规则波作用下刚性挺水植物波生流试验研究
2021-06-08蒋昌波曾思益陈克伦
陈 杰,姚 震,蒋昌波,3,曾思益,陈克伦
(1. 长沙理工大学水利工程学院,湖南 长沙 410114;2.洞庭湖水环境治理与生态修复湖南省重点实验室,湖南 长沙 410114; 3.水沙科学与水灾害防治湖南省重点实验室,湖南 长沙 410114)
在水生群落中,水动力过程会对动植物的物质循环、幼虫分散、固碳过程等产生影响,从而影响群落的生物生产。对沿海水生植物而言,由波生流引起的水生植物流是影响植物周围水体中溶解物质和颗粒物质在生态系统中的交换强度、维持距离以及停留时间的直接动因[1- 2]。研究由波生流引起的水生植物流对于分析水生生态系统循环机制具有重要意义。
近年来,众多学者针对由波生流引起的水生植物流特性开展了大量的试验及理论推导工作。Luhar等[3]基于边界层流动理论提出了沉水植物冠顶波生流理论模型;Abdolahpour等[4- 6]对沉水植物刚柔性以及植物密度进行试验研究,提出了含植物特征参数的沉水植物时均流速最大值预测公式,认为沿海沉水植物冠层与周围环境之间的物质交换可以由植物带顶部的垂直混合速率来控制,以及研究了柔性模型与刚性模型的垂直扩散系数;van Veelen等[7]通过采用不同刚度的植被模型进行物理模型试验,研究了柔性植被摇摆作用对流速结构的影响;Chen和Zou[8]采用欧拉- 拉格朗日混合网格的方法对柔性植物进行数值模拟,研究了植被弯曲刚度对经验阻力系数和惯性系数的影响。从目前国内外研究现状可知,以往由波生流引起的水生植物流的研究对象大多为沉水植物,而对挺水植物的研究更关注于消波特性方面[9- 12],对水生植物流的研究较少。
中国沿海水生生态群落中,红树林生态系统具有较高的生物多样性[13],其生态系统具有强大的汇碳能力,对缓解全球气候变暖有重大意义[14]。本文以红树林为原型,设计具有单茎与簇状根特征的刚性挺水植物波生流试验,并通过与Abdolahpour等[4]的沉水植物试验进行对比分析,对刚性挺水植物的植物特征参数及拖曳力长度尺度进行假设并验证,分析挺水植物茎部对根部波生流的影响作用,并将沉水植物时均流速最大值预测公式拓展至挺水植物适用范围。
1 理论分析
1.1 试验设计簇状挺水植物特征参数
沉水植物几何形状及分布条件决定了植物带内部的水流结构,假设沉水植物带中每株植物对水流结构的作用效应相同,且不考虑边壁效应,并将这种作用进行参数化,则可用植物特征参数lf与lp表示[15]:
λf=Af/AT
(1)
λp=Ap/AT
(2)
式中:λf表示植被群迎水阻力参数;λp表示植被群单元阻力参数;Af为单株沉水植物迎流投影面积;Ap为单株沉水植物垂向投影面积;AT为单株沉水植物下底自由表面面积(植物带所占总面积除以植物带内总株数)。若植物为矩形(正方形)排布,且植株间距为S,每柱沉水植物模型直径为d,高为hr,则该模型中植物特征参数为λf=hrd/(S+d)2、λp=(πd2/4)/ (S+d)2。
对于簇状挺水植物,本研究作以下假设:①单株挺水植物簇状根系中的每个单根对水流结构的作用效应相同,且不考虑边壁效应;②挺水植物在规则波作用下的时均淹没部分等于静水面时的淹没部分,对挺水植物计算植物特征参数时只考虑静水面情况。
基于上述假设,并将下标r表示为根部,s表示为茎部,r+s表示为根茎整体。如图1所示,均匀分布(非正方形)沉水植物中茎部直径为Ds,高为hs。单株植株具有单根数量为n,其中单根直径为Dr,高为hr,单植株间距分别为L与W,单植株根部所占最大直径为D(图1中虚线圆部分),则该挺水植物根部植物特征参数为:

图1 挺水植物排布方式(非正方形)示意Fig.1 Schematic diagram of arrangement (non- square) of submerged plants
(3)
(4)
因此挺水植物根部与茎部总植物特征参数为:
(5)
(6)
1.2 挺水植物根部顶端拖曳力长度尺度
在沉水植物冠顶拖曳力研究中,Belcher等[16]把淹没粗糙体(沉水植物)的冠顶拖曳力与电动力学中的测试电荷产生的作用力进行类比,得出沉水植物的冠顶拖曳力与淹没粗糙体(沉水植物)尾流区域中的流体动量衰减程度有关,并提出了淹没粗糙体(沉水植物)的冠顶拖曳力物理模型:
(7)
式中:fid为冠层单位体积拖曳力,fid=(fix,fiy,fiz);Ui为时空平均流速,Ui=(Ux,Uy,Uz);Ld为冠顶拖曳力长度尺度,与Ui不相关;当在粗糙体外时fid=0.
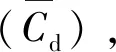
(8)
式中:Ne为淹没粗糙体(沉水植物)区域中粗糙个体的总株数目;Cd为拖曳力系数;β为粗糙体区域中的固体体积分数;hc为粗糙体个体的高度。采用上述植物特征参数λf与λp表示,可得:
(9)
对于簇状挺水植物,本研究作以下假设:当茎密度未达到一定阈值时,挺水植物波生流机制仍以根部作用为主要因素,且茎部作用可以类比为同一植物参数的根部作用。
基于上述假设,并参考Coceal和Belcher[18]在研究平均空气流对城市区域内建筑物顶部作用中,采用体积平均法计算平均建筑物高度。对挺水植物根部与茎部淹没部分采用体积法,建立挺水植物计算模型,可得计算模型高度(hv):
(10)
将式(5)、式(6)和式(9)代入式(10)得挺水植物根部顶端拖曳力长度尺度:
(11)
1.3 沉水植物波生流预测模型
Luhar等[3]认为当波浪经过沉水植物带时产生的拖曳力导致了波浪雷诺应力的产生,且该波浪雷诺应力以及产生的波生时均流速可以用边界层流动理论进行求解,基于线性波理论推导出了沉水植物冠顶波生时均流模型:
(12)

但上述理论公式未考虑沉水植物带密度以及沉水植物植株排布方式等因素,存在一定的局限性。由此Abdolahpour等[4]针对植物带密度以及植物刚柔性开展试验研究,认为沉水植物冠层内任意高度的平均波生流强度与该高度(静水面状态下)的流体质点在振荡运动时的最大速度差(δU)成正比。根据该假设理论,结合试验数据拟合,提出了沉水植物时均流速最大值预测模型:
(13)

(14)
2 试验概述
试验在长沙理工大学波浪水槽内进行,水槽长40.0 m,宽0.5 m,高0.8 m。试验布置如图2所示,水槽左端为一台推板式造波机,另一端设有由多孔吸波材料构成的消能网,以减小波浪反射。

图2 试验布置Fig.2 Experimental setup
在Chen等[19]、He等[20]的试验基础之上,根据学者对海岸红树林的研究成果[21],同时考虑试验条件,本文采用考虑根和茎的挺水植物模型。如图3(a)所示,该模型以新加坡Rhizophora为植物原型,由于该种红树林其树根根须数量、根须直径、对底面投影面积等几何因素非常复杂,本试验仅对植物带根茎部分做概化,并采用变态模型,横向比尺为8∶39、竖向比尺为1∶310。刚性挺水植物茎部、根部采用有机玻璃圆管进行模拟,有机玻璃圆管动态弹性模量(Edy)为4.35×109N/m2、静态弹性模量(Est)为3.15×109N/m2。如图3(b)所示,单株挺水植物的茎部由直径为2.0 cm、高为45.0 cm的圆管组成;根部采用簇状结构分布,由28根圆管组成,每根圆管直径为0.8 cm、高度为14.1 cm,两者在本文试验波浪作用下均不会产生变形,可以较好地模拟刚性挺水植物的根茎部分。为使植物模型固定在水槽底部,安置有机玻璃底板进行辅助固定,底板尺寸长×宽×高为200 cm×49.5 cm×0.8 cm,并按图3(c)所示平面布置方式在底板上进行穿孔。
本试验共设计2种模型,如图3(b)所示,M1为挺水植物根部模型,M2为挺水植物根部与茎部模型,如图3(c)所示,模型各植株间均为矩形分布。考虑实际情况中的波况及试验条件限制,试验水深(h)分别采用30.0 cm和35.0 cm。采用规则波,入射波高(H)分别为6.0 cm、8.0 cm和10.0 cm,入射波周期(T)分别为1.3 s、1.6 s和1.9 s。具体植物模型特征参数及试验工况如表1所示。
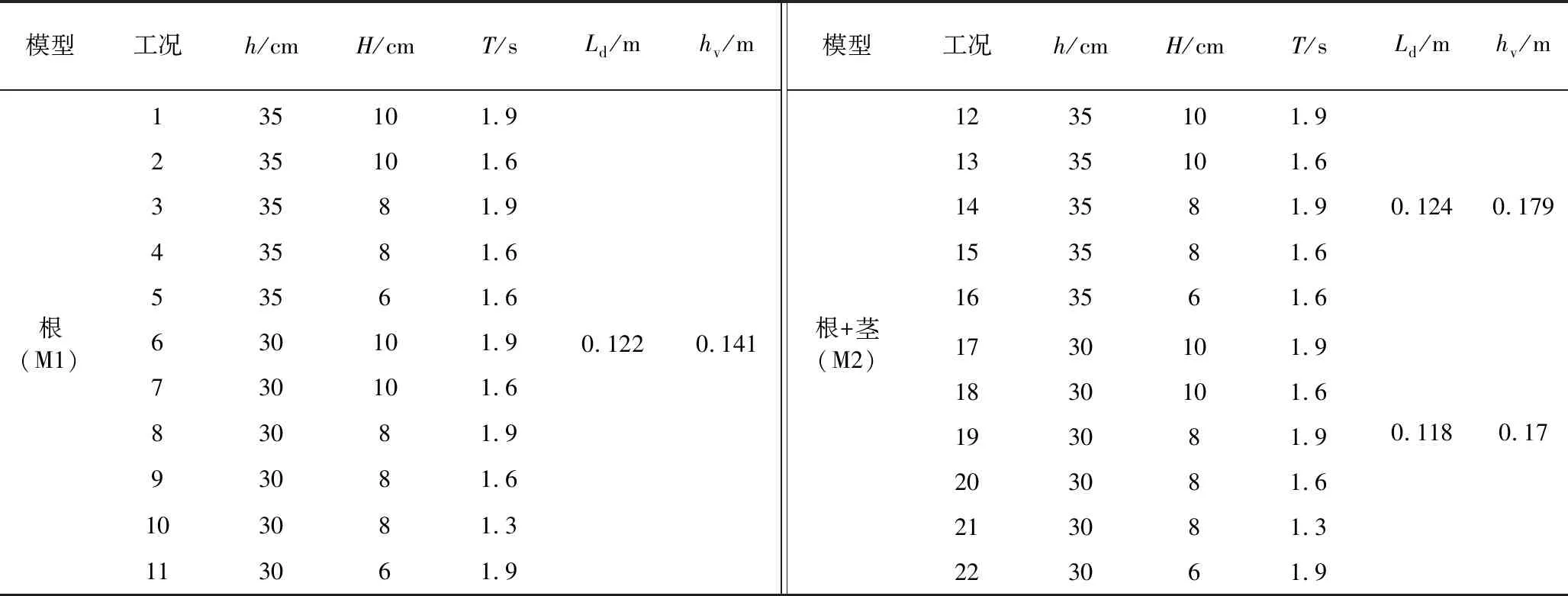
表1 试验工况
试验开始前,进行了大量预备试验,确保造波机的稳定性和重复可靠性。试验时,首先放置植物模型,再摆放仪器设备位置,并标定流速仪测量位置。随后对造波机进行预热,以保证造波机性能稳定。同时开启数据采集系统,并确保浪高仪与流速仪性能良好。调整水位达到试验水深,待水面平静后开始造波,测量波高与流速数据。完成试验数据测量后,停止造波,调整流速仪垂向位置,待水面平静后继续该工况造波。如此直至该工况下整个植物断面流速测量完毕,则为1个工况试验完成。完成1个工况试验后,需待水面平静后,再进行下1个工况造波,重复上述步骤完成下1个工况试验。
如图2所示,在植物带前设置2台ULS 80D超声波浪高仪(G1、G2)测量入射波数据,可通过式(15)计算得到入射波的厄塞尔数(Ur)。
(15)
式中:Lw为波长。
在植物带中,使用三维剖面流速仪(S1)沿垂直剖面测量植物带内流速数据,通过计算均方根流速来探究在植物带内的振荡流强度变化。本试验在计算均方根流速时采用Abdolahpour等[4]提出的通过希尔伯特变换求解均方根流速的方法,如图(4)所示,以剖面流速仪中的1处测量点数据为例,对波传播方向上的流速时间序列数据取10个整周期数据(如图4(a)所示),并对这10个整周期数据进行希尔伯特变换,得到10个整周期内的所有相位速度点,其中包括了平均、湍流、振荡3个分量速度(如图4(b)所示),再通过平均法得到相位平均振荡流速(如图4(c)所示)。因而在任意高度上的波传播方向上均方根振荡流速Urms为
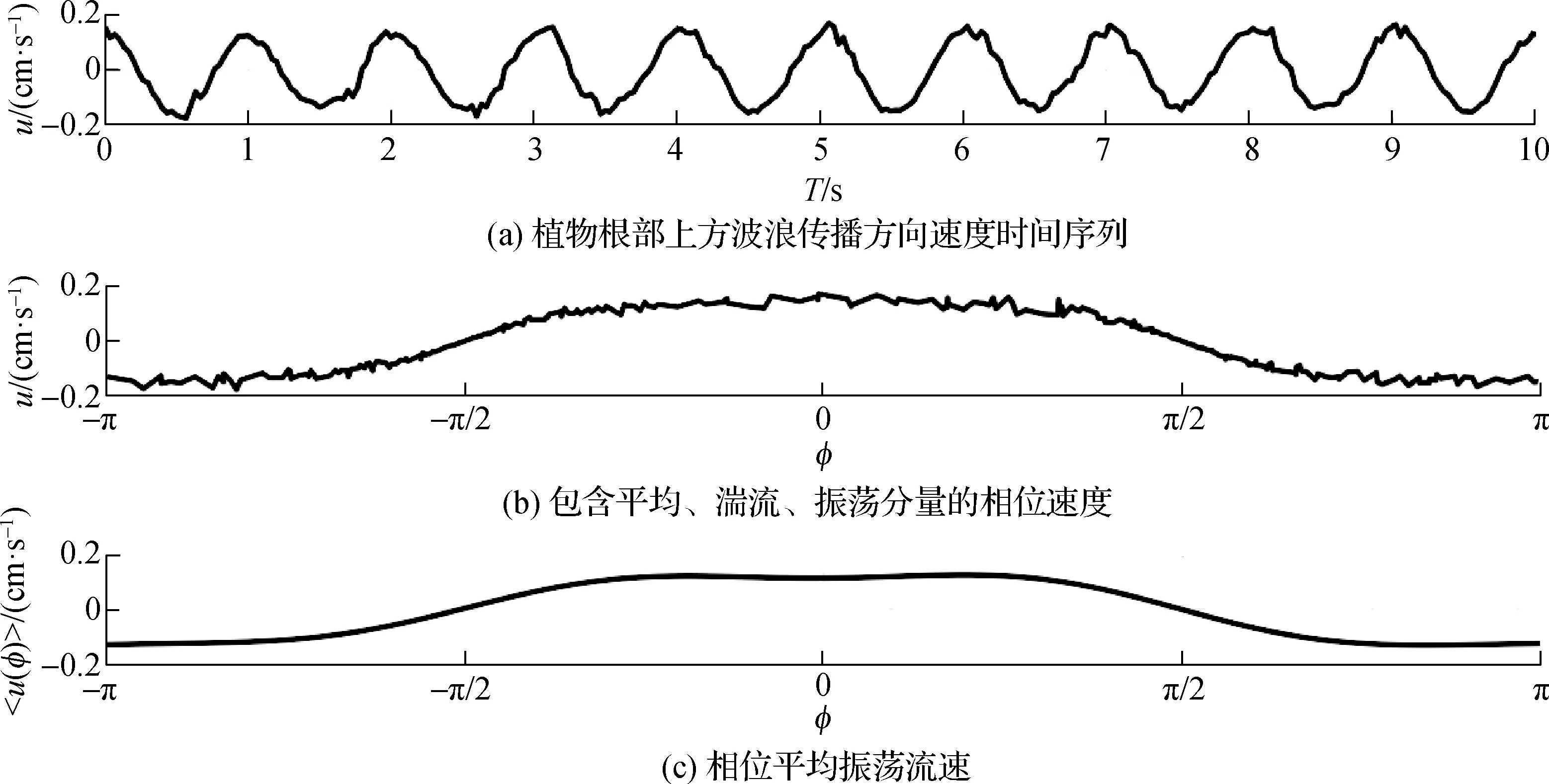
图4 希尔伯特法计算均方根流速(M1- 工况8)Fig.4 Root mean square velocity by Hilbert(Run M1- 8)
(16)
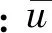
3 结果分析
3.1 挺水植物茎部对均方根振荡流速的影响
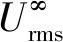
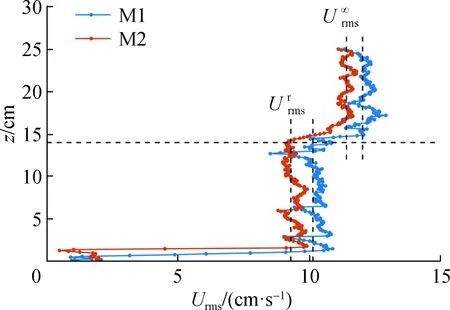
图5 M1与M2模型Urms垂向变化(M1- 工况1、M2- 工况12)Fig.5 Table of relevant parameters of M1 and M2 modelsUrms under the same incoming wave condition
(17)
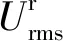
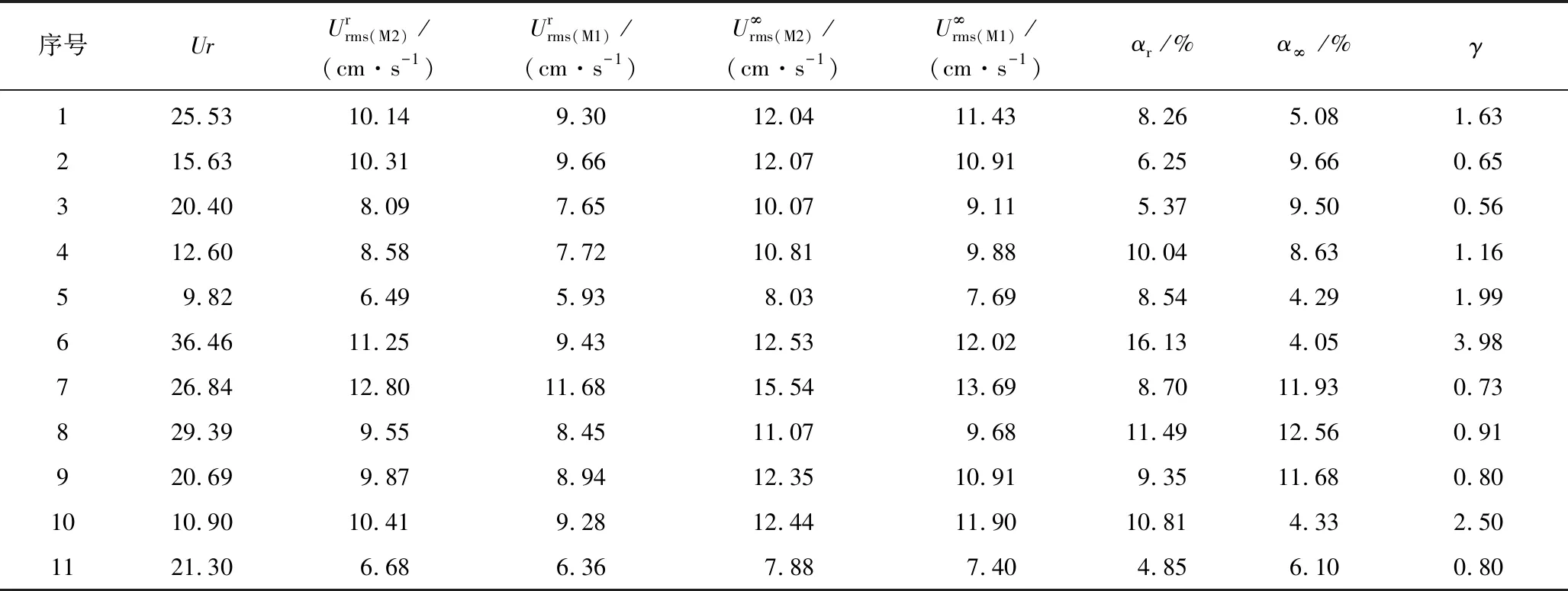
表2 相同来波条件下M1、M2模型Urms相关参数表
(18)
图6给出了根部相对衰减强度(γ)随厄塞尔数(Ur)的变化情况。结果显示γ总体上随Ur增大呈现先减小后增大的趋势。综上所述,挺水植物茎部对Urms起到了衰减作用,且随Ur由小至大,挺水植物茎部主要作用区域从根部区域逐渐转移至根部上方区域,当Ur到超过某一阈值时再增大,主要作用区域从部上方区域再次逐渐转移至根部区域。
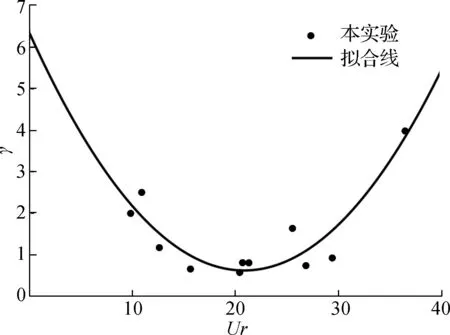
图6 γ与Ur变化关系Fig.6 Relationship between the γ and the Ursell number
对γ与Ur进行非线性回归拟合,可得:
γ=0.013 7Ur2+0.575 2Ur+6.540 1
(19)
式(19)的拟合度R2为0.832 8,表明γ与Ur函数关系较好,说明挺水植物茎部对Urms的主要作用区域随着来波的非线性程度变化而变化。
3.2 挺水植物茎部对均方根流速的影响
图7(a)至图7(k)分别给出了在不同Ur值(如表2中序号1至11所示Ur值)的情况下,挺水植物根模型(M1)与根茎模型(M2)2种模型时均流垂向分布情况。可以看出当Ur在发生变化时,M1与M2模型时均流速最大值所对应的量纲一垂直位置(z-hr)/δT均在[-1,1]的区间内,这与Abdolahpour等[4]在对淹没刚性植物冠层进行试验时结果一致。但Abdolahpour等[4]并未考虑植株排布方式,与本试验采用的簇状排布方式不同,造成本试验中M1模型在不同Ur值的情况下,出现了时均流沿垂向先增大后减小的分布情况,但并未全部出现如Abdolahpour等[4]所提出的平均波生流坍塌现象。
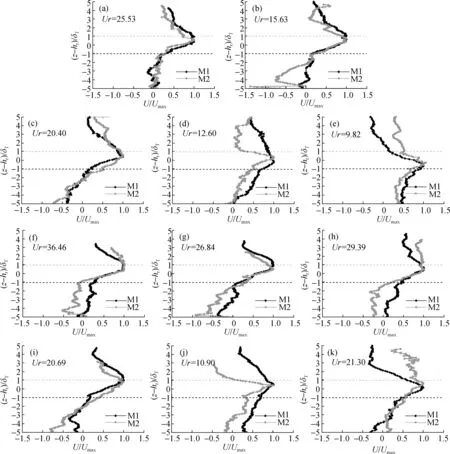
图7 挺水植物根、根茎模型时均流垂向分布Fig.7 Vertical distribution diagram of mean flow in root and stem models of emergent plants
此外,对比M1与M2模型时均流速最大值所对应的量纲一垂直位置随Ur的变化关系可以发现,当Ur值逐渐增大时,M1与M2模型时均流速最大值所对应的量纲一垂直位置沿着某一平均值上下波动。如图8所示:M1模型平均值为15.027 2,方差为0.832 9;M2模型平均值为15.018 1,方差为0.396 0。这表明波浪非线性程度变化会引起时均流速最大值所对应的量纲一垂直位置的波动,且M1模型的波动要明显大于M2模型。综上所述,在来波不同波浪非线性程度即不同水动力条件下,挺水植物茎部对时均波生流垂向分布具有稳定作用,其主要表现在时均流速最大值所对应的量纲一垂直位置。
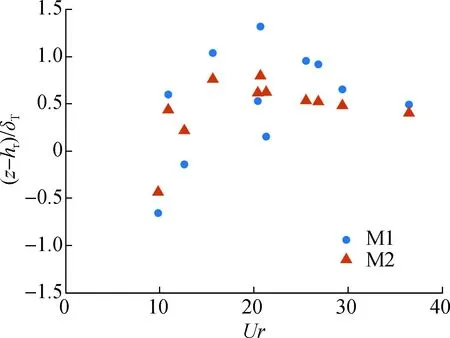
图8 量纲一垂直位置随Ur变化Fig.8 Relationship between the normalized vertical position maximum current speed and the Ursell number
3.3 挺水植物时均流速最大值预测模型
图9分别给出了本试验结果与Abdolahpour等[4]试验结果的时均流速最大值与流体质点经过淹没植物层顶部所经历的流速差关系图。可以看出,本文M1模型与M2模型试验结果与Abdolahpour等[4]试验结果一致。表明当淹没植株(根部)排布为簇状、挺水植物茎部密度为60株/m2时,由植物淹没部分引起的波生时均流强度符合Abdolahpour等[4]提出的理论,即任意高度的时均流强度与以围绕该高度做非规则圆周运动的粒子所经历的速度差成正比,且以时均流速最大值为主要研究对象。因此,当茎密度为60株/m2,挺水植物与具有同一植物特征参数的沉水植物产生的时均流速最大值相同,故与前文假设一致,当挺水茎密度未达到一定阈值时,挺水植物波生流机制仍以根部作用为主要因素,且茎部作用可以类比为同一植物特征参数的根部作用。

图9 时均流速最大值与流体质点经历速度差关系Fig.9 Relationship between the maximum of the temporal- averaged flow velocity and the velocity differential experienced by particles that encounter the top of the root
将挺水植物计算模型相关参数代入Abdolahpour等[4]提出的预测模型式(13)计算预测值,结果显示本试验结果与预测值的拟合度R2为0.766 9。考虑到本文的植株根系分布方式为簇状分布与Abdolahpour等[4]不同,因此基于式(13)进行非线性回归拟合,对式中的系数项进行修正,可由式(20)表示:
(20)
如图10所示,修正后式(20)的拟合度较高且拟合度R2为0.881。值得指出的是式(20)是本文通过对变态模型试验分析得出。所以该处理方式可能会导致式(20)用于实际植物时,存在一定的不确定因素。
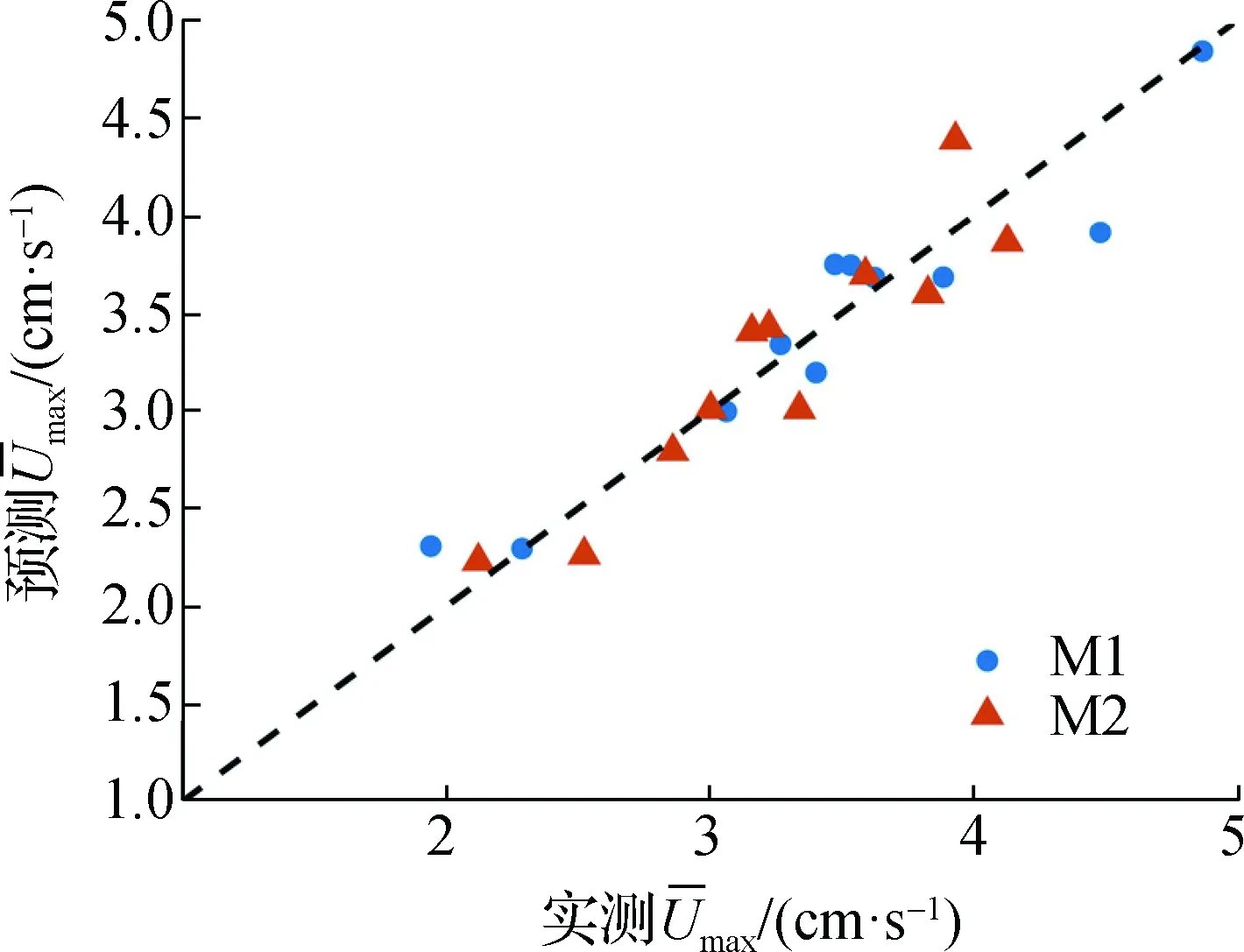
图10 簇状挺水植物根、根茎模型的时均流速最大值预测模型参数修正Fig.10 Parameter modification of the in the root and rhizome models of rigid cluster emergent plants
4 结 论
基于物理模型试验,探究了规则波作用下挺水植物波生流特性。试验采用根茎形式与根形式2种概化模型进行对比,主要结论如下:
(1) 挺水植物茎部对均方根振荡流速具有衰减作用。主要作用区域随着来波的非线性程度增大而上下移动,先从根部区域逐渐移动至根部上方区域,再从根部上方区域逐渐返回至根部区域。并结合本试验数据,提出了挺水植物茎部的主要作用区域预测公式。
(2) 挺水植物茎部对时均流垂向分布具有稳定作用。挺水植物在不同水动力环境下,茎部可以稳定根系内的时均流速分布,起到了保护根系及其附属生物的作用。
(3) 与前人研究结果进行对比,发现挺水植物产生的波生流机理与沉水植物类似,当挺水茎密度未达到一定阈值时,挺水植物波生流仍以根部作用为主要因素,茎部作用可以类比为同一植物参数的根部作用。通过体积平均法建立挺水植物计算模型,对沉水植物预测公式进行系数项修正,提出当植株根系分布为簇状且茎密度为60株/m2时,挺水植物时均流速最大值预测模型。
