外源CO对采后枣果实链格孢菌浸染过程中蛋白组的影响
2021-06-19于有伟郭毅晶张静茹张少颖
于有伟,郭毅晶,康 琳,王 琼,张静茹,张少颖
(山西师范大学食品科学学院,山西临汾 041004)
枣(ZiziphusjujubaMill.)为鼠李科枣属植物,在我国已经有4000年的栽培历史,枣果实滋味甘甜,口感清脆,含有各种丰富的糖、矿物质、氨基酸、维生素等营养物质,特别是维生素C的含量很高,而且还含有丰富的黄酮、多酚等功能成分,具有养胃、健脾、补血、保肝等医疗保健作用,是一种药食同源的果品[1]。但是鲜枣不耐贮藏,采后极易感染链格孢属(Alternaria)、镰刀菌属(Fusarium)、青霉属(Penicillium)、毛霉属(Mucor)等微生物,其中,由链格孢菌(A.alternata)引发的黑斑病是采后枣果实的主要病害,引起果实腐烂变质,影响货架期,降低了鲜枣的商品价值[2-3],因此,研究枣果实的采后病害防控有重要意义。
近年来,CO在园艺产品采后生理方面的研究表明,通过CO熏蒸可以抑制鲜切莲藕的褐变[4],延缓鲜切月季的衰老[5],保持桃、冬枣等果实的采后品质,延缓果实的采后衰老,延长其货架期[6-8]。高浓度的CO还可以作为杀菌剂,抑制苹果贮藏期间的Botrytiscinerea,Moniliniafructicola和Penicilliumexpansum的生长[9]。王东[10]的研究也表明,CO可能参与了HO-1调控的烟草对烟草花叶病毒的抵御。也有研究表明,低浓度的CO处理在抑制冬枣采后果实的腐烂[7,11]上有积极作用,主要是由于CO处理能通过提高果实抗病相关物质(多酚、黄酮、木质素)的含量和抗病相关蛋白(几丁质酶、β-1,3葡聚糖酶、苯丙氨酸解氨酶、多酚氧化酶)的活性增强枣果实对采后病害的抵御能力。目前,CO在果蔬采后贮藏保鲜方面的研究还不多,CO在果实采后抗病方面的研究还仅停留在其抗病作用的初步探究阶段,其具体的抗病作用机制还有待于进一步的研究。
从蛋白水平入手,利用蛋白质组学的方法探索植物与病原菌相互作用的分子机制,是植物抗病研究的重要手段。目前,对于采后受链格孢菌侵染而患黑斑病的枣果实在蛋白质表达差异方面的研究相对较少,而对外源CO在增强采后枣果实抵抗链格孢菌浸染过程中蛋白差异表达调控的研究还未见报道。因此,为了进一步了解CO处理对枣果实链格孢菌侵染过程中果肉蛋白质差异表达的影响,本研究用CO处理枣果实再对处理后的枣果实接种病原菌,研究外源CO处理后采后枣果实响应黑斑病过程中的蛋白表达及差异蛋白质的功能和调控作用,为进一步解析CO在诱导枣果实采后抗病性中的作用机制奠定基础。
1 材料与方法
1.1 材料与仪器
供试冬枣(沾化冬枣) 采自临汾市尧乡冬枣种植基地,白熟期采收,采摘后迅速运回实验室,选择大小一致、无病虫害和机械损伤的果实作为试验材料;链格孢菌(BNCC115062) 购自北纳生物有限公司;CO气体(纯度99.99%) 购自北京华能特种气体有限公司。
AKTA Purifier 100全自动蛋白层析系统 美国通用电气医疗集团;759S紫外可见分光光度计 上海棱光技术有限公司;MDF-U53V超低温冰箱 日本三洋集团;H1850R台式高速冷冻离心机 湖南湘仪实验室仪器;EASY-nLCTM1200色谱系统和Q-Exactive质谱仪 美国Thermo Scientific公司。
1.2 实验方法
1.2.1 链格孢菌孢子悬浮液的配制 参照He的方法[12],将链格孢菌接种于PDA平板上,28 ℃下培养10 d。用含有0.05% Tween-20的无菌水将孢子从平板上冲洗取出,三层纱布过滤后,显微镜计数,将孢子悬浮液的浓度调整为1×105个/mL。
1.2.2 试验处理及取样 选取60个大小一致、无病虫害和机械损伤的枣果实,用2%的次氯酸钠溶液清洗晾干后,再平均分成2组,每组30个果实。根据前期预实验的CO浓度筛选结果,10 μmol/L CO熏蒸2 h的处理效果最佳,因此,本实验分别进行如下处理:CO+A.alternata处理(记为COA),将果实置于密闭干燥器中用10 μmol/L CO熏蒸2 h,取出后在室温下放置24 h,然后在枣果实的中部接种A.alternate的孢子溶液5μL;A.alternata接种处理(记为A),用打孔器在在赤道处打深 2 mm,直径为3 mm的小孔,再用移液枪向小孔中注入5 μL孢子溶液进行接种。处理后两组果实均置于带孔的塑料盒中于室温下(20±2 ℃,相对湿度75%~85%)贮藏。分别于处理后0、24、48和96 h后,削去枣的表皮后,切取距接种点5 mm的枣果肉10 g用于蛋白组的分析。
1.2.3 蛋白组分析
1.2.3.1 蛋白质提取和肽段酶解 样品采用SDT(100 mmol/L Tris/HCl pH7.6,0.1 mol/L DTT,4%(w/v)SDS)裂解液提取蛋白质[13],用BCA(bicinchoninic acid)法定量后,再用FASP(Filter aided proteome preparation)法进行胰蛋白酶酶解,用C18 Cartridge填料的固相萃取小柱对酶解的肽段脱盐,肽段经冻干、复溶后,测定OD280定量。
1.2.3.2 肽段标记 分别取100 μg多肽样本,按照试剂盒说明(ITRAQ-8plex)标记各个样本。
1.2.3.3 SCX色谱分级 混合标记后的肽段,分级使用全自动蛋白层析系统(AKTA Purifier 100)。色谱柱的平衡液(pH3.0)为25% ACN,10 mmol/L KH2PO4;色谱柱的洗脱液(pH3.0)为500 mmol/L KCl,10 mmol/L KH2PO4,25% CAN。色谱柱平衡后,上样分离,流速1 mL/min。液相梯度如下:0~25 min,洗脱液的线性梯度为0%~10%;25~32 min,为10%~20%;32~42 min,为20%~45%;42~47 min,为45%~100%;47~60 min维持在100%;60 min后,洗脱液重置为0%;边洗脱边检测OD214,洗脱组分的分收集为每2 min/次,各组分冻干后用C18 Cartridge脱盐[13]。
1.2.3.4 LC-MS/MS数据采集 分级样品的分离用HPLC系统Easy nLC(纳升流速)进行[13]。缓冲液:A液为0.1%甲酸水溶液,B液为0.1%甲酸乙腈水溶液。用95% 的A液平衡色谱柱。上样柱为Thermo scientific EASY column(5 μm×100 μm×2 cm,C18),上样后,再经Thermo scientific EASY column分析柱(3 μm×75 μm×10 cm,C18)分离,流速为250 nL/min。液相梯度如下:0~50 min,B液线性梯度从0%~35%;50~58 min,从35%~100%;58~60 min,维持在100%。
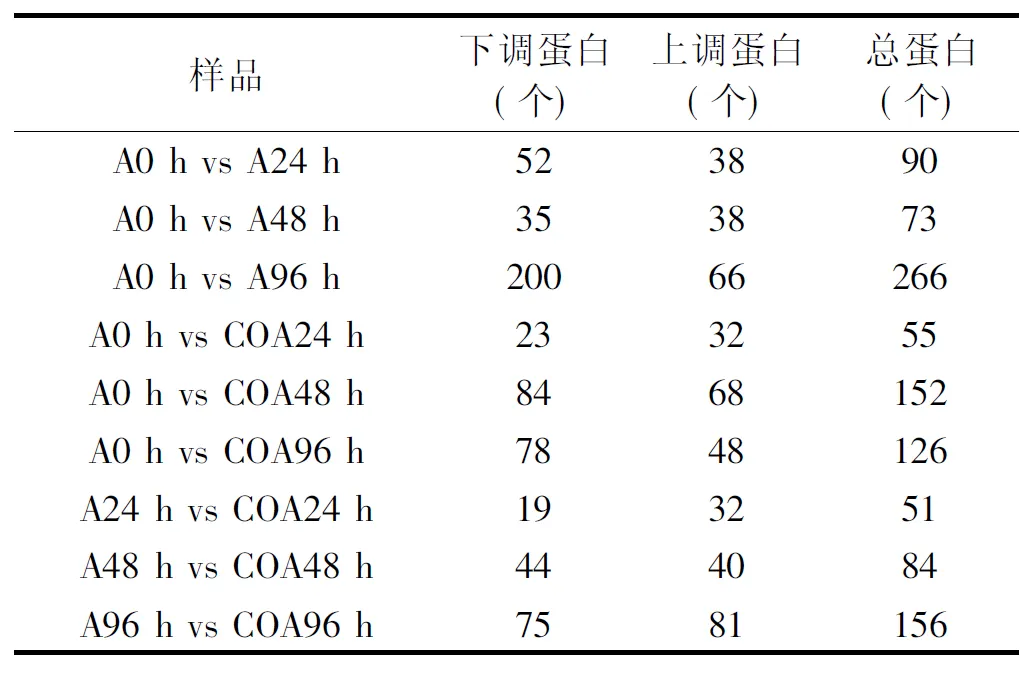
表1 不同时期和不同处理的差异蛋白个数Table 1 Number of differential expressed proteins in different periods and different treatments
质谱分析Q-Exactive质谱仪进行[13],正离子检测,分析时长为60 min,母离子扫描范围300~1800 m/z,一级质谱分辨率为70000 at m/z 200,AGC(Automatic gain control)target为3e6,Maximum IT为10 ms,动态排除时间为40.0 s。按照下列方法采集多肽和多肽的碎片的质荷比:每次全扫描后采集10个碎片图谱(MS2 scan),MS2 Activation Type为HCD,Isolation window为2 m/z,二级质谱分辨率为17500 at 200 m/z,Normalized collision energy为30 eV,Underfill ratio为0.1%。
1.2.3.5 蛋白质鉴定和定量分析 用Proteome Discoverer 1.4检索质谱测试原始数据的相应数据库,再进行蛋白质鉴定及定量分析,以差异倍数值大于1.5倍为显著上调、小于0.667为显著下调为标准。
1.2.3.6 生物信息学分析 GO功能注释利用软件Omicsbean(http://www.omicsbean.cn/)对目标蛋白质集合进行GO注释。KEGG通路注释利用软件Omicsbean对目标蛋白质集合进行KEGG通路注释。
1.3 数据分析
数据的方差分析和绘图选用DPS v14.10和Microsoft Excel 2010进行,差异显著性检验(P<0.05)用t测验。
2 结果与分析
2.1 蛋白质鉴定结果和差异蛋白分析
由表1可以看出,所有样本共鉴定出的肽段数为5730,蛋白为2158;其中差异蛋白总数为1053,上调的差异蛋白数为443,下调的差异蛋白数为610;与0 h相比,接种处理的冬枣果实在24、48和96 h都有不同数量的蛋白质的差异表达蛋白,其中,处理后24、48 h时,上调差异蛋白数分别为38和38,下调差异蛋白数分别为52和35,而96 h后上调和下调的差异蛋白数量分别上升为66和200;在24、48和96 h时,CO+接种处理果实中的蛋白与0 h的相比也出现了不同数量的差异表达,但是从总量上看,除48 h的样品外,24和96 h的差异蛋白数量均低于接种处理的;而相对于接种,CO+接种处理在24、48和96 h时样品的上调差异表达蛋白质数分别为32、40、81,下调差异表达蛋白质数分别为19、44和75。
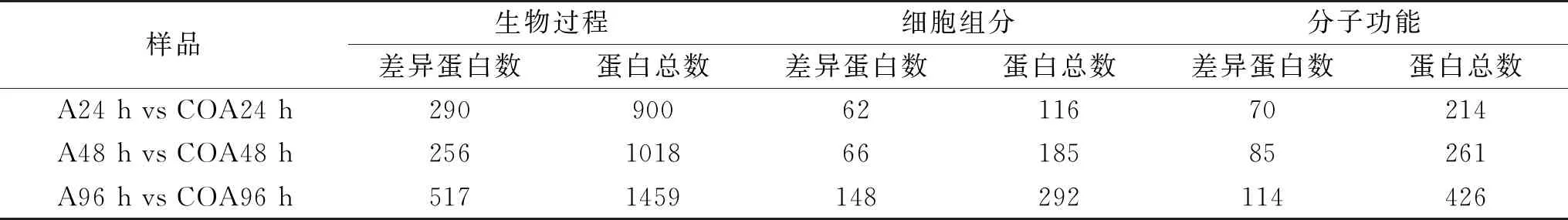
表2 差异蛋白富集统计分析Table 2 Statistical analysis of differential protein enrichment
2.2 差异蛋白的GO分析
如表2所示,相对于单独接种处理,CO+接种处理后随着贮藏时间的延长,在细胞组分、分子功能和生物过程三种类别上富集的总差异蛋白数增大。生物过程上,在48 h富集的差异蛋白达到显著水平的有256个,低于24 h和96 h的蛋白数290和517,而在细胞组分和分子功能上,三个时间点上达到显著水平的差异蛋白数均呈现上升趋势。
为了进一步了解差异蛋白质的GO富集情况,分别选取在细胞组分、分子功能和生物过程三个类别上24、48和96 h 时间点的分析显著性排名前十的条目进行分析(见图1)。细胞组分上(图1A),在24、48和96 h时,两个处理3个时期共有18个类别的显著性富集,而且随贮藏时间延长,各组分的富集量增加且差异更显著,差异蛋白的富集在细胞质膜和细胞器的积累增加,细胞液和细胞质的富集程度在三个时期均较高。分子功能上(图1B),三个时间段上共有14个类别的显著性富集,共有的富集包括脱落酸结合、蛋白结合、类异戊二烯结合、激素结合、乙醇结合、蛋白磷酸酶抑制剂活性和磷酸酶抑制剂活性和单羧酸结合。处理后48 h,差异蛋白的分子功能富集最多。生物过程上(图1C),48和96 h时,有3个显著共同类别富集,包括过单一生物代谢过程、单一生物细胞过程和单一生物过程,而96 h时的差异蛋白生物过程的富集量更大。
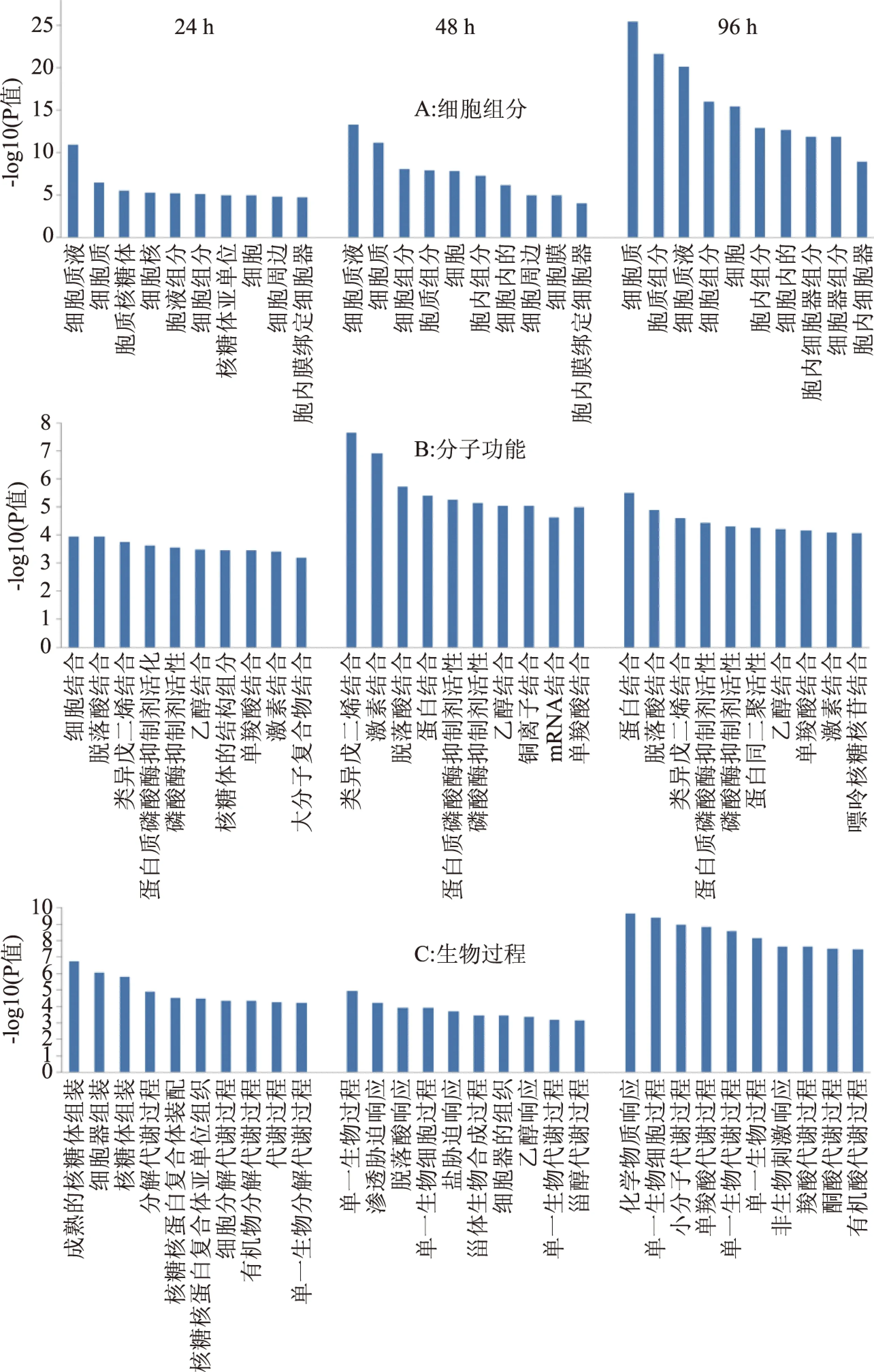
图1 差异蛋白GO富集分析Fig.1 GO enrichment analysis of differential proteins 注:细胞组分(A)、分子功能(B)和细胞过程(C)。
2.3 差异表达蛋白质 KEGG 通路富集分析
运用KEGG数据库对差异蛋白进行代谢途径富集分析。处理后24 h,冬枣果实的差异表达蛋白主要富集在5个代谢途径(图2),其中核糖体、甾体生物合成相互作用途径富集度较高,其后为植物-病原菌、mRNA监视途径、果糖和甘露糖代谢通路;处理后48和96 h差异表达蛋白富集的主要代谢途径明显增多,均为9个。48 h差异蛋白主要富集在鞘糖脂生物合成、植物激素信号转导、氨酰-tRNA生物合成、磷脂酰肌醇信号系统、光合组织中的碳固定、维生素B6和生物素代谢、赖氨酸生物合成和碳代谢通路;96 h差异蛋白富集在甾体生物合成、糖酵解/糖异生、碳代谢、细胞内吞作用、氨基酸生物合成、单萜生物合成、内质网中的蛋白加工、错配修复和亚油酸代谢通路。差异蛋白在48和96 h共同富集的途径为碳代谢,而甾体生物合成途径在24和96 h都有富集。
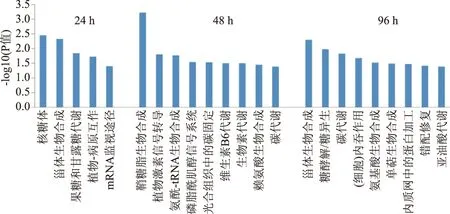
图2 差异表达蛋白的KEGG 通路富集分析Fig.2 KEGG pathway enrichment analysis of differential proteins
2.4 目标差异蛋白
根据差异蛋白GO富集和KEGG途径富集的分析结果,发现经CO处理后,枣果实响应A.alternata过程中的差异蛋白主要富集在5个主要的功能过程,分别为:碳水化合物和能量代谢、氨基酸和脂类代谢、次生代谢产物的生物合成和防御相关蛋白、信号的感知和信号转导、转录和转录后调控。共筛选出72个主要的目标差异蛋白(表3)。筛选出的差异蛋白主要包括:碳水化合物和能量代谢相关的蛋白16个;氨基酸和脂类代谢相关的蛋白12个;次生代谢产物的生物合成和防御相关蛋白19个;信号的感知和信号转导的相关蛋白10个;转录和转录后调控的相关蛋白15个。

表3 差异蛋白功能分类Table 3 The functional classification of DEPs
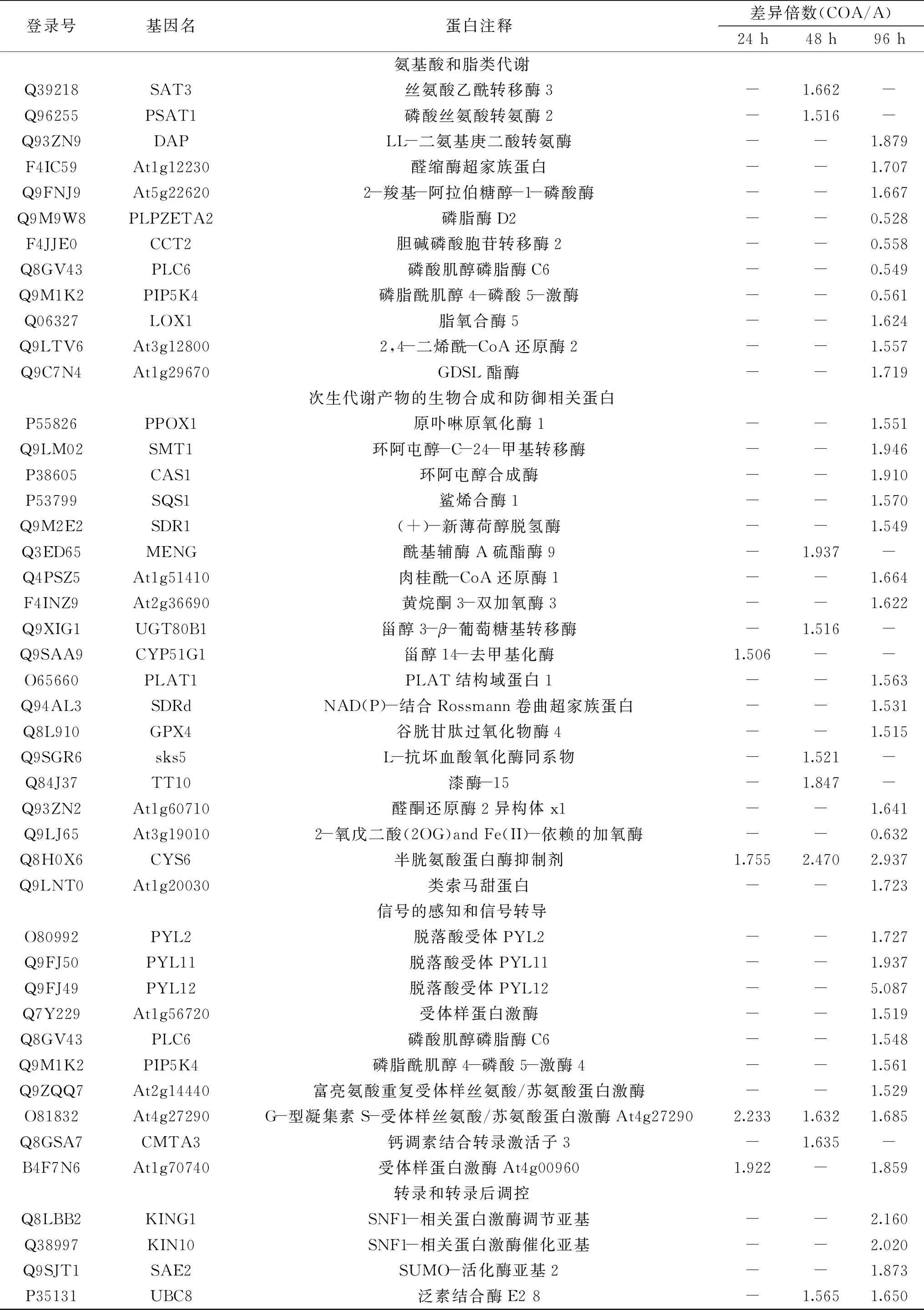
续表

续表
3 讨论
3.1 碳水化合物和能量代谢相关蛋白
有证据表明,植物防御需要大量能量,而产能的初级代谢在植物抗性反应中起着重要作用[14-15]。植物抵御病原菌入侵也会导致能量流失,引发细胞内的碳水化合物代谢增强[16]。本研究筛选出16个与碳水化合物和能量代谢有关的差异蛋白。其中,在接种后96 h内,有5个上调的与能量代谢相关的差异蛋白(P93306、F4HXY8、O82628、Q9FT52和Q5M729),6个与糖酵解及三羧酸循环相关的上调差异蛋白(Q9SN86、Q9M039、Q9C9P3、Q9MB58、Q9C9C4和P25696)以及5个上调的糖苷酶蛋白(Q9SU50、Q8VZJ2、F4IHD3、Q9SCW1和Q9FT97)。Li等[17]的研究表明,外源NO处理可以提高糖酵解和三羧酸循环相关蛋白的表达丰度,从而为桃果实应答采后病原菌提供能量,本研究的结果证明,外源CO处理也有类似的效果。有研究表明,可溶性糖在果实对多种胁迫源的响应中起重要作用[18],海藻糖酶、β-1,3-葡萄糖苷酶和半乳糖苷酶作为糖苷水解酶可以将糖苷键水解生成小分子可溶性糖,三者均在植物抗病过程发挥作用[19-21]。本研究也发现,外源CO处理提高了枣果实响应链格孢菌侵染过程中细胞内的海藻糖酶、2个β-1,3-葡萄糖苷酶和2个半乳糖苷酶的表达量。
3.2 脂类和氨基酸代谢相关蛋白
植物体内的氨基酸是许多其它生物代谢产物的前体物质,而这些代谢产物涉及植物逆境响应等多种功能,是植物免疫系统必需组成部分[22-23]。文景芝等[24]的研究表明,大豆种子分泌物中游离氨基酸在一定程度上可能作为信号分子参与大豆疫霉寄主选择性和寄主抗病性。此外,L-谷氨酸的代谢物GABA、Met、Lys和Arg,也能诱导番茄果实对灰霉病菌产生抗性[25]。本研究结果显示,在枣果实和病原菌相互作用的96 h内,CO处理可以诱导氨基酸代谢相关的5个差异蛋白的上调,涉及His、Lys及Met的生物合成。
脂肪酸和脂类是植物细胞的必需成分,不仅为各种代谢过程提供结构完整性和能量,还可作为信号转导介质[26]。脂类代谢及其代谢产物也参与植物对病原菌胁迫的响应,包括氧化脂类,与脂氧合酶途径(如与JA的合成)有关;不饱和脂肪酸途径,与膜脂类组成和防御信号的感知和转导有关;以及长链脂肪酸途径[27-29]。CO处理后,发现了7个与脂类代谢相关的差异蛋白,其中Q8GV43和Q9M1K2参与磷脂酰肌醇信号调控和磷酸肌醇代谢;Q9M9W8和F4JJE0参与甘油磷脂代谢;Q9LTV6和Q06327参与脂类氧化;Q9C7N4涉及脂肪的水解。有报道表明,Q9LTV6和Q06327参与了植物抗病诱导过程中的JA、SA、Eth和ABA信号响应和调控[30-31]。因此推测,CO可能通过改变枣果实的脂类代谢参与脂类物质介导信号转导,从而抵御病原菌的入侵。
3.3 次生代谢产物的合成和防御相关蛋白
植物可以合成多种次生代谢产物来对抗生物和非生物的胁迫[32-33]。多酚、黄酮、木质素,甾醇及其它萜类物质等次生代谢产物都能帮助植物抵御病原菌的入侵[34]。本研究的结果显示,外源CO处理后,1个卟啉代谢相关蛋白(P55826),9个甾醇、木质素、黄酮醇及其它萜类合成相关蛋白(Q9LM02、P38605、P53799、Q9M2E2、Q3ED65、Q4PSZ5、F4INZ9、Q9XIG1、Q9SAA9)上调,因此,CO可能通过上调次生代谢产物的合成增强枣果实抵御病原菌的入侵的能力。研究发现,病原菌入侵时,果实组织会在短期内产生大量活性氧(ROS),阻止病原菌的侵害[35]。低浓度的ROS也可作为信号分子,诱导防御基因的表达。而当植物细胞内的ROS积累过多时,会导致细胞膜脂过氧化,破坏寄主的防御系统。植物体内的ROS清除系统在保持ROS的动态平衡中起关键作用[36]。本研究在CO处理组的枣果实中发现了7个与ROS平衡或氧还平衡相关的差异蛋白(O65660、Q94AL3、Q8L910、Q9SGR6、Q84J37、Q93ZN2、Q9LJ65);还发现2个上调的抗病相关的差异蛋白Q8H0X6和Q9LNT0。由此可见,外源CO在维持ROS的代谢平衡和提高抗病相关蛋白的表达量上发挥积极作用。
3.4 信号感知和信号转导
在植物对抗病原菌入侵的初期,植物细胞与病原菌的相互识别是关键[37]。植物通过细胞表面的受体分子与病原菌激发子相结合,启动多种防御机制,使细胞对病原菌产生抗性[38]。本研究发现,外源CO处理诱导了Q9ZQQ7、O81832、Q7Y229和B4F7N6的表达。已有研究证实,以上4种蛋白均参与了植物细胞对病原菌激发子的识别,介导下游防卫反应的发生[39-41]。
在植物胁迫应答过程中,CO信号与其他胁迫响应信号网络重叠,且可能通过与这些信号分子协同作用调控胁迫反应[42]。本研究分析了CO诱导枣果实抵抗病原菌侵染过程中涉及的信号转导的差异蛋白,根据这些差异蛋白在信号转导途径中的作用把它们分为与Ca2+信号、磷脂酰肌醇信号、ABA信号及JA信号相关的四个类别。Ca2+作为第二信使,参与了植物细胞与病原菌互作的信号转导过程,而磷脂酰肌醇信号又参与Ca2+信号的调节。本研究发现外源CO处理后,1个Ca2+信号相关的差异蛋白(Q8GSA7)和2个磷脂酰肌醇信号相关差异蛋白(Q8GV43和Q9M1K2)上调,这表明了CO可以通过Ca2+途径调控枣果实对病原菌的防御反应。研究还发现了3个与ABA信号转导相关的蛋白(O80992、Q9FJ50和Q9FJ49)和1个与JA生物合成相关的蛋白(Q06327)。有研究表明,外源ABA可以诱导植物对病原菌的抗性[43-44],而且ABA的受体蛋白O80992可能是一种PR蛋白,参与植物的抗病过程[45]。基于以上分析,我们认为Ca2+、磷脂酰肌醇信号、ABA及JA信号可能参与了CO对枣果实抗病性的诱导。
3.5 转录和转录后调节
基因转录和转录后的调节在植物细胞抵抗病原菌入侵的过程中也发挥着至关重要的作用[17]。本研究发现,CO处理改变了一些与蛋白质合成、加工修饰和降解相关的蛋白表达,包括7个核糖体蛋白、2个SNF1-相关蛋白激酶、3个介导泛素化的蛋白和3个参与蛋白加工的热激蛋白。虽然,有关核糖体参与植物与病原菌互作的报道很少,但有研究表明,核糖体蛋白L12参与了烟草和拟南芥对病原菌胁迫的响应[46],而核糖体蛋白S30也参与了花生对青枯病防御[47]。由此可以推测,CO处理后,核糖体差异蛋白可能参与枣果实对病原菌的防御应答。SNF1-相关蛋白激酶,是一类Ser/Thr蛋白激酶,参与植物的抗病信号调控[48]。本研究中检测到的2个SNF1-相关蛋白激酶均为上调表达,推测其可能在CO诱导的枣果实抗病过程中发挥积极作用。泛素化是植物体内一种广泛存在的细胞反应调控机制,参与植物的抗病调控[49]。本研究中3个介导泛素化蛋白的上调表达也暗示蛋白质的泛素化修饰可能参与了CO诱导的枣果实对黑斑病的防御反应。在本研究中发现的3种热激蛋白在外源CO处理下被鉴定出上调表达。热激蛋白参与植物的抗病[50],Li等[17]也发现,在NO处理诱导的桃果实抗病过程中,果实中的热激蛋白表达上调,这与本研究结果相似。
4 结论
本研究的蛋白组学分析结果显示,CO在枣果实响应A.alternate的早期侵染过程中的作用包括:a.感知病原菌入侵的信号并使转导激活,然后通过转录及转录后的调节来调控下游蛋白的表达;b.次生代谢产物的合成启动并与抗病相关蛋白协同作用;c.碳水化合物、氨基酸和脂类代谢等基础代谢途径重构以响应A.alternate的胁迫。
