两种诱导子对番茄抗病及根际细菌群落的影响
2021-06-23苏莹莹金朝霞崔宇兴
苏莹莹, 金朝霞, 崔宇兴
(大连工业大学 生物工程学院,大连 116034)
番茄在种植生产中常受多种病虫害影响,由灰葡萄孢菌(Botrytiscinerea)引起的灰霉病易造成番茄生长缓慢,果实减产[1]。目前,针对灰霉病的防治主要为化学防治,但化学农药长期施用易污染环境,造成农作物累积毒素,危害人体健康。生物防治通过来源于植物根际的促生菌、拮抗菌及其相关菌制剂对防治番茄灰霉病起到良好的拮抗及防治效果[2-3]。在植物与病原菌互作过程中,生物及化学因子可作为诱抗剂激发植物天然的防御机制,引发植物的诱导抗病性[4]。β-氨基丁酸(BABA)是一种非蛋白氨基酸,可从番茄根系物中分离得到,研究表明BABA在番茄[5]、烟草[6]以及马铃薯[7]等作物中可通过激发植物系统抗性来抵抗病害,是一种具有潜能的高效植物诱抗剂。假单胞菌属是根际益生菌的重要组成部分,作为重要的生防菌,利用竞争作用、产生抗生素等抑菌物质以及诱导植物产生抗病性等机制发挥抗病效果[8-9]。
根际微生物组成也是反映作物健康情况的重要指标,与作物病虫害的发生关系密切。通过改善土壤营养组成、与有机体互作调控植物生长、响应生物及非生物胁迫等方式参与植物根际微生态调节[10-11]。但在防治番茄灰霉病的机制研究中,不同诱导因子的防病机制与番茄根际细菌群落的关系尚不明确。研究选取课题组保存的具有广谱抗性的绿针假单胞菌(Pseudomonascholoeaphtis)PA6以及具有诱抗效果的β-氨基丁酸(BABA),通过盆栽试验及实时荧光定量PCR技术研究不同诱导下的番茄灰霉病发生及抗性基因的表达情况,并用高通量测序研究根际细菌的群落响应特征,从植物系统抗性及根际细菌群落结构变化角度完善番茄抗灰霉病的研究。
1 材料与方法
1.1 材料
供试番茄品种为“中蔬四号”,购于青县王镇店种子繁育站,生防菌绿针假单胞菌(Pseudomonascholoeaphtis)PA6以及病原菌灰葡萄孢菌(Botrytiscinerea)均由本课题组保藏。化学试剂β-氨基丁酸(BABA)购自北京索莱宝科技有限公司;育苗土壤按营养土∶蛭石∶椰糠(3∶1∶1)经高温灭菌后混合而成,其中营养土由江苏淮安苏淮提供,蛭石及椰糠由淮安淮农农业科技开发有限公司提供。
1.2 方法
1.2.1 番茄培养条件
番茄种子经过75%乙醇、NaClO消毒,经过2 d浸种暗处理催芽,待种子在无菌平皿中长至两叶一心时,移至装有通过高温灭菌土壤的56 孔育苗盘中。继续培养番茄植株至四叶一心期,用于后续诱导试验。
1.2.2 番茄植株诱导预处理
配制绿针假单胞菌PA6菌悬液调整浓度为(1×108mL-1),并配制0.5 mmol/L的BABA溶液备用。设3组处理,CK:无菌蒸馏水处理;PA:绿针假单胞菌菌悬液处理;BABA:0.5 mmol/L的BABA溶液处理。利用灌根法,每组60 mL处理液分3次进行灌根处理,沿番茄幼苗根部缓慢灌入根际土壤中,每组处理15株,共计45株。上述处理后的番茄植株继续培养2 d后待接种。
1.2.3 灰葡萄孢菌接种及病情调查
配制灰葡萄孢菌孢子悬液(1×106mL-1)采用叶片喷施法接种病原菌,侵染后植株经16 h/8 h光暗交替培养3 d,5 d后观察发病情况计算发病率,病害分级标准参照闫艳华等[12]方法计算病情指数,并于第5天收集根际土壤及番茄叶片进行后续试验。
1.2.4 番茄NPR1和PR1基因表达分析
利用TRIzol法提取番茄叶片总RNA并反转录,以反转录合成的cDNA为模板,参照表1相应基因引物,使用实时荧光定量PCR仪(LightCycle®480II)进行PCR扩增。按照20 μL体系,每个反应包括10 μL的SYBR GREEN I,上下游引物各0.2 μL,cDNA 2 μL并补加7.6 μL ddH2O,引物如表1所示,设定PCR反应条件:95 ℃ 预变性5 min;95 ℃ 变性15 s,50 ℃ 退火45 s,40个循环;72 ℃ 延伸10 min。得到抗性基因NPR1、PR1以及番茄内参基因actin的Ct值并利用2-ΔΔCT方法进行计算分析基因表达量的变化。
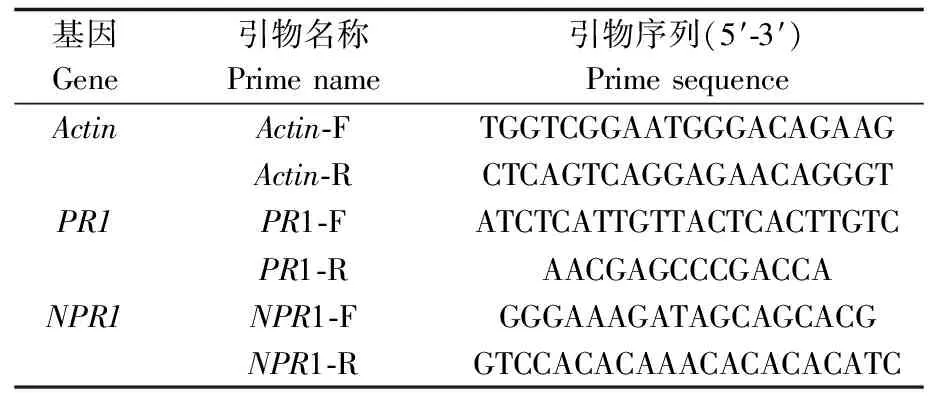
表1 试验所用引物序列
1.2.5 根际土壤收集
采用抖根法从土壤里取出不同处理组的番茄后,剧烈抖动植株根系至仅有1 mm厚度的土壤保留在根系表面,将这部分土壤收集并经液氮冷冻后装入无菌袋中,置于-80 ℃冰箱备用。
1.2.6 土壤微生物DNA提取及16S rDNA高通量测序
采用CTAB方法对各组根际土壤DNA进行提取,并测定DNA纯度及浓度,由北京诺禾致源生物信息科技有限公司(天津)对细菌16S rDNA V4区域利用带Barcode的特异引物进行PCR扩增,将目标条带进行胶回收并纯化后进行文库构建,使用Thermofisher的Ion S5TMXL进行上机测序。
1.2.7 数据分析及处理
高通量测定试验分析利用Cutadapt V1.9.1软件对序列数据进行质量控制,再通过Mothur方法校正去除嵌合体序列获得有效数据,最后采用Uparse v7.0.1001软件对所有样品序列进行OTU(Operational taxonomic units)聚类分析,用Mothur方法与SILVA132(http://www.arb-silva.de/)的SSUrRNA数据库进行物种注释分析,获得不同分类水平的分类信息,统计各样本的群落组成并利用Mothur 1.30.1 软件绘制作图。其他试验数据用Excel处理并用SPSS 23.0进行差异显著性分析。
2 结果与分析
2.1 绿针假单胞菌及BABA诱导处理对番茄植株灰霉病的影响
由于不同的农艺措施会影响相关抑菌剂的抗病效果[13]。为排除绿针假单胞菌对叶片接种病原菌拮抗作用的影响,两种诱导因子均采用灌根的施用方式,通过对比番茄植株叶片发病率及病情指数可知,诱导处理组的番茄发病率下降,相比空白对照组,接种病原菌3 d后绿针假单胞菌处理组病情指数降低了60.4%,BABA处理组降低了36.6%,第5天不同处理组的发病率及病情指数相比第3天均有所上升,但诱导处理组的发病速率减缓(图1)。可见两种诱导因子均能有效改善番茄发病情况。
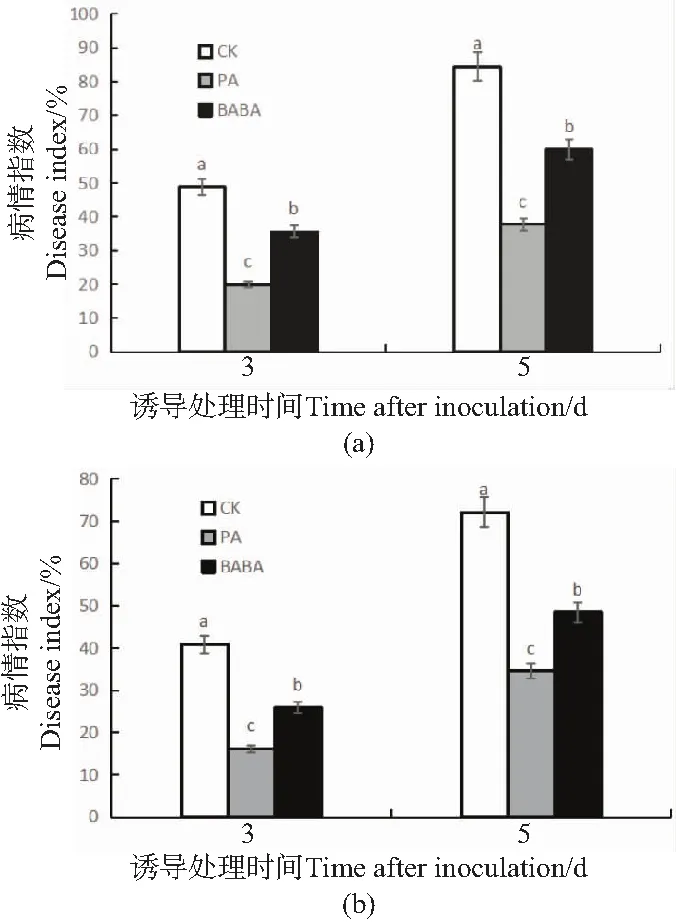
(a)发病率;(b)病情指数。CK:蒸馏水+灰葡萄孢菌;PA:绿针假单孢菌+灰葡萄孢菌;BABA:BABA+灰葡萄孢菌。不同字母表示差异显著(P<0.05)
2.2 抗性基因NPR1及PR1的表达分析
由植物系统抗病性引发的防御反应被多个基因调控,受诱导因子影响会激发不同的信号通路,促进抗性基因的表达。NPR1作为植物诱导抗病性中多种信号转导途径的关键调控节点,位于SA信号通路下游,PR基因上游[14-15]。在植物被诱导产生系统获得抗性(SAR)时,SA信号通路被激活,PRs等病程蛋白基因会做出响应,其中编码PR-1a蛋白的PR1基因是植物抗病性变化的重要标志[16]。如图2所示,经绿针假单胞菌以及BABA诱导后,NPR1基因上调表达,分别为对照组的3.3倍和2.4倍。两组诱导处理中PR1基因的表达量约为对照组的2.9倍。可见相对未经诱导预处理的对照组,两个诱导处理组的番茄叶片中NPR1及PR1基因均显著上调。乔俊卿等[17]发现经枯草芽孢杆菌PTS-394预处理的番茄,防御基因NPR1,PR-1a基因短期高表达,有效减轻了植株灰霉病情况。研究利用不同诱导处理得到类似结果,表明两种诱导处理均可诱导番茄产生系统抗性,激发SA信号通路,进而提高番茄抵抗病原菌的能力。
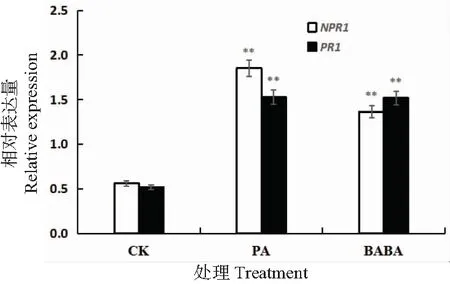
CK:蒸馏水+灰葡萄孢菌;PA:绿针假单孢菌+灰葡萄孢菌;BABA:BABA+灰葡萄孢菌。**表示相关性达到显著水平(P<0.01)
2.3 16S rDNA测序鉴定分析根际细菌群落结构
2.3.1 样品组间主坐标分析
基于加权的主坐标分析结果如图3所示,前两个主成分累计贡献率为42.61%,分别以24.78%、17.83%的贡献率成为解释CK、PA及BABA等3组样品差异的最大特征。BABA及CK处理在PC2轴上相距较远,PA及CK处理在PC1轴上相距较远,说明BABA及PA处理都使根际细菌群落组成发生改变。PA及BABA处理在PC1及PC2轴上都相距了一定距离,表明PA及BABA诱导处理下的细菌群落结构差异明显。3种处理的组间距离较远、组内间距较近,说明组内群落结构相似,组间群落结构差异显著。
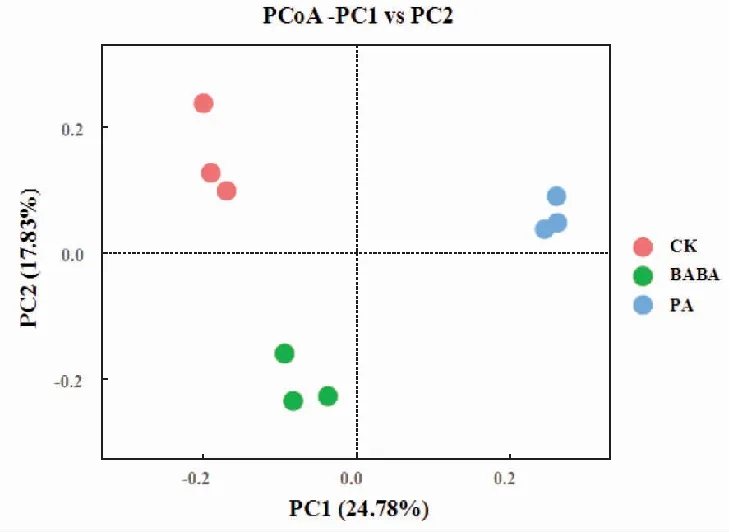
CK:蒸馏水+灰葡萄孢菌;PA:绿针假单孢菌+灰葡萄孢菌;BABA:BABA+灰葡萄孢菌
2.3.2 根际细菌OTU特异性分析
绿针假单胞菌及BABA诱导处理样品间共有、特有的OTU情况(图4)显示,诱导处理组及对照组的番茄根际土壤中的细菌OTU数目存在差异。其中,CK、PA以及BABA 等3组的OTU数目分别为1 592、1 473和1 671,三者OTU数量表现为BABA处理组>CK组>PA处理组,说明BABA处理中的细菌种类最丰富。3组共有的OTU数目为852,且为各组细菌OTU数目的50%以上,说明各处理组优势菌基本组成相似。PA及BABA组共有的细菌OTU数量为1 221,其中PA组中特有252个细菌OTU,而BABA组中特有细菌OTU数量为450,说明不同诱导处理中,BABA预处理根际土壤具有特异性细菌的菌群更多。
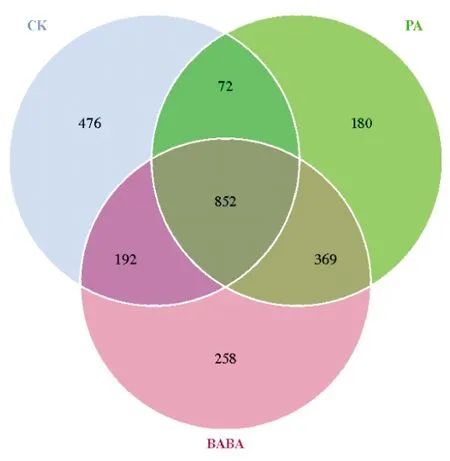
CK:蒸馏水+灰葡萄孢菌;PA:绿针假单孢菌+灰葡萄孢菌;BABA:BABA+灰葡萄孢菌
2.3.3 门水平根际细菌优势菌比较
由图5可知,在门水平上,CK、PA以及BABA等3组样品的优势菌门大致相同,各样品中比例最高的为变形菌门(Proteobacteria),其他高丰度优势菌门主要为拟杆菌门(Bacteroidetes)、放线菌门(Actinobacteria)以及厚壁菌门(Firmicutes)。由图6可知,变形菌门(Proteobacteria)在各组所占比例分别为CK 33.61%、PA 41.39%、BABA 41.81%,经诱导预处理后,高丰度菌群中变形菌门(Proteobacteria)丰度在PA及BABA组中均显著上升,低丰度菌群中浮霉菌门(Planctomycetes)及酸杆菌门(Acidobacteria)丰度下降。门水平上优势菌组成不受诱导处理的影响,但菌群丰度存在差异,这与赖宝春等[18]关于辣椒不同感病情况根际细菌的门水平分析相似。
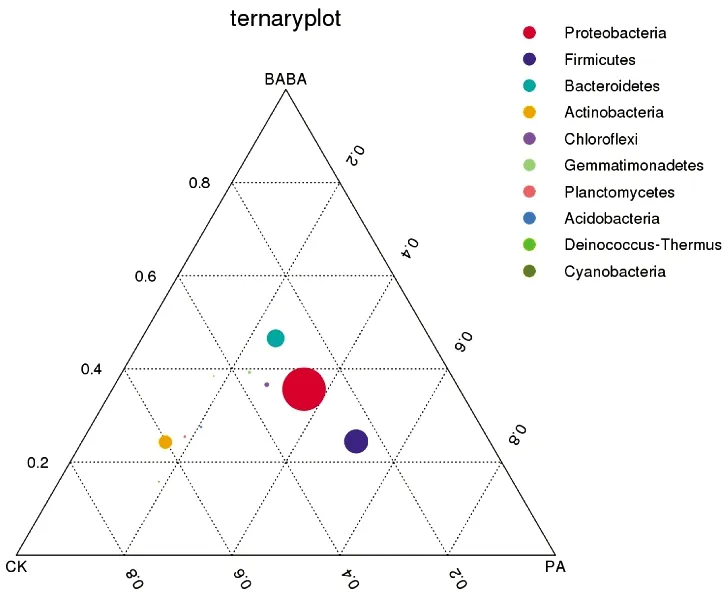
CK:蒸馏水+灰葡萄孢菌;PA:绿针假单孢菌+灰葡萄孢菌;BABA:BABA+灰葡萄孢菌
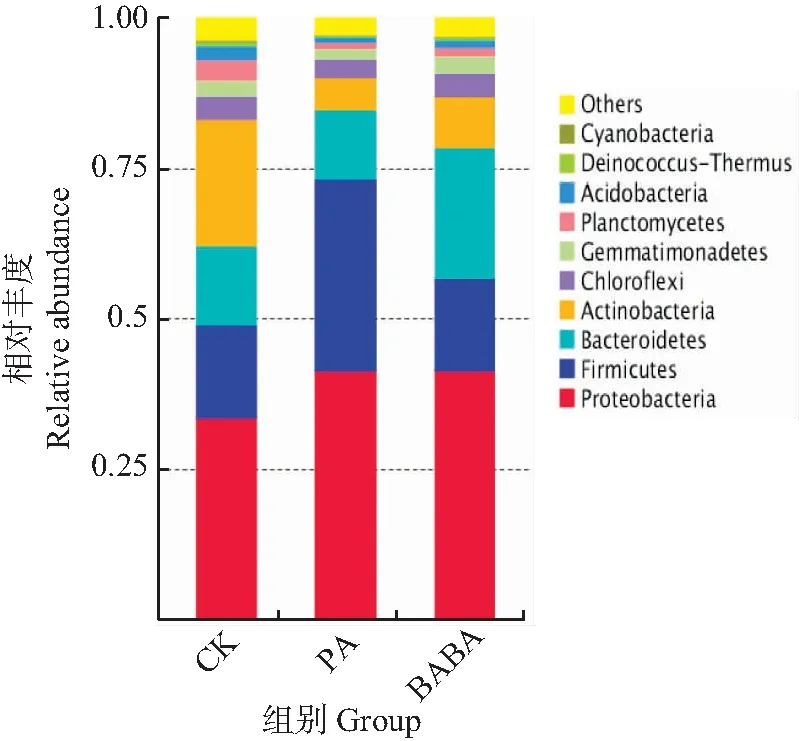
CK:蒸馏水+灰葡萄孢菌;PA:绿针假单孢菌+灰葡萄孢菌;BABA:BABA+灰葡萄孢菌
2.3.4 属水平根际细菌优势菌比较
从8个主要细菌门中筛选出丰度排在前35的菌属。由图7可知,CK组主要富集芽孢杆菌属(Bacillus)、lamia菌属、拟杆菌(Bacteroides)、类卡诺氏菌属(Nocardioides)、微杆菌属(Microbacterium),一些不明细菌菌属也有较高丰度,但这些菌属在PA组及BABA组中的丰度均有所下降。与CK相比,两个诱导处理组都明显富集短波单胞菌属(Brevundimonas)、成对杆菌属(Dyadobacter)以及鞘脂菌属(Sphingobium)。彭方仁等[19]从油茶根际分离得到一株具有潜在固氮作用的鞘脂菌属,可改善土壤磷成分;孙卓等[20]分离自人参根际土壤得到的土壤短波单胞菌对人参锈腐病具有防治作用;吕娜娜等[21]发现成对杆菌属是抑病型蕉园根际土壤的特有菌属。本研究中番茄经绿针假单胞菌PA6以及β-氨基丁酸的诱导处理后,灰霉病病情减轻,番茄根际中短波单胞菌属、成对杆菌属以及鞘脂菌属丰度均有所上升,猜测上述3种菌属和番茄根际的丰富度与调节植物生长、增强植物抗病性有关,是潜在的根际生防菌。
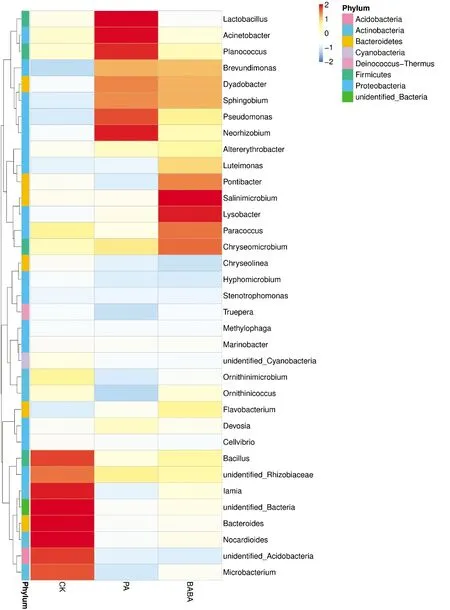
CK:蒸馏水+灰葡萄孢菌;PA:绿针假单孢菌+灰葡萄孢菌;BABA:BABA+灰葡萄孢菌
PA组中丰度较高的还有乳杆菌属(Lactobacillus)、不动杆菌属(Acinetobacter)、动性球菌属(Planococcus)、假单胞菌属(Pseudomonas)以及新根瘤菌属(Neorhizobium)。有研究表明:乳杆菌属(Lactobacillus)通过产生苯乳酸起到果蔬真菌病害的防治作用[22];不动杆菌属(Acinetobacter)可通过提高土壤脱氢酶活性改善土壤活力[23];动性球菌属(Planococcus)可作为根际优势菌提高黑果枸杞等盐生植物适应力[24];假单胞菌属(Pseudomonas)的部分菌属常作为主要的植物根际促生菌[8]。BABA组丰度较高的还有Pontibacter菌属、Salinimicrobium菌属、溶杆菌属(Lysobacter)、副球菌属(Paracoccus)以及Chryseomicrobium菌属。其中溶杆菌属可通过改善根系环境促进大蒜苗期生长[25];Salinimicrobium在盐生植物黑果枸杞的根际高丰度富集可改善植物抗逆性[24]。
3 结论
研究表明绿针假单胞菌PA6及BABA两种诱导因子可降低番茄接种病原菌后的发病率及病情指数,增强抗性基因NPR1、PR1表达量,激发SA信号通路使番茄产生系统抗性,根际菌群分析中门水平的优势菌组成不受诱导处理影响,但门水平及属水平上的优势菌丰度差别显著,根际益生菌群丰度显著上升。郝剑霞等[26]采用2种外源物质(菌种S.fredii及促生剂DSC)研究对大豆农艺特征及根际菌群的影响,发现根际土壤存在功能菌群作为植物营养及相关生化反应来源,外源物质优化了菌群结构进而改善植物生长。与此类似,研究中不同诱导处理引起菌群种类及丰度的明显变化。BABA主要通过激活植物的活性氧保护机制,引起植株抗性成分变化提高植株抗逆性及抗病性[27],假单胞菌属可以激发植物诱导抗性,其本身也可通过根际定植及合成次生抗性代谢物参与根际菌群结构调节,调节植物微生态,间接改善植物生长[9,28]。相比BABA处理,PA处理的防治效果更佳,推测绿针假单胞菌可能直接调整根际功能菌群,其功能菌群与植物抗病性关系更密切,通过植物-微生物互作强化诱导抗病的效果以防治番茄灰霉病。后续将重点分析对应功能菌属引发的番茄根际效应及诱导抗性机制,为番茄灰霉病的生防机制提供理论依据。
