幼龄红豆树生长和形质性状家系变异分析
2021-07-11肖德卿邓章文罗芊芊徐肇友陈焕伟徐洪辉周志春
肖德卿,邓章文,罗芊芊,徐肇友,沈 斌,陈焕伟,徐洪辉,周志春*
(1. 中国林业科学研究院亚热带林业研究所,浙江省林木育种技术研究重点实验室,浙江 杭州 311400;2. 江西省抚州市林业科学研究所,江西 抚州 344114;3. 浙江省龙泉市林业科学研究院,浙江 龙泉 323700)
植物对环境的适应和自身遗传结构在适应环境过程中的改变可通过生长和形质性状表达出来[1],通过对植物生长和形质性状变异进行研究,可掌握其遗传变异规律,提升其遗传改良潜力[2]。同时,充分了解生长和形质性状的变异有助于准确地评估改良的可靠性[3]。植物的生长和形质性状变异越丰富,越能体现其对环境的适应[1,4],而开展多点区域试验可为不同立地和地区选出具有稳定遗传性状的优良家系。辛娜娜等[5]从福建建瓯和江西永丰两地点分别初选出24个和17个木荷(Schima superbaGardn.et Champ.)早期速生优良家系。洪舟等[6]在分析降香黄檀(Dalbergia odoriferaT. Chen)生长性状遗传变异的基础上,根据育种值筛选出10个速生且稳定的优良家系,并进一步筛选出12株可作为无性系种子园材料的优良单株。陈孝丑[7]通过对枫香(Liquidambar formosanaHance)生长和材性的家系遗传变异规律研究,筛选出9个速生优良家系、28株优良单株以及7个可推广的生长材性兼优家系。林木遗传改良中进行早期优良家系和优良单株的选择,在缩短林木育种周期的同时,可有效地提高用材收益。
红豆树(Ormosia hosieiHemsl. et E.H.Wilson)是豆科(Fabaceae)红豆属(OrmosiaJasks.)半常绿或落叶大乔木,自然分布于浙江、江西、福建、湖北、重庆和四川等省(市),是国家二级重点保护野生植物[8],其木材质地坚硬,花纹别致,是上等的雕刻和装饰等珍贵用材。但红豆树因秋梢生长而时常受冻害,导致顶芽枯死,一些苗木在造林后第2年会形成2个侧枝替代顶梢生长,或形成1个与顶梢竞争的侧梢,因此红豆树人工林树干通直度较低,易形成多杈干等缺陷从而影响经济价值[9-12]。现阶段红豆树研究的重点是在优良种质资源收集和保护的基础上,系统分析其生长和形质性状的变异规律,加强其优良品种选育,改良其生长和形质指标,为红豆树种子园的建立及无性快繁提供优良种质材料的同时,也为区域化推广提供理论依据。李峰卿等[10]对不同产地的红豆树家系种子和幼苗性状变异进行研究,以苗高和地径为筛选指标选出浙江龙泉生长表现优异的19个红豆树优良家系。本研究通过对浙江龙泉和江西抚州两个试验点的3年生红豆树家系试验林进行全林测定,系统分析其生长和形质性状的家系遗传变异规律及性状间的相关关系等,以期为红豆树的遗传改良和品种选育提供参考依据。
1 材料与方法
1.1 材料来源与试验地概况
2015年10月至11月分别收集来自浙江、江西、福建和四川4个省份的76株红豆树优树自由授粉种子,2016年5月于浙江省龙泉市林业科学研究院省级保障性苗圃进行育苗[3]。2017年利用培育的1年生家系裸根苗分别在浙江龙泉和江西抚州两地点营建红豆树优树家系测定林。浙江龙泉(119°07′ E,27°59′ N)和江西抚州(116°21′ E,27°57′ N)均属于中亚热带季风气候区,浙江龙泉年均气温17.6 ℃,年均降水量为1 699.4 mm,无霜期为263 d;江西抚州年均气温17.8 ℃,年均降水量为1 600 mm,无霜期为275 d。两试验地点的土壤均为酸性红壤,土层在80 cm以上,肥力中等。
1.2 试验设计与测定方法
浙江龙泉和江西抚州两地点均采用完全随机区组设计,株行距2.0 m × 2.5 m,其中浙江龙泉为5次重复,8株单列小区,江西抚州为7次重复,5株单列小区。浙江龙泉参试家系为76个,江西抚州参试家系为71个,两试验点共有家系69个。于2019年11月底对两地点红豆树优树家系试验林进行全林生长和形质性状调查,生长性状包括树高、地径和冠幅,形质性状包括当年抽梢长、一级分枝数、最大分枝角、最大分枝长、最粗分枝基径和树干通直度。其中,树干通直度分为通直(5分)、较通直(4分)、一般(3分)、弯曲(2分)和严重弯曲(1分)5个级别。
1.3 数据处理
数据处理前依照拉依达准则[13]剔除异常数据,以单株测定值为单元进行性状方差分析,一级分枝数及树干通直度经X1/2数据转换,最大分枝角经反正弦转换,最粗分枝基径经1/X转换。
单点方差分析的统计模型为:Yijk=u+Bi+Fi+BFij+Eijk
式中,Yijk为单点试验第i个区组第j个家系第k个单株观测值,u为群体平均效应,Bi为第i个区组效应,Fj为第j家系的效应,BFij为第i家系和第j区组的互作效应,Eijk为机误。

采用SAS 8.0软件的GLM和VARCOMP等过程进行性状方差分析,估算方差分量和表型相关系数,进而估算家系遗传力、单株遗传力和性状间遗传相关系数。其中,单点方差分析、家系遗传力、单株遗传力、表型相关系数、遗传相关系数、加性遗传方差和加性遗传协方差等计算公式见参考文献[14]。
2 结果与分析
2.1 红豆树生长和形质性状家系遗传变异
单点方差分析结果表明(表1),浙江龙泉和江西抚州两地点的3年生红豆树生长和形质性状都存在极显著家系差异。浙江龙泉点3年生家系树高、地径和当年抽梢长的变幅分别为0.94~2.02 m、1.67~3.07 cm和0.22~0.49 m,树高、地径和当年抽梢长最大家系分别为最小家系的2.15、1.84和2.23倍。在江西抚州点,树高、地径和当年抽梢长的变幅分别为1.01~2.05 m、1.93~3.53 cm和0.34~0.95 m,树高、地径和当年抽梢长最大家系分别为最小家系的2.03、1.83和2.79倍。与生长性状一样,红豆树的一级分枝数、最大分枝角、最大分枝长和树干通直度等在家系间也均达到极显著的水平,变异系数为23.3%~65.1%,如浙江龙泉和江西抚州两地点的红豆树家系最粗分枝基径变幅分别为0.62~1.28 cm和0.65~1.45 cm,这为早期速生和分枝习性优良的红豆树家系选择提供了较大潜力。

表1 红豆树生长和形质性状的单点方差分析Table 1 Single point analysis of variance of growth and form-quality traits of O. hosiei
2.2 红豆树生长和形质性状的遗传力估算
红豆树生长和形质性状的遗传力估算值表明(表2),除树干通直度外,其余性状在浙江龙泉和江西抚州两地点的家系遗传力估算值均较高,其变幅为0.36~0.83。如浙江龙泉点树高和地径的家系遗传力估算值分别为0.83和0.73,江西抚州点树高和地径的家系遗传力估算值均为0.70,意味着红豆树家系幼林生长受中等至偏强的家系遗传控制。与家系遗传力相比,仅浙江龙泉点红豆树树高单株遗传力估算值(0.87)高于家系遗传力估算值(0.83),受偏强的遗传控制,两地点红豆树其余性状的单株遗传力估算值均低于家系遗传力估算值,受中等至偏弱的遗传控制,其变幅为0.04~0.61。
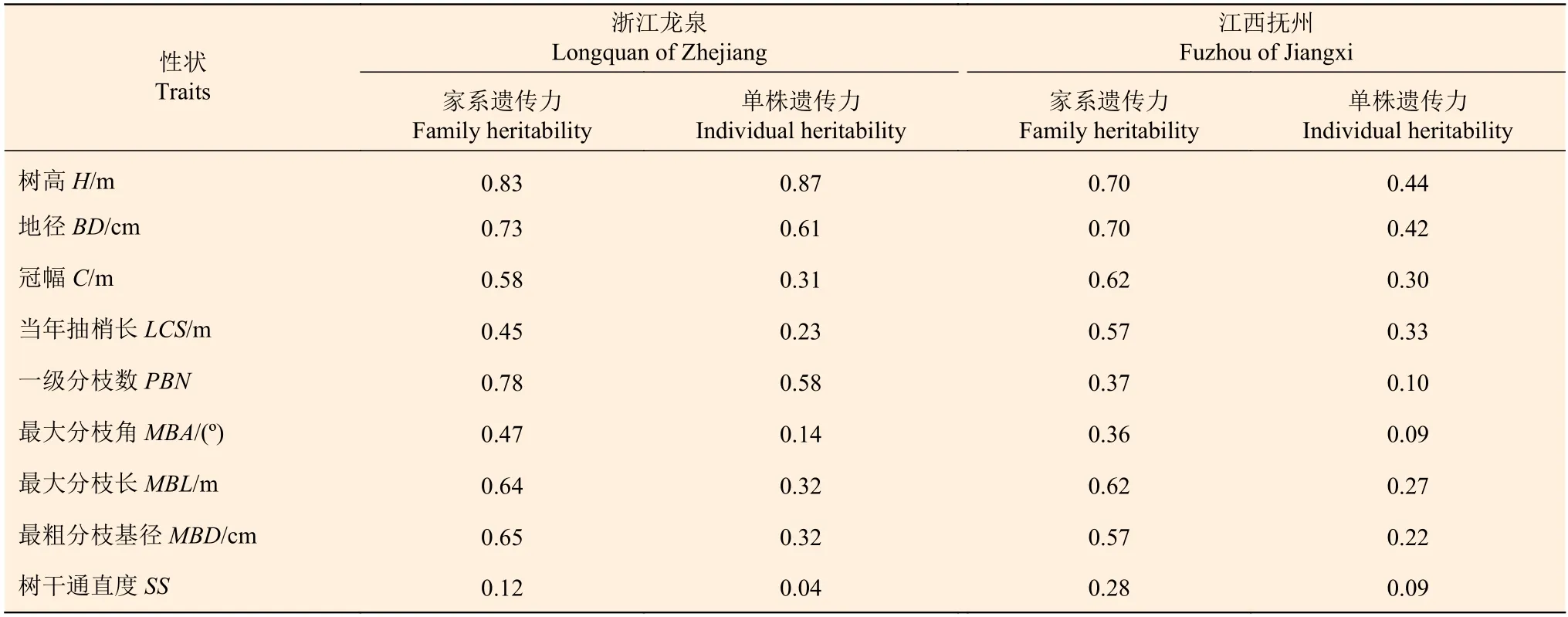
表2 红豆树生长性状和形质性状的遗传力估算值Table 2 Estimated of heritability of growth and form-quality traits of O. hosiei
2.3 红豆树生长和形质性状的遗传和表型相关
从表3可以看出,浙江龙泉和江西抚州两地点红豆树家系性状间遗传相关系数总体上大于表型相关系数。3年生红豆树家系树高、地径和冠幅间均呈极显著的遗传正相关(rg= 0.344~0.728),树高、地径与一级分枝数、最粗分枝基径间也呈极显著的遗传正相关(rg= 0.351~0.715),这表明红豆树家系树高与地径的生长量越大,其一级分枝数越多且分枝越粗。在浙江龙泉点,最大分枝角与树高、冠幅、当年抽梢长、最大分枝长和最粗分枝基径等性状的遗传相关程度较弱(rg= -0.017~0.183),为选择速生性较好、分枝角度较大和分枝较细的品系提供了可能。总体来看,红豆树家系生长性状与形质性状间普遍呈显著或极显著正相关,为培育速生优质干材,应加强对红豆树家系早期修枝和选育。
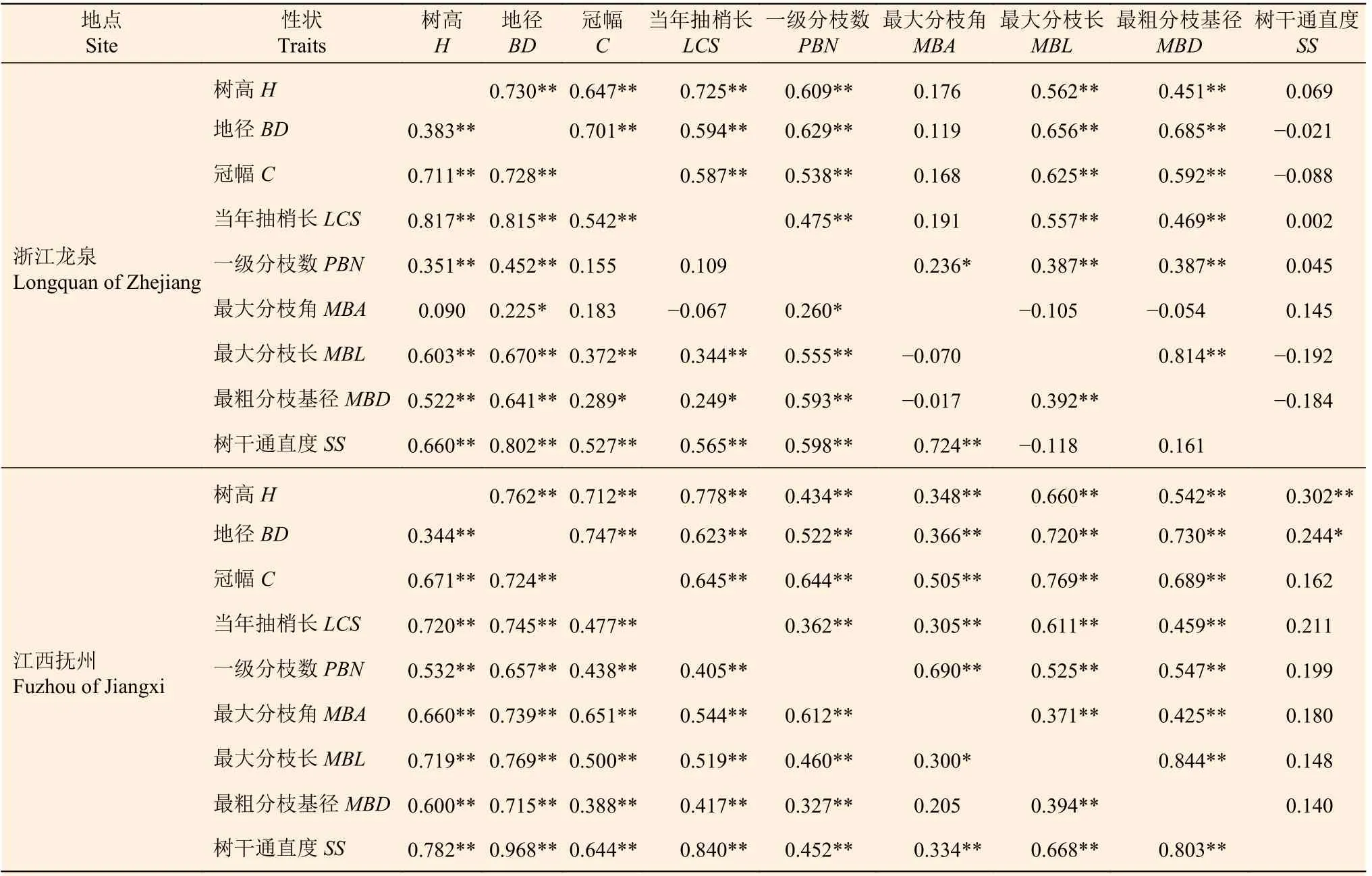
表3 两试验点红豆树家系生长和形质性状间的遗传与表型相关系数Table 3 Genetic and phenotypic correlation coefficients between growth and form-quality traits of O. hosiei
3 讨论
遗传与变异是林木遗传育种的基础[15],本研究在前期红豆树优树种子和种苗性状变异研究的基础上[10],探究浙江龙泉和江西抚州两地点3年生红豆树家系生长和形质性状的遗传变异规律。单点方差分析结果表明,两地点的红豆树生长和形质性状均存在极显著的家系变异,各性状的变异系数均大于20.0%,幼苗期也曾发现红豆树家系苗高和地径存在显著性差异[10],这为速生优质家系的选择提供了理论依据和物质基础。红豆树易形成多杈干,植株的分枝数较多会降低树干通直度,影响了优质干材的形成[16],因此选择分枝数少且细的家系选育更具意义。与江西抚州点相比,浙江龙泉点树高、冠幅和一级分枝数均较大,分枝基径较细,树干通直度较低。究其原因,可能是浙江龙泉点比江西抚州点降水稍多。水分为植物制造碳水化合物、维持细胞质水合作用所必需,又是植物养分的运输工具,降水的增加促进其树高生长、冠幅伸展和分枝数增多。这说明通过加强水肥管理促进红豆树生长的同时,应重视红豆树早期的修剪除蘖,以利于速生通直干材的形成。
3年生红豆树生长和形质性状的家系遗传力普遍保持着较高水平,受中等至偏强的的家系遗传控制,有利于优良家系的选择。两地点各性状的单株遗传力普遍低于家系遗传力,受中等至偏弱的遗传控制,浙江龙泉点的家系遗传力和单株遗传力大体高于江西抚州点,可能与浙江龙泉点的立地条件有关,重复间和重复内立地环境相对一致会显著降低试验的环境效应。研究中还发现,浙江龙泉点家系树高的单株遗传力大于家系遗传力,这可能是因为红豆树自由授粉家系试验群体中包含部分自交或近交的后代[17],且性状遗传力的估算值与研究材料、家系数量和试验地环境等因素也有关。总体来说,红豆树生长和形质性状具有较大的遗传改良潜力,这为其优良家系和单株的选择提供了可能。
分析生物性状之间的相关关系可为优质家系的选择提供帮助[18]。遗传相关比表型相关更能准确反映由遗传效应引起的性状间的相关变化[19]。浙江龙泉和江西抚州两地点红豆树家系地径与树干通直度的遗传相关均较高,在江西抚州点,两个性状的遗传相关系数甚至接近于1,表明通过对地径的选育可以选出树干通直度较高的植株,在同一群体中,可同时实现这两个性状的提高[20]。浙江龙泉点的3年生红豆树优树家系最大分枝角与树高及最粗分枝基径等其它生长和形质性状相关性弱,在遗传上较为相互独立。辛娜娜等[5]和罗芊芊等[18]在木荷和南方红豆杉家系研究中也发现其最大分枝角与最粗分枝基径不相关或弱相关的规律,这有利于分枝细、分枝角大即易自然整枝家系的选择。两地点的红豆树家系树高、地径与一级分枝数和最粗分枝基径的相关性较强,说明树高和地径生长量突出的红豆树家系,其分枝数多,分枝较粗,在生产上可通过一级分枝数来间接选择速生的红豆树,并通过早期修枝和合理密植来培育优质干材。
限于林木生长周期较长,现阶段试验林处于幼龄阶段,仅对红豆树家系的生长和形质性状进行初步分析,今后还需继续对其生长进行跟踪调查,并结合多区域测定来进一步验证红豆树家系的适应性和稳定性,为不同区域和立地筛选出产量较高且通直度较好的优质家系和单株。
4 结论
幼龄红豆树生长和形质性状存在极显著的家系遗传差异,变异幅度较大,多数性状受中等至偏强家系遗传控制,且各性状间的相关性差异也较大,树高、地径与一级分枝数、最粗分枝基径的相关性较强,有利于选育速生且通直度较高的红豆树优良家系和单株。幼龄红豆树生长和形质性状家系变异的研究结果,可为红豆树种质资源利用和遗传改良提供参考依据。
