The Variation of Microbial (Methanotroph) Communities in Marine Sediments Due to Aerobic Oxidation of Hydrocarbons
2021-06-25LIJingLIUChanglingWUNengyouHEXingliangHAOXiluoLIFengCHENYeandMENGQingguo
LI Jing, LIU Changling, *, WU Nengyou, HE Xingliang, HAO Xiluo, LI Feng, CHEN Ye, and MENG Qingguo
The Variation of Microbial (Methanotroph) Communities in Marine Sediments Due to Aerobic Oxidation of Hydrocarbons
LI Jing1), 2), LIU Changling1), 2), *, WU Nengyou1), 2), HE Xingliang1), HAO Xiluo1), 2), LI Feng1), CHEN Ye3), and MENG Qingguo1), 2)
1),,,266071,2),,266071,3),,266100,
Methanotrophs in marine sediments and overlying water attenuate the emissions of methane into the atmosphere and thus play an important role for the global cycle of this greenhouse gas. However, gas released from natural hydrocarbon seeps are not pure methane but commonly mixed hydrocarbons. Currently, how methanotrophic bacteria behave in the co-presence of methane and heavier hydrocarbons remains unknown. In this paper, the bacteria were cultured aerobically in fresh sediment samples (collected from Bohai Bay in eastern China) at 28℃ under the atmospheres of pure methane and methane+ethane+propane mixed gas, respectively. The prevailing terrigenous-alkanes and fatty acids in the original sediment samples varied consistently after incubations, confirming the proceeding of aerobic bacterial activities. The real-time quantitative PCR assay and sequencing of theand particulate methane monooxygenase () genes revealed the changes of microbe communities to a methanotroph-dominating structure after incubations. Particularly, after incubations the family Methylococcaceae (type I methanotrophs) became dominant with proportions higher than 40%, whereas Methylocystaceae (type II methanotrophs) nearly disappeared in all incubated samples. Moreover, the species of methanotrophs from the samples treated with pure methane were dominated by, whereastook the predominant proportion in the samples treated with mixed gas. The phenomenon suggests that some methanotrophs may also utilize ethane and propane. Collectively, this study may help to gain a better understanding of the effects and contributions of microbial activities in marine hydrocarbon seep ecosystems.
methanotroph; aerobic oxidation of hydrocarbons; sediment incubation; lipid biomarker; microbial community
1 Introduction
Ocean plays a significant role in global climate change since it is the largest carbon reservoir on the earth and hosts substantial hydrocarbons. These hydrocarbons (particularly methane) are recognized as ‘greenhouse’ gases and can absorb infrared radiation much more strongly than carbon dioxide, although their concentrations are two to six orders of magnitude lower than that of carbon dioxide in atmosphere (Lashof and Ahuja, 1990). Marine hydrocar-bons can occur as dissolved and free gases in water and sediments, as well as crystalline solids (natural gas hydra- tes). Every year the leakage of oil and gas and decomposition of gas hydrates from their submarine reservoirs release substantial volumes of hydrocarbons (Etiope, 2015). Nonetheless, much of them may never reach the atmosphere. This is attributed to the submarine microbial com-munity which consumes more than 90% of the generated hydrocarbonsanaerobic/aerobic oxidation processes (Valentine., 2001; Hinrichs and Boetius, 2002; Reeburgh, 2007). However, the microbial populations that con-tribute to such processes in submarine environments are still not well illustrated (Redmond., 2010).
Compared to microbial anaerobic oxidation (Wang., 2014), aerobic oxidation is more common and widespread in marine systems since oxygen is still abundant in the deep-sea below 3000km (Boetius and Wenzhöfer, 2013). Some studies have been specially conducted to examine the features of microbes related to the aerobic oxidation of me-thane (., Holmes., 1995; Dingand Valentine, 2008; Valentine, 2011; Cui., 2018). The known methanotrophic bacteria are separated into two assemblages,., γ-proteobacteria (type I) and α-proteobacteria (type II) (Han-son and Hanson, 1996) and the key enzyme that they rely on for methane oxidation, methane monooxygenase (MMO), is found in two forms. The particulate MMO (pMMO) is associated with intracellular membranes and has been found in almost all described methanotrophs, whereas the soluble MMO (sMMO) is free in the cytoplasm and is only found in few genera (Inagaki., 2004; Op den Camp., 2009; Redmond., 2010). Although MMO is generally restricted to the oxidation of methane, recent studies found that it might also oxidize C2H6–C4H10alkanes, indicating that methanotrophic bacteria may notonly grow on methane but also on multi-carbon compounds such as ethane and propane (Hazeu and de Bruyn, 1980; Berthe-Corti and Fetzner, 2002; Kinnaman., 2007). However, in submarine environments how methanotrophic bacteria behave in the co-existence of methane and higher hydrocarbons is not well established. This limits the quan-titatively modelling of the microbial contributions in preventing marine hydrocarbon from entering atmosphere, as well as the understanding of the role of microbial community in global carbon circle.
In this study, we compared the aerobic microbial consumption of pure CH4with that of mixed gases by incubating marine sediment samples collected from Bohai Bay, eastern China, which is a prolific gas- and oil-bearing re- gion (and thus a natural seep of hydrocarbon gases). Specifically, we aim to characterize the assemblages of me- thanotrophic bacteria present in the natural seeps and determine how these microbial communities behave during the aerobic oxidation in the co-occurrence of methane and larger hydrocarbons. The results help to gain a better understanding of the effects and contributions of microbial activities in marine hydrocarbon seep ecosystems.
2 Materials and Methods
2.1 Sediment Sample
The marine sediment sample used in this study is the same as Li(2019). This sample is used because Li(2019) had demonstrated that it contains the microbial communities that can degrade hydrocarbons (including methane, ethane and propane)aerobic oxidation. Petrographically, this marine sediment sample is mainly composed of clay and was collected from the surface se- diment layer in the Shaleitian Coal Oil Point seep field in Bohai Bay, eastern China. The water depth of sampling po-sition is 22.7m andtemperature is 19℃. The com- position of natural gas emitted from this area is characte- rized by 93% methane, 3% ethane and 4% propane (Zhang, 2016). The sample was collected by a remotely operated submersible box sampler which weighed nearly 2.0kg. More details about the sediment sample and collection procedures can be found in Li. (2019).
2.2 Sediment Incubations
To illustrate the response of microbial community structure to aerobic oxidation, microbial aerobic oxidation ex- periments were conducted by using the sample mentioned above. The detailed procedures for sediment incubations were given by Li. (2019) and are summarized here. Artificial seawater was added to the sediment at a ratio of 1:1 (v/v) with a pH of 7.0. The sediment sample was stirredthoroughly and fifty milliliters of the sediment slurry were added into a 120mL serum bottle, which was then sealed with a butyl rubber septum and an aluminum crimp cap. The headspace of the bottle was subsequently purged with sterilized ultrahigh purity (UHP) N2and then O2and hydrocarbons were added. Both pure methane and mixed gases (methane:ethane:propane=79.96:10.04:10.00) were used, corresponding to incubation i and incubation ii, respectively. The ratio of gas mixtures was set according to the composition of hydrocarbon gases in marine seepages (methane: 62.30%–100.00%; ethane: 0–7.70%; pro- pane: 0–18.80%) (.,Milkov, 2005; Liu., 2015). The two incubations were performed with another one killed treatment sample as the control. The samples were incubated at 28℃ in the dark on an orbital shaker set at 120r min−1. The headspace gases were subsampled for compositional measurement and isotope analyses at intervals of 3–7 days to monitor the process of the aerobic oxidation re-action. Once hydrocarbons had completely been consumed, the slurry sediments (15mL) were sampled for lipid and DNA analyses.
2.3 Lipid Biomarker Analysis
The freeze-dried, homogenized and powdered samples (including the original and incubated samples) were extracted twice with a mixture of dichloromethane (DCM)/ MeOH (3:1, v/v) by using a Dionex Accelerated Solvent Extraction system (ASE 200, Dionex Inc) at 110℃. The volume of the extract was reduced under a stream of N2and then the extract was desulfurized by using copper sticks. After desulfurization, the total lipid extracts were saponified with 6% KOH in MeOH for 12h and then separated into different fractions through silica gel flash column chromatography. The neutral lipid fraction was driedunder a gentle N2stream and derivatized by using N, O-bis (trimethylsilyl)-trifluoroacetamide (BSTFA) in acetonitri- le to form TMS-ethers (Trimethylsilyl ethers) at 70℃ for 1h before instrumental measurements. Hexane was subsequently added to extract fatty acids following the addition of HCl (pH<2). Fatty acids were methylated with BF3-MeOH to form fatty acids methyl esters (FAMEs), then all lipids were analyzed by gas chromatography-massspectrometry (GC-MS; Agilent 7890A/5975C). Separation was achieved on a HP-5MS column (30m×0.25mm×0.25μm). The injector temperature was set at 290℃. We used He as a carrier gas at 1.0mLmin−1. For-alkane analysis, the initial column temperature of 80℃ was held for 1min, then programmed to 310℃ at 5℃min−1and held for 20min. The temperature programs for fatty acids were employed as follows: 60℃ for 1min, followed by 80–200℃ at 10℃min−1intervals, 200–250℃ at 5℃min−1intervals, and then 250–290℃ at 2℃min−1intervals and held at 290℃ for 10min. Internal standards (C24D50for neutral lipids and-C19-COOH for fatty acids) were addedto samples before GC-MS analysis to aid in quantification. Lipid concentrations were determined by ratios of the peak areas to those of the internal standards. Lipids were tentatively identified by comparing the retention times with that of the standard which periodically runs under same conditions, and by using the standard spectral library (NIST02L) and reported literature (Hu, 2006).
2.4 DNA Extraction
Total DNA was extracted from the original and incubated samples by using the EZNA® Soil DNA kit (Omega Bio-Tek, Inc., Norcross, GA, USA) according to the ma- nufacture’s recommended protocol. Genomic DNA was eluted with 100μL of elution buffer and its quality was verified by agarose gel electrophoresis. A negative control reaction (using DNA-free water to replace the test sample) was also included in the extraction step to verify the absence of environmental contamination, as conducted by Sagheddu. (2017). Each DNA sample was divided into two portions for MiSeq PE300 sequencing and real-time PCR. Purified DNA was stored at −20℃ until used.
2.5 Real-Time Quantitative PCR Assay
The abundance ofandgenes in the DNA extracts was determined by using a real-time quantitative PCR (qPCR) assay. The qPCR amplification was carried out by 9600 plus real-time PCR system (Applied Biosystems, USA). Fragments of bacterialgenes (468bp, V3-V4 region) andgenes (508bp) were amplified by the primer pairs 338F-806R and A189F- mb661R, respectively. These primers were demonstrated to work effectively in previous studies (Shrestha., 2008; Armingohar., 2014). The reaction mixture (20 μL) consisted of 10μL of SYBR®Premix Ex Tag TM (Ta- KaRa, Dalian, China), 0.2μmolL−1of each primer, and 2μL of the template DNA. Data are means±SD of three te- chnical replicates.
2.6 High-Throughput Sequencing and Data Processing
PCR amplification and Illumina MiSeq sequencing of the DNA extracts were conducted in Majorbio Bio-Pharm Technology Co., Ltd., Shanghai, China. The bacterial and methanotrophic community were analyzed by using the primer pairs 338F-806R and A189F-mb661R targeting theandgenes for PCR amplification. Thecommunity structure, richness, and diversity of bacteria (and methanotrophs in particular) were studied through Illumina high-throughput sequencing of bacterialandgenes. The amplicons were paired-end sequenced on an Illumina MiSeq PE300 platform. The raw reads have been submitted to the NCBI Sequence Read Archive (SRA) database with the accession number of SRP189409. Paired-end reads (andgenes) with no less than 20 bases were merged by the FLASH program, and truncated by removing low-quality fragments, barcodes and primers. The sequences were clustered against the database at 97% sequence similarity with UCLUST(Sengupta and Dick, 2017). Assembledandgene sequences were taxonomically classified by using the SILVA SSU 128 and NCBI databases, respectively, with a confidence threshold of 70%. The phylogenetic tree was constructed by neighbor-joining method in the Mega 5.0 software (Saitou and Nei, 1987; Tamura., 2007).
3 Results
3.1 Lipid Biomarker
Figs.1 and 2 show the chromatograms of three total lipid fractions (one initial sample and two incubated samples), with compounds listed in Tables 1 and 2. Both-alkanes and fatty acids were determined in this study.
-alkanes in all studied sediments range in carbon number from 11 to 40, with C15–C20-alkanes as the most dominant homologues (accounting for 65.6% to 86.6% of total-alkanes) (Table 1 and Fig.1). Compared to the ori- ginal sample, concentrations of most-alkanes in sediment samples from two incubations were elevated noticeably (Fig.1). Average chain lengths (ACL) for three samples were similar (30), and all values were within the range calculated from land-derived vegetation (Guillemot., 2019). A strong odd-over-even carbon number predominance, indicated by carbon preference index (CPI), was observed in the original sediment sample (CPI=4.4). In contrast, a much lower CPI value was observed in the sample from incubation i (1.7). The CPI value in the sample from incubation ii, however, is slightly high (2.6) but still much lower than that of the original sediment sample. Low CPI values of two incubated samples reflect degraded organic matter (Grewer., 2018) and similar values also have been reported by Bröder. (2016) and Fang. (2018).
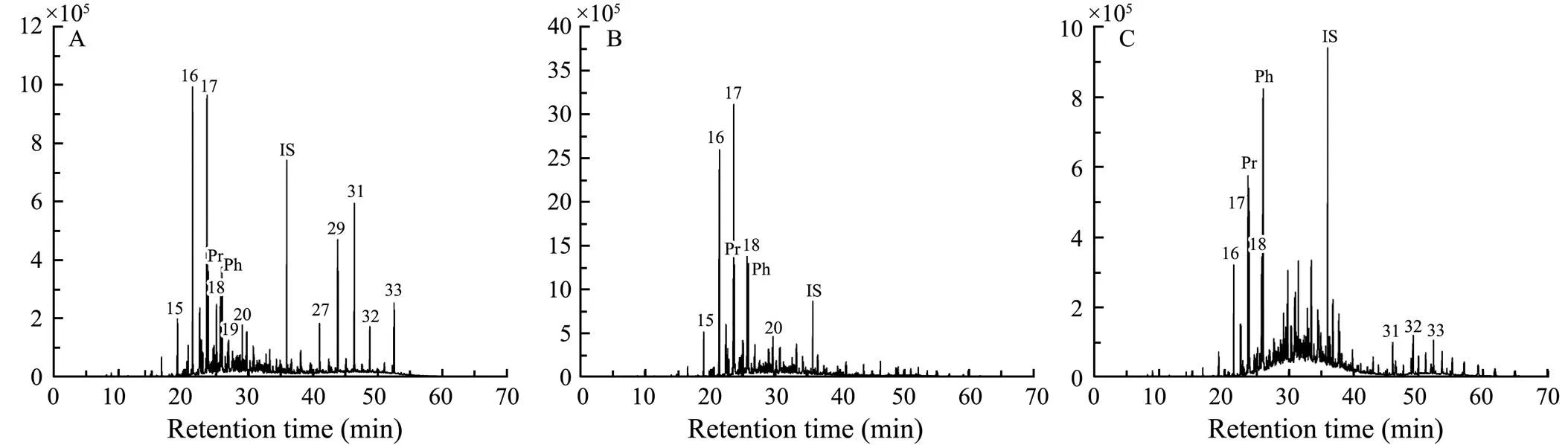
Fig.1 Distribution patterns of n-alkanes in studied sediment samples (m/z=57; IS, internal standard). A, original sample; B, incubation i sample; C, incubation ii sample.

Fig.2 Distribution patterns of fatty acids in studied sediment samples (m/z=74; IS, internal standard). A, original sample; B, incubation i sample; C, incubation ii sample.

Table 1 Concentration of n-alkanes (%) for sediment samples used in this study
Notes: ACL=(25C25+27C27+29C29+31C31+33C33+35C35)/(C25+C27+C29+C31+C33+C35); CPI=(∑oddC21–33+∑oddC23–35)/(2×∑oddC22–34).
Fatty acids in the sediments range in carbon number from C8to C30. In the initial sediment sample, saturated fatty acids dominated in the record, with C14:0, C16:0and C18:0being the most abundant homologues (Table 2 and Fig.2). Compared to the original sample, the incubated samples displayed much higher proportions of C14:0and lower pro- portions of C18:0. Besides, noticeably low ratios of long-chain-length to short-chain-length lipids (TARFA) were ob- served in two incubated samples, which were approxima- tely 6 to 9 times lower than that in the original sediment sample. Of note, there was no discernible trend in the concentration of-alkanes between two incubated samples.
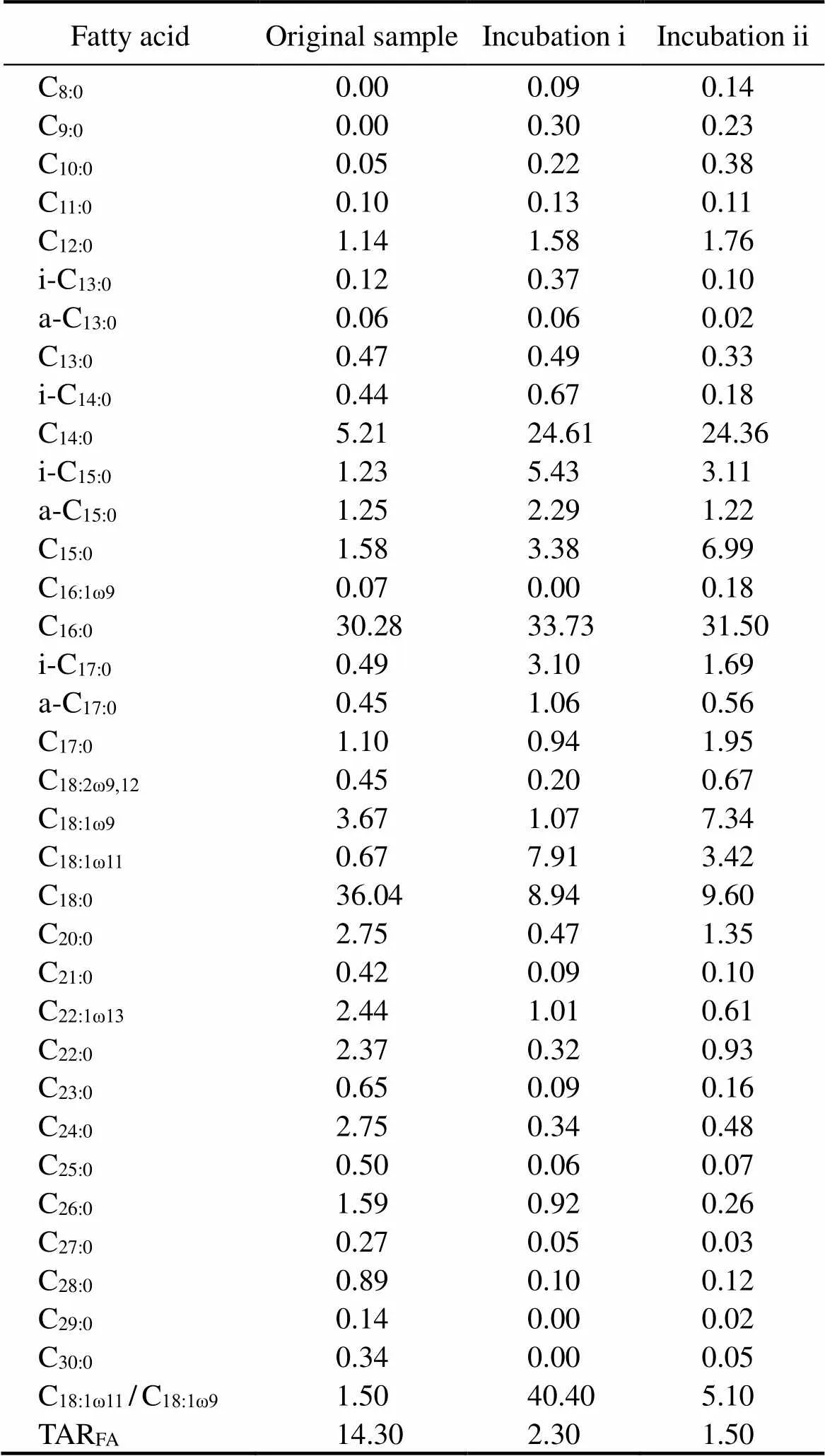
Table 2 Concentration of free fatty acid (%) for sediment samples used in this study
Note: TARFA=(C24+C26+C28)/(C12+C14+C16).
3.2 Bacterial 16S rRNA Gene
The high-throughput sequencing analysis was performed by targeting the bacterialgene of the studied samples. The calculated Alpha-diversity indices are presented in Table 3. Good’s coverage values (at the 97% similarity level) were greater than 0.99 for all samples, indicating that the majority of bacterial community was captured at this sequencing depth. A total 433 OTUs were obtained in the original sediment sample, whereas the number of OTUs in the enrichment samples after incubations decreased significantly. Correspondingly, compared to the original sample, the lower Shannon and Chao 1 in- dices were observed in experimental samples, indicating lower richness and diversity. In contrast with incubation i, total OTUs of incubation ii decreased (157. 104) whereasShannon and Chao 1 indices increased (2.1. 2.4 and 197. 119, respectively), demonstrating that hydrocarbon com-positions influenced the microbial community activity in two incubations.
In this study 171 different bacterial families were detect- ed for all samples, and the dominant families are shown in in Fig.3a. For the original sample Planococcaceae was the most abundant family accounting for 48.11% of all sequences, followed by Moraxellaceae (18.55%), Methylocystaceae (3.60%, type II methanotroph) and Carnobacteriaceae (3.15%). After aerobic oxidation incubations, the bacterial composition changed drastically. Methylococcaceae, which belongs to type I methanotrophs and only accounted for 0.05% in the original sample, became the most abundant family in the experimental samples (40.81% for incubation i and 41.38% for incubation ii). The predominant family in the original sample, Planococcaceae, only accounted for 7.58% and <1.00% in sediment samples from incubations i and ii, respectively. Moraxellaceae,which was also abundant in the original sample, decreased significantly after incubations. A similar variation pattern was also seen for Methylocystaceae, which accounted for <0.1% in incubated samples. Discrepancies between two incubations are also noticeable. Phyllobacteriaceae, Rhi- zobiaceae, and Rhodocyclaceae were more abundant in the sample from incubation ii (each accounting for 10%), whereas Flavobacteriaceae was second only to Methylococcaceae in the sample from incubation i (Fig.3a).

Table 3 Results of bacterial community diversity and richness based on the 16S rRNA and pmoA genes in sediments

Fig.3 Taxonomic composition of bacterial (a) and methanotrophic (b) communities in studied sediment samples.
3.3 Functional Gene
Illumina sequencing ofgene was specially carried out to examine the methanotroph diversity in sediment samples. Chao 1 species richness and Shannon’s diversity indexes were listed in Table 3. Good’s coverage values ofgene (at the 97% similarity level) were 1, suggesting that the obtained sequences could adequately reveal the diversity of methane-degrading bacteria in samples. 14020 high-quality sequences were obtained from the ori- ginal sediment sample, which were classified into 55 OTUs by using a similarity cutoff of 97%. Compared to the ori- ginal sample, the sediment samples from two incubations were similar and displayed much lower OTUs (3 for incubation i and 4 for incubation ii), as well as lower Shan- non (0.3) and Chao 1 indices (3.5).
The community structure of thegenes in samples was analyzed based on the dominant OTUs and by constructing phylogenetic trees (Figs.3b and 4). Phylogenetic analysis revealed the presence of both type I methanotro- phs (species,andspfrom the family Methylococcaceae) and type II methanotrophs (speciesspfrom the family Methylocystaceae) in studied sediment samples. However, only a few ofsequences (1.94%) affiliated tosp. (OTU29) were observed in the original sample, while the remainder were either non-methanotrophs (., Rhodospirillaceae bacteriums) or could not be assigned to any species that are deposited in public domain database. In contrast, all dominantsequences from two incubations were identical and affiliated to type I methanotrophs. Nevertheless, in the sample from incubation i, the number ofsequences affiliated to(OTU56) was dominant (91.99%) among type I methanotrophs, whereas those affiliated tosp. (OTU57) and(OTU58) only accounted for 4.88% and 3.14% respectively. In the sample from incubation ii, however,was predominant, followed byi, whereassp. was much less active, with their proportions being 91.53%, 7.64% and 0.01%, respectively.

Fig.4 Neighbor-joining tree of 7 most abundant OTUpmoA, with selected reference sequences. The relative abundances (%) for different samples are given to the right of each OTUpmoA. Bootstrap values >50% (500 repetitions) are shown on nodes. The scale bar indicates sequence dissimilarity between nodes.
3.4 Abundance of Total Bacterial 16S rRNA and pmoA Genes
The abundances of bacterialandgenes were determinedqPCR (Fig.5). Compared to the ori- ginal sediment sample, both of two incubations are characterized by much higher abundances of the bacterialgene, with the average value being (2.2±0.2)×1010copiespergramsoil. Although the abundances ofgene also increased after aerobic oxidation incubation, dif- ferent variation patterns were displayed between two incubations. The abundance ofgenes from incubation i is 1.4×109copiespergramsoil, which is much higher than that from incubation ii (2.3×107copiespergramsoil).
4 Discussion
4.1 Sedimentary Sources and Microbial Activity Indicated by Lipid Analysis
High concentrations of long chain-alkanes and fatty acids, as well as high ACL values were generally regard- ed as indicators of land-derived input (Meyers, 2003; Guillemot., 2019). The CPI value is also an effective indicator of organic matter sources, with high values normally revealing terrestrial vegetation sources and low va- lues (around 1) generally related to the significant bacterial activity or petroleum pollution (Zhu., 2005; Duncan., 2019; and references therein). Parkes and Taylor (1983) also suggested that the fatty acid analysis could be used to distinguish different types of microbes within sediments, which has been demonstrated by subsequently published studies (Sundh., 1995; Dedysh., 2007).
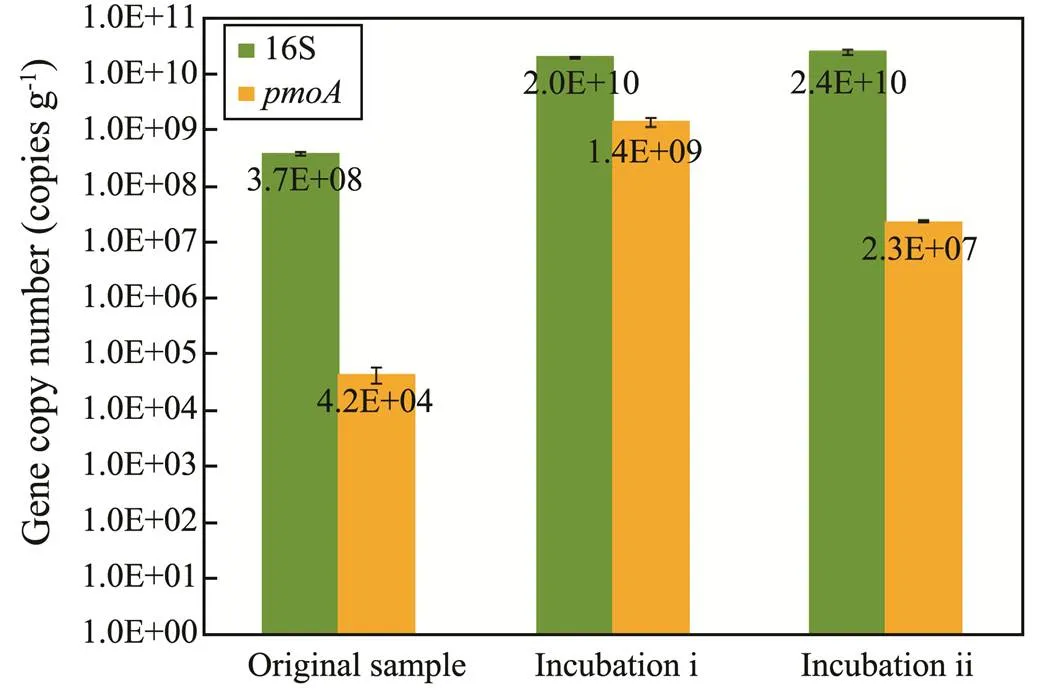
Fig.5 Bacterial 16S rRNA and pmoA genes copy numbers quantified by using qPCR for studied sediment samples.
In this study, the high ACL value (30.4) and the strong odd-over-even carbon number predominance of-alkanes (CPI=4.4), as well as the dominance of long-chain fatty acids, were observed in the original sediment sample, col- lectively indicating the prevailingly terrigenous origin of organic matters in the sediments from Bohai Bay. These features are similar to those of-alkanes and fatty acids from leaf waxes of higher plants (., Bush and McInerney, 2013), in eolian dust samples (., Schreuder., 2018) and in marine sediments (Sinninghe Damsté., 2001), all of which supported a terrigenous origin.
There was no significant variation between two samples treated with pure methane and mixed gas. Nevertheless, in contrast with the original sample, the-alkanes and fatty acid data of the incubated samples indicate the mixture sources of organic matters and noticeable bacterial reworking. The CPI values of-alkanes decreased in two samples that experienced aerobic oxidation, apparently related to the significant bacterial activity (Huang., 1999; Zhu., 2005). Similar values have also been re- ported by Bröder. (2016) and Fang. (2018). Higher concentrations of short chain acids (., lower TARFAvalues, Table 2) in those two samples were also related to the presence of bacterial communities that uti- lize these fatty acids as their carbon sources. This utilization has been reported in studies from estuaries (Zhang., 2015), lakes (Arnold., 2018) and marine sediments (Parkes and Taylor, 1983). Although in this study high concentrations of cyclopropyl fatty acids (Ñ17 andÑ19), both of which were expected in the aerobic bacteria according to Parkes and Taylor (1983), were not detected in incubated samples, they are characterized by higher C18:1ω11/C18:1ω9values (40.4 and 5.1 for incubations i and ii respectively) than the original sample (1.5). According to Volkman. (1980), these high C18:1ω11and low C18:1ω9concentrations (and thus high C18:1ω11/C18:1ω9ratios) are typical of the aerobic bacterial activity.
In conclusion, although the limited sample number of this study prevents any further implications from lipid analysis, the noticeable contrasts of-alkanes and fatty acids between the original sample and two incubated samples demonstrated that microbial aerobic oxidation progres- sed in the two incubations. This is further demonstrated by results from the DNA analyses.
4.2 Hydrocarbon-Oxidizing Bacterial Activity in Response to Aerobic Oxidation
Significant variations in the bacterial communities wereseen between uninoculated and incubated samples, as well as between two incubated samples. Compared to incuba- ted samples, the uninoculated sample was characterized by a high proportion of type II methanotrophs (family Methylocystaceae) and a low proportion of type I methanotrophs (family Methylococcaceae). Generally, the com- petition between types I and II methanotrophs depends on the concentrations of CH4and O2and also on the pre- sence of nitrogen (Bull., 2000; Siljanen., 2012; Sengupta and Dick, 2017). Previous studies suggested that type I methanotrophs seem to prefer environments with plentiful O2and limited CH4, whereas type II methanotrophs dominate in environments with high concentrations of CH4and limited O2(Graham., 1993; Amaral and Knowles, 1995; Shrestha., 2008). The results from this study thus agree with this finding since the se- diment samples here were collected from the seafloor of the gas- and oil-rich Bohai Bay, which corresponds to an environment with plentiful CH4and limited O2, whereas the incubations were conducted with adequate O2and limited hydrocarbons.
Monitoring the gas composition during the incubation ii showed that both ethane and propane were completely consumed after the incubation, which indicated the occurrence of ethane- and propane-oxidizing bacteria in the study samples. However, currently less is known about the organisms that oxidize ethane or propane in marine environments. The known such organisms, which are pri- marily high G+C Gram-positive bacteria (,,,, and) orspecies (Redmond., 2010), were not observed in our samples. Nevertheless, the comparison between two incubations shows that the amounts of families Phyllobacteriaceae, Rhizobiaceae, and Rhodocycla- ceae increased significantly in incubation ii, seemingly proposing that these organisms might consume ethane and/or propane. To test whether this is correct, pure cultures of these organisms should be conducted in the following work. It should be noted that Redmond. (2010) suggested that some species from the family Methylococcacea, which had previously been thought to be obligate methanotrophs, might be capable of growing on ethane or propane. Our study seems to support this since the proportion of Methylococcacea noticeable increase in samples from incubations i to ii.
Interestingly, although nearly all methanotrophs found in two incubations were type I, these species are quite different. Thewas active when pure methane acts as carbon source, whereastook the predominant proportion when mixed gases are available. The observed discrepancies could result from the higher methane concentrations in incubations i than ii, but more likely indicated that some me- thanotrophs could utilize ethane and propane, as suggested by Berthe-Corti and Fetzner (2002), Kinnaman. (2007) and Redmond. (2010). Whatever the case, these results indicate that the bacterial community in marine sediments may give the information regarding marine hydrocarbon seepages, especially for the leakage of trace amounts of hydrocarbons (Okita., 2020). Characte- ristic methanotrophic communities could be used not only to determine whether hydrocarbon leakage occurred, but also to know about the hydrocarbon composition.
4.3 Application to Natural Systems
The distributions of microbial communities in marine sediments and overlying water have previously been applied to approximate the occurrence and the extent of hy- drocarbon (especially methane) oxidation. This approach is well established for methanotrophs under pure methane condition, but less so under mixed gas condition. This study identifies several methanotroph lineages as participants in the oxidation of gaseous hydrocarbons in marine seeps. The fact that the methanotroph community structure is quite different under pure methane and mixed gas conditions is promising. Once justified by following studies, this can be used to determine the gas compositions in hydrocarbon seeps by sequencinggenes, especially in places where the direct analyses of gas composition are hard. The characteristics of methanotrophs presented in this work are specific to oxic marine sediments at 28℃ to facilitate the rapid growth of bacteria, which is slightly higher than thetemperature in the sampling place. Besides, in a given environment, the expression of methanotrophs will depend on a variety of environmental factors including pH, salinity, nutrient level,. Thus, care must be taken in applying the results obtained in this study to the natural environmental systems.
5 Conclusions
In this study, we have combined both lipid and DNA analyses to elucidate the role of methanotrophs in the marine sediments collected from Bohai Bay, with different hydrocarbons as carbon source over the incubation period. Although both incubations demonstrated that type I me- thanotrophic populations might play a particularly important role in the marine ecosystem, they exhibited the pronounced variations with respect to their activities and population sizes.was predo- minant when pure methane acts as carbon source, whereasare much more active when ethane and propane were also involved. The results collectively suggest that sequencing ofandgenes may be used to identify the occurrence and/or extent of aerobic oxidation, as well as to determine the gas compositions in hydrocarbon seeps.
Acknowledgements
This work was supported by the Natural Science Foundationof Shandong Province (No. ZR2020QD070), the National Natural Science Foundation of China (No. 4187 6051), and the China Geological Survey Project (No. DD 20190221).
Amaral, J. A., and Knowles, R., 1995. Growth of methanotrophs in methane and oxygen counter gradients., 126: 215-220.
Armingohar, Z., Jørgensen, J. J., Kristoffersen, A. K., Abesha- Belay, E., and Olsen, I., 2014. Bacteria and bacterial DNA in atherosclerotic plaque and aneurysmal wall biopsies from patients with and without periodontitis., 6: 23408.
Arnold, T. E., Kenney, W. F., Curtis, J. H., Bianchi, T. S., and Brenner, M., 2018. Sediment biomarkers elucidate the Holo- cene ontogeny of a shallow lake., 13: e0191073.
Berthe-Corti, L., and Fetzner, S., 2002. Bacterial metabolism of-alkanes and ammonia under oxic, suboxic and anoxic conditions., 22: 299-336.
Boetius, A., and Wenzhöfer, F., 2013. Seafloor oxygen consum- ption fuelled by methane from cold seeps., 6: 725-734.
Bröder, L., Tesi, T., Andersson, A., Eglinton, T. I., Semiletov, I. P., Dudarev, O. V., Roos, P., and Gustafasson, Ö., 2016. Historical records of organic matter supply and degradation sta- tus in the East Siberian Sea., 91: 16-30.
Bull, I. D., Parekh, N. R., Hall, G. H., Ineson, P., and Evershed, R. P., 2000. Detection and classification of atmospheric methane oxidizing bacteria in soil., 405: 175-178.
Bush, R. T., and McInerney, F. A., 2013. Leaf wax-alkane dis- tributions in and across modern plants: Implications for paleoecology and chemotaxonomy., 117: 161-179.
Cui, H. P., Su, X., Wei, S. P., Zhu, Y. H., Lu, Z. Q., Wang, Y. F., Li, Y., Liu, H., Zhang, S., and Pang, S., 2018. Comparative analyses of methanogenic and methanotrophic communities between two different water regimes in controlled wetlands onthe Qinghai-Tibetan Plateau, China., 75: 484-491.
Dedysh, S. N., Belova, S. E., Bodelier, P. L., Smirnova, K. V., Khmelenina, V. N., Chidthaisong, A., Trotsenko, Y. A., Liesack, W., and Dunfield, P. F., 2007.sp. nov., a novel type II methanotrophic bacterium possessing ‘sig- nature’ fatty acids of type I methanotrophs., 57: 472-479.
Ding, H., and Valentine, D. L., 2008. Methanotrophic bacteria occupy benthic microbial mats in shallow marine hydrocarbon seeps, Coal Oil Point, California., 113: G01015.
Duncan, B., McKay, R., Bendle, J., Naish, T., Inglis, G. N., Moos- sen, H., Levy, R., Ventura, G., Lewis, A., and Chamberlain, B., 2019. Lipid biomarker distributions in Oligocene and Miocene sediments from the Ross Sea region, Antarctica: Implications for use of biomarker proxies in glacially-influenced settings.,516: 71-89.
Etiope, G., 2015.. Springer, Switzerland, 118-130.
Fang, J., Wu, F., Xiong, Y., Li, F., Yang, H., Wang, S., and Xie, Y., 2018. Sources of organic matter in the surface sediments from Lake Sihailongwan Maar and Lake Zhanjiang Maar (Lake Huguangyan Maar) in China., 69: 18-23.
Graham, D. W., Chaudhary, J. A., Hanson, R. S., and Arnold, R. G., 1993. Factors affecting competition between type-I and ty- pe-II methanotrophs in 2-organism, continuous-flow reactors., 25: 1-17.
Grewer, D. M., Lafrenière, M. J., Lamoureux, S. F., and Simpson, M. J., 2018. Spatial and temporal shifts in fluvial sedimentary organic matter composition from a high Arctic watershed impacted by localized slope disturbances., 123: 113-125.
Guillemot, T., Stockhecke, M., Bechtel, A., Ladd, S. N., Nelson, D. B., and Schubert, C. J., 2019. Paleoenvironmental and paleoclimatic variations around Lake Van (eastern Turkey) recorded by sedimentary source specific biomarkers 250–130ka (MIS7 and MIS6)., 225: 105997.
Hanson, R. S., and Hanson, T. E., 1996. Methanotrophic bacteria., 60: 439-471.
Hazeu, W., and de Bruyn, J. C., 1980. Ethane oxidation by metha- ne-oxidizing bacteria., 46: 443-455.
Hinrichs, K. U., and Boetius, A., 2002. The anaerobic oxidation of methane: New insights in microbial ecology and biogeoche- mistry. In:. Wefer., eds., Springer, Berlin, 457-477.
Holmes, A. J., Owens, N. J. P., and Murrell, J. C., 1995. Detection of novel marine methanotrophs using phylogenetic and functional gene probes after methane enrichment., 141: 1947-1955.
Huang, Y., Street-Perrott, F. A., Perrott, R. A., Metzger, P., and Eglinton, G., 1999. Glacial-interglacial environmental changes inferred from molecular and compound-specific δ13C analyses of sediments from Sacred Lake, Mt. Kenya., 63: 1383-1404.
Hu, J. F., Zhang, H. B., and Peng, P. A., 2006. Fatty acid composition of surface sediments in the subtropical Pearl River Estuary and adjacent shelf, southern China., 66: 346-356.
Inagaki, F., Tsunogai, U., Suzuki, M., Kosaka, A., Machiyama, H., Takai, K., Nunoura, T., Nealson, K. H., and Horikoshi, K., 2004. Characterization of C1-metabolizing prokaryotic communities in methane seep habitats at the Kuroshima Knoll, southern Ryukyu Arc, by analyzing,,,,andgenes., 70: 7445-7455.
Kinnaman, F. S., Valentine, D. L., and Tyler, S. C., 2007. Carbon and hydrogen isotope fractionation associated with the aerobic microbial oxidation of methane, ethane, propane and butane., 71: 271-283.
Lashof, D. A., and Ahuja, D. R., 1990. Relative contributions of greenhouse gas emissions to global warming., 344: 529- 531.
Li, J., Liu, C. L., He, X. L., Santosh, M., Hu, G. W., Sun, Z. L., Li, Y., Meng, Q., and Ning, F., 2019. Aerobic microbial oxidation of hydrocarbon gases: Implications for oil and gas exploration., 103: 76-86.
Liu, C. L., Meng, Q. G., He, X. L., Li, C. F., Ye, Y. G., Zhang, G. X., and Liang, J. Q., 2015. Characterization of natural gas hydrate recovered from Pearl River Mouth Basin in South China Sea., 61: 14-21.
Meyers, P. A., 2003. Applications of organic geochemistry to pa-leolimnological reconstructions: A summary of examples from the Laurentian Great Lakes., 34: 261- 289.
Milkov, A. V., 2005. Molecular and stable isotope compositions of natural gas hydrates: A revised global dataset and basic interpretations in the context of geological settings., 36: 681-702.
Okita, N., Hoaki, T., Suzuki, S., and Hatamoto, M., 2020. Characteristics of aerobic methane-oxidising bacterial community at the sea-floor surface of the Nankai Trough., 71:1252-1258.
Op den Camp, H. J., Islam, T., Stott, M. B., Harhangi, H. R., Hynes, A., Schouten, S., Jetten, M., Birkeland, N., Pol, A., and Dunfield, P., 2009. Environmental, genomic and taxono- mic perspectives on methanotrophic Verrucomicrobia., 1: 293-306.
Parkes, R. J., and Taylor, J., 1983. The relationship between fatty acid distributions and bacterial respiratory types in contemporary marine sediments., 16: 173-189.
Redmond, M. C., Valentine, D. L., and Sessions, A. L., 2010. Iden- tification of novel methane-, ethane-, and propane-oxidizing bacteria at marine hydrocarbon seeps by stable isotope probing., 76: 6412-6422.
Reeburgh, W. S., 2007. Oceanic methane biogeochemistry., 107: 486-513.
Sagheddu, V., Patrone, V., Miragoli, F., and Morelli, L., 2017. Abundance and diversity of hydrogenotrophic microorga- nisms in the infant gut before the weaning period assessed by denaturing gradient gel electrophoresis and quantitative PCR., 4: 29.
Saitou, N., and Nei, M., 1987. The neighbor-joining method: A new method for reconstructing phylogenetic trees., 4: 406-425.
Schreuder, L. T., Stuut, J. B. W., Korte, L. F., Damsté, J. S. S., and Schouten, S., 2018. Aeolian transport and deposition of plant wax-alkanes across the tropical North Atlantic Ocean., 115: 113-123.
Sengupta, A., and Dick, W. A., 2017. Methanotrophic bacterial diversity in two diverse soils under varying land-use practices as determined by high-throughput sequencing of thegene., 119: 35-45.
Shrestha, M., Abraham, W. R., Shrestha, P. M., Noll, M., and Conrad, R., 2008. Activity and composition of methanotrophic bacterial communities in planted rice soil studied by flux measurements, analyses ofgene and stable isotope probing of phospholipid fatty acids., 10: 400-412.
Siljanen, H. M., Saari, A., Bodrossy, L., and Martikainen, P. J., 2012. Seasonal variation in the function and diversity of me- thanotrophs in the littoral wetland of a boreal eutrophic lake., 80: 548-555.
Sinninghe Damsté, J. S., Schefuss, E., Versteegh, G. J. M., and Jansen, J. H. F., 2001. Marine and terrigenous lipids in sou- theast Atlantic sediments (Leg 175) as paleoenvironmental in- dicators: Initial results. In:. Ocean Drilling Program, Texas A & M University, 1-34.
Sundh, I., Borgå, P., Nilsson, M., and Svensson, B. H., 1995. Estimation of cell numbers of methanotrophic bacteria in boreal peatlands based on analysis of specific phospholipid fatty acids., 18: 103-112.
Tamura, K., Dudley, J., Nei, M., and Kumar, S., 2007. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0., 24: 1596-1599.
Valentine, D. L., 2011. Emerging topics in marine methane biogeochemistry., 3: 147-171.
Valentine, D. L., Blanton, D. C., Reeburgh, W. S., and Kastner, M., 2001. Water column methane oxidation adjacent to an area of active hydrate dissociation, Eel River Basin., 65: 2633-2640.
Volkman, J. K., Eglinton, G., Corner, E. D., and Forsberg, T. E. V., 1980. Long-chain alkenes and alkenones in the marine co- ccolithophorid., 19: 2619- 2622.
Wang, F. P., Zhang, Y., Chen, Y., He, Y., Qi, J., Hinrichs, K. U., Zhang, X., Xiao, X., and Boon, N., 2014. Methanotrophic archaea possessing diverging methane-oxidizing and electron- transporting pathways., 8: 1069-1078.
Zhang, N. L., 2016. Geochemical of hydrocarbon gas from the surface seabed sediment in Bohai Bay Basin. Master thesis. Jilin University, 1-70.
Zhang, X., Bianchi, T. S., and Allison, M. A., 2015. Sources of matter in sediments of the Colville River Delta, a multi-proxy approach., 87: 96-106.
Zhu, C., Pan, J. M., Lu, B., Hu, C. Y., Liu, X. Y., Ye, X. R., and Xue, B., 2005. Source indication and accumulative effect of sedimentary organic matter in the Changjiang Estuary, the old Huanghe River subaqueous delta and the East China Sea shelf., 23: 36-46 (in Chinese with English abstract).
July 23, 2020;
November 4, 2020;
December 10, 2020
© Ocean University of China, Science Press and Springer-Verlag GmbH Germany 2021
. Tel: 0086-532-85755855 E-mail: qdliuchangling@163.com
(Edited by Chen Wenwen)
杂志排行

Journal of Ocean University of China的其它文章
- Case Study of a Short-Term Wave Energy Forecasting Scheme:North Indian Ocean
- Temporal and Spatial Characteristics of Wave Energy Resources in Sri Lankan Waters over the Past 30 Years
- Vibration Deformation Monitoring of Offshore Wind Turbines Based on GBIR
- Dependence of Estimating Whitecap Coverage on Currents and Swells
- 3-Aminopropyltriethoxysilane Complexation with Iron Ion Modified Anode in Marine Sediment Microbial Fuel Cells with Enhanced Electrochemical Performance
- Removal of Sulfamethazine by Corn Biochars from Aqueous Solution: Sorption Mechanisms and Efficiency