蔗糖水平可能是导致其单铃种子质量和子指更大的又一原因,这与Luo等[5]的研究结果一致。棉花种胚在花后20 d开始积累淀粉而同期种皮中的淀粉浓度下降[27],而本研究发现种胚和种皮的淀粉浓度均在花后24 d达到峰值,此后下降。在花后45 d,种胚中的淀粉基本消失,取而代之的是等量的半乳糖苷类物质,如棉子糖和水苏糖[27]。A201 比A705 种皮具有较高的蔗糖和己糖水平。种皮中的蔗糖和己糖既可以通过共质体途径向纤维输送,也可以通过质外体途径向种胚运输,其中精准的控制机制还不清楚[1]。现有证据表明内种皮最内侧的转移细胞促进了质外体途径的运输[10],因此种皮内较高水平的蔗糖和己糖对于种胚的发育可能是有利的。
蔗糖合酶是反映棉花种子和纤维库强度一个重要的生化标记,酶活性高意味着库器官贮藏物质合成活性强[2,28]。本研究进一步认为作为蔗糖合酶催化作用的底物蔗糖同样可以反映种子和纤维物质合成能力的一个标记。纤维干物质积累能力强的A705纤维具有较高的蔗糖浓度,而种子干物质合成能力强的A201 种子具有较高的蔗糖浓度。A705 纤维具有较高的蔗糖浓度而种子(种胚和种皮)具有较低的蔗糖浓度,可能是其衣分较高的重要原因。综上所述,在棉铃发育过程中,高衣分小子基因型A705纤维具有较高的蔗糖和己糖水平,而种子中较低;反之低衣分小子基因型A201 种子(种胚和种皮)具有较高的蔗糖和己糖水平,而纤维中较低。纤维中较高而同时种子中较低的蔗糖和己糖水平可能是导致A705较高衣分的重要原因。
