云南小驳骨丛枝植原体的分子鉴定及相关基因序列分析*
2021-04-07许杏萍杨子祥刘俊男万琼莲
许杏萍,苏 帆,杨子祥,刘俊男,万琼莲,蔡 红
(1.云南农业大学,农业生物多样性与病虫害控制教育部重点实验室,云南 昆明 650201;2.云南省农业科学院 热区生态农业研究所,云南 元谋 651300;3.玉溪师范学院 化学生物与环境学院,云南 玉溪 653100)
植原体(phytoplasma)是一类非螺旋状、无细胞壁且专性寄生于韧皮部的植物病原微生物。植原体主要依靠叶蝉和飞虱等媒介昆虫进行传播,亦可由植物营养繁殖材料和菟丝子等方式进行传播,感病植株通常表现为丛枝、花变叶、黄化、矮化和节间缩短等症状,目前植原体可危害1 000 多种植物,给多种经济作物的生产造成了严重影响[1-3]。
植原体的系统发育进化关系及分类鉴定主要依赖于高度保守的16S rRNA基因序列的遗传变异性[4-5]。目前基于16S rRNA基因序列RFLP分析的相似系数对植原体株系进行划分,已确定了36 个16Sr 组和150 多个亚组[6-7]。然而在植原体分类鉴定体系中,以高度保守的16S rDNA序列信息作为唯一的系统发育依据,在鉴定亲缘关系密切的植原体株系上存在缺陷且难以对组内植原体株系进行更精确的分类鉴定[8]。为解决植原体分类学的局限性,研究人员开始利用保守性相对较低的非核糖体基因,如核糖体蛋白(rp)基因和secY基因等作为辅助分类的分子遗传标记,有助于更准确地对植原体16Sr 组内株系的真实系统发育关系进行更精细的划分,可进一步将植原体16Sr 组划分为多个不同的亚组[9-12]。
小驳骨(Gendarussa vulgarisNees)又名驳骨草、接骨草等,属于双子叶植物纲唇形目爵床科(Acanthaceae)驳骨草属(GendarussaNees),具有极高的药用价值[13-14]。2019 年,课题组在元谋地区发现的疑似感染植原体而诱发的丛枝症状小驳骨病株,其植株生长缓慢、矮化、节间缩短、腋芽丛生为簇状并且大部分叶片萎缩变小并黄化,这将会降低小驳骨的产质量和药用价值。
基于作为系统发育分子标记的rp和secY基因具有能够高度准确地辨别16Sr 组中亚组间的系统发育关系的能力,同时利用多个系统发育参数可以更好地识别和定义植原体株系。本研究将采用PCR 技术等研究方法对小驳骨植原体16S rRNA、rp及secY基因进行扩增及序列测定,根据这3 个基因的序列信息构建系统发育树进行系统发育关系的比较分析,最终从亚组水平上准确高效地鉴定小驳骨丛枝植原体株系的分类地位。同时通过对secY 蛋白亚基的蛋白特性、蛋白跨膜结构及信号肽进行分析,从蛋白分泌途径初步了解植原体病害的致病机理,为研究小驳骨植原体病害的发生及防治提供理论支持。
1 材料与方法
1.1 试验材料与主要试剂
自然表现丛枝症状的小驳骨植株采自云南省元谋县,并保存于实验室,并以实验室保存的苦楝丛枝植原体病株为阳性对照。
新型快速植物基因DNA 提取试剂盒及PCR产物纯化试剂盒由百泰克生物技术有限公司提供;DNA Marker、TransStart®FastPfu DNA Polymerase 和克隆载体试剂盒pEASY®-Blunt Simple Clonging Kit 均来自北京全式金生物技术有限公司;引物由硕擎生物科技有限公司合成。
1.2 方法
1.2.1 感病小驳骨植株总DNA 提取和PCR 扩增
选取感病小驳骨植株幼嫩叶片进行总DNA提取,提取总DNA 步骤参照新型快速植物基因DNA 提取试剂盒说明书。
以小驳骨病样总DNA 为模板,参照PÉREZLÓPEZ 等[7]和LEE 等[8]设计的植原体通用引物对P1/P7 和R16F2n/R16R2 对植原体16S rRNA基因通过巢式PCR 进行扩增,同时采用引物对rp (II) F1/rp (II) R1、secY-F/secY-R 分别对rp和secY基因进行直接PCR 扩增。引物合成序列见表1。16S rRNA及rp基因的PCR 扩增反应条件参考万琼莲等[15]的研究;secY基因的PCR 扩增反应条件为95 ℃ 2 min,95 ℃ 20 s,45 ℃ 20 s,72 ℃ 40 s,35 cycle,72 ℃延伸 5 min。
1.2.216S rRNA、rp和secY基因序列的克隆及测定
PCR 扩增产物在1%琼脂糖凝胶中进行电泳检测,对PCR 产物进行纯化,并与载体pEASY®-Blunt Simple Cloning Vector 连接转化到化学感受态细胞Trans1-T1 Phage Resistant 中,37 ℃培养1 h 后取100 μL 均匀涂布于经IPTG 和X-gal 处理过的LB 培养基平板上,过夜培养,第2 天进行蓝白斑的挑选以及菌液PCR 的阳性克隆子检测,将含有目的基因片段的菌液送生工测序公司进行序列测定。
1.2.316S rRNA、rp及secY基因序列分析
将测序所获得的16S rRNA、rp及secY基因的核苷酸序列用Vector NTI 11.5.3.软件去除序列两端的载体,以确定测序结果的准确性。利用NCBI 中的ORF Finder (https://www.ncbi.nlm.nih.gov/orffinder/)寻找rp和secY序列的最大开放阅读框并进行分析。将序列提交到NCBI 中BLAST比对确认,并进行同源序列检索;采用MUSCLE(https://www.ebi.ac.uk/Tools/msa/muscle/)对序列进行在线同源性比对,分析基于16SrII 组各植原体16S rRNA、rp及secY基因序列的亲缘关系。利用MEGA X 软件,以邻接法(neighbour-joining)构建分别基于16S rRNA、rp及secY基因的系统进化树以便进行系统发育关系的比较分析,确定小驳骨丛枝植原体株系的分类地位。16S rRNA基因序列通过植原体在线分析网站iPhyClassifier (https://plantpathology.ba.ars.usd a.gov/cgibin/resource/iphyclassifier.cgi)进行虚拟RFLP 分析鉴定,明确该株系的候选种分类地位。相关植原体株系信息如表2。
1.2.4secY基因编码的蛋白亚基的蛋白特性分析及结构预测
分别通过生物信息学在线分析网站Prot-Param (http://web.expasy.org/protparam/)、Protscale (https://web.expasy.org/protscale/)、TMHMM(http://www.cbs.dtu.dk/services/TMHMM-2.0/)和SignalP (http://www.cbs.dtu.dk/ services/SignalP/)对secY基因编码的蛋白特性、蛋白亲疏水性、蛋白跨膜结构区和信号肽进行分析。
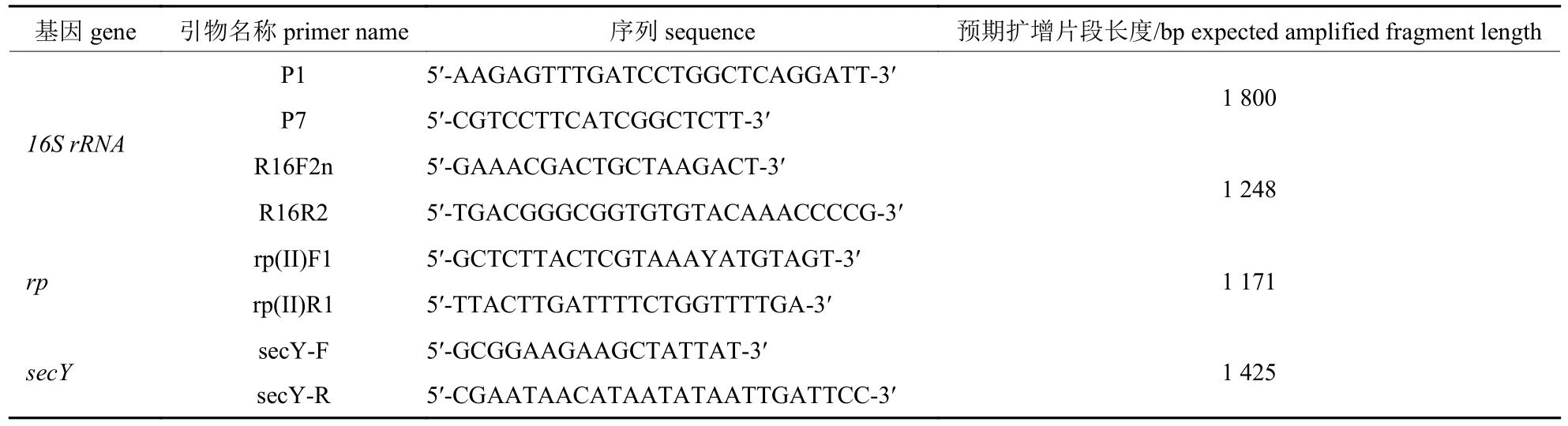
表1 引物序列表Tab.1 Primer sequence listing
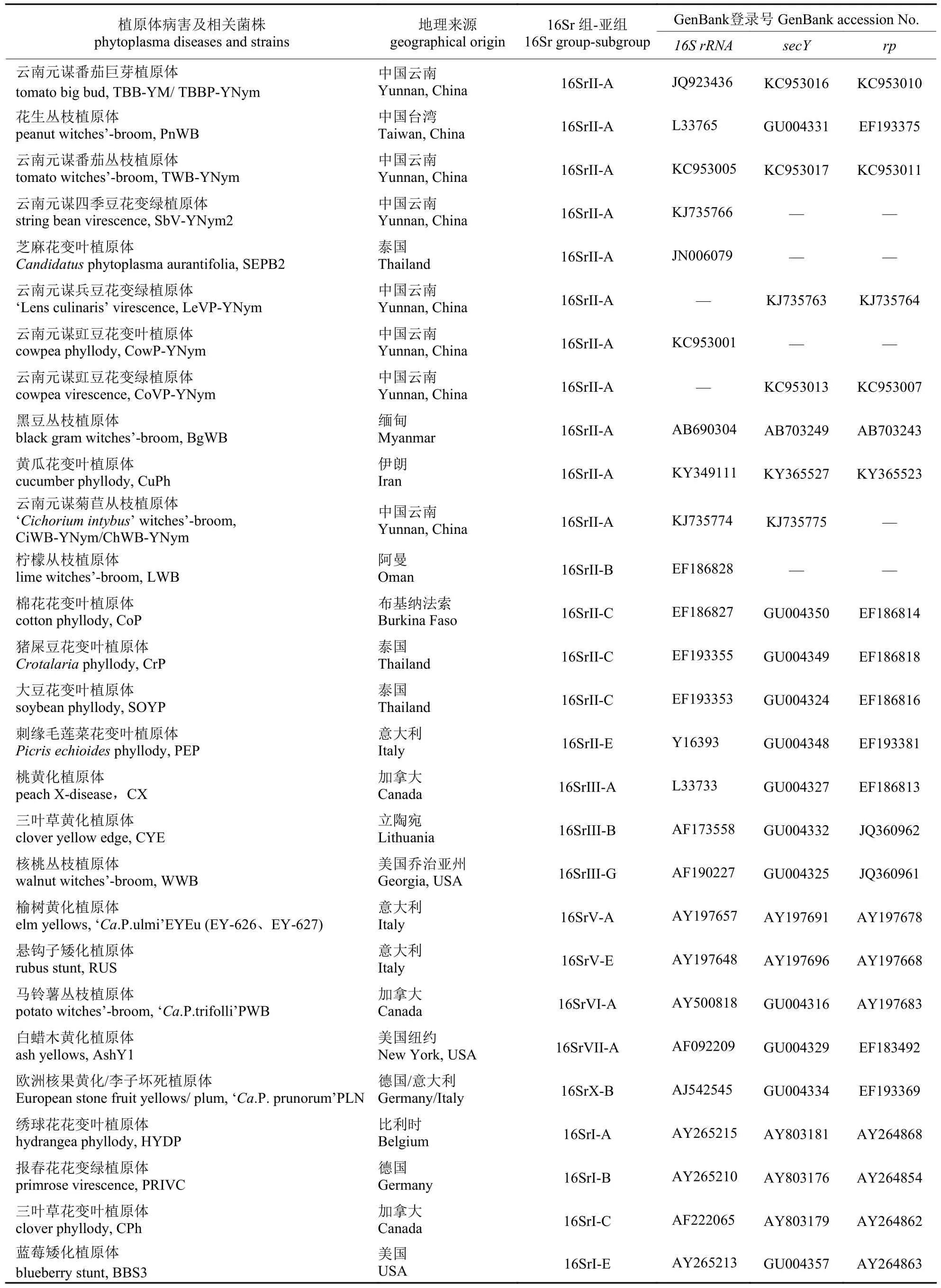
表2 植原体各株系16S rRNA、rp 和secY 基因序列登录号Tab.2 The accession No.of 16S rRNA,rp gene and secY gene sequence of phytoplasma strain
2 结果与分析
2.1 小驳骨丛枝病症状
由图1 所示:感病小驳骨植株严重矮化,节间明显缩短,侧芽细弱,枝条丛生成簇状,叶片小而黄。

图1 云南元谋小驳骨丛枝病症状Fig.1 The symptom of G.vulgaris Nees witches’-broom disease of Yuanmou County,Yunnan Province
2.2 序列结果分析
2.2.116S rRNA、rp和secY基因序列扩增结果
小驳骨感病植株总DNA 经PCR 扩增,均得到大小与阳性对照一致的特异性条带,而双蒸水和健康植株总DNA 均未扩增出特异性条带。测序结果表明:从小驳骨感病植株样品总DNA中分别扩增得到了1 248 bp 的16S rRNA基因片段(nested PCR) (MN535183)、1 171 bp 的rp基因片段(MN543078)及1 425 bp 的secY基因片段(MN543069),这与预期扩增片段长度相符。由此将该植原体株系命名为云南元谋小驳骨丛枝植原体 (Gendarussa vulgariswitches ’-broom phytoplasma,GvWB-YNym)。
2.2.2 小驳骨丛枝植原体(GvWB-YNym)16S rRNA、rp以及secY基因序列分析
(1)16S rRNA基因序列分析
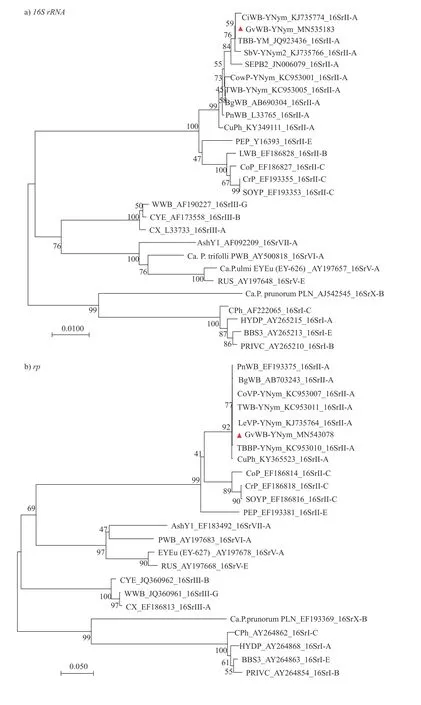
经BLAST 搜索,显示GvWB-YNym 的16S rRNA基因序列与16SrII-A 亚组为最佳匹配同源序列。经MUSCLE 进行在线比对结果(表3)所示:该序列片段与16SrII-A 亚组的番茄巨芽植原体(Tomato big bud,TBB-YM,GenBank 登录号:JQ923436)的相似率高达100.00%。由图2 所示:GvWB-YNym 与植原体16SrII 组明显聚为一枝,且又与归属为16SrII-A 亚组的番茄巨芽植原体(Tomato big bud,TBB-YM,GenBank 登录号:JQ923436)及来自云南元谋的菊苣丛枝植原体(‘Cichorium intybus’ witches’-broom,CiWB-YNym,GenBank 登录号:KJ735774)处于同一枝。
(2)rp基因序列的分析
将测序所得的rp基因序列提交到NCBI 网站ORF Finder 进行在线分析,分析结果表明:rp序列包含了rps3(nt550-1170)基因部分序列和rpl22(nt90-476)基因全部序列,分别编码了206 和128 个氨基酸。由表3 所示:该序列与16SrIIA 亚组各株系相似率均高于99.74%,且与16SrIIA 亚组中的番茄丛枝(TWB-YNym)相似率高达100%。由图2 所示:GvWB-YNym 与16SrIIA 亚组同处1 个分枝簇上,且与16SrII 组聚为一大进化枝,这与基于16S rRNA基因序列构建的系统发育树具有相近之处,表明植原体类群之间具有相似的关联性。因此,依据rp基因序列可将GvWB-YNym 归为16SrII-A 亚组。
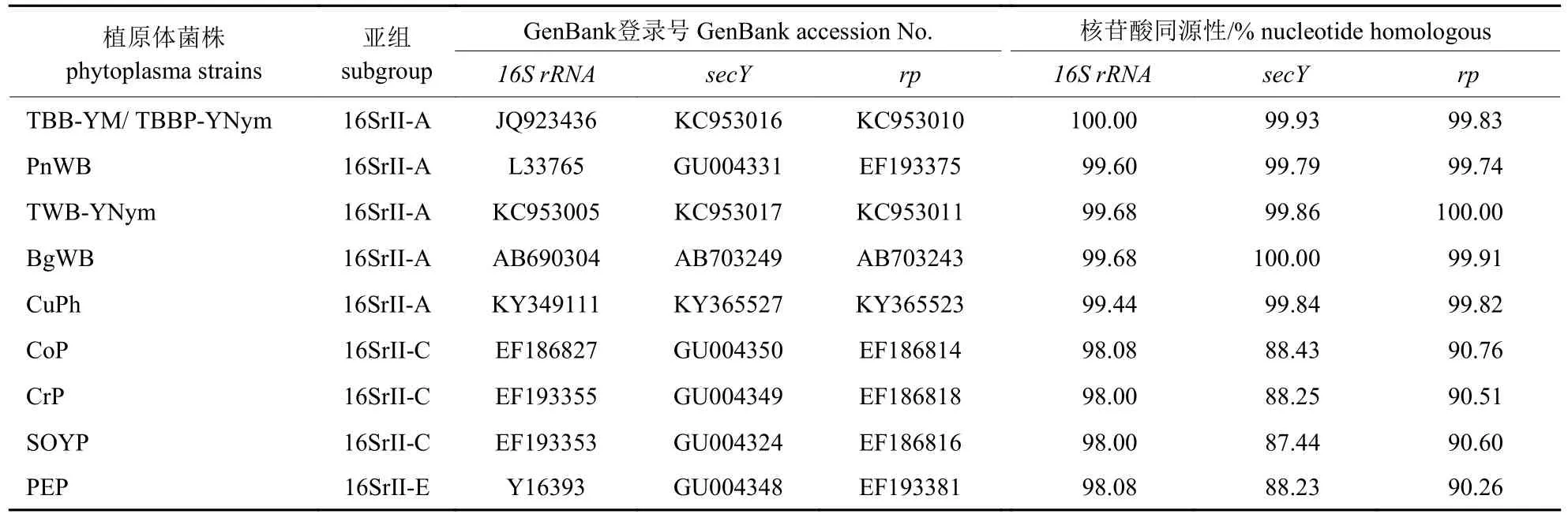
表3 小驳骨丛枝植原体(GvWB-YNym)16S rRNA、rp 和secY 基因核苷酸序列与部分16SrII 亚组株系的同源性分析Tab.3 Homology analysis of nucleotide sequences between GvWB-YNym 16S rRNA,rp and secY gene with other phytoplasma strains of 16SrII subgroup
(3)secY基因序列的分析
将secY基因序列提交至NCBI 网站ORF Finder 进行在线分析,结果显示:secY基因序列包含了整个secY基因(nt74-1336),编码了420 个氨基酸。由表3 所示:GvWB-YNym 与植原体16SrII-A 亚组中的5 个植原体相似率达99.79%以上,与16SrII-A 亚组中的印度黑豆丛枝(Bg-WB)相似率达到100%。由图2 所示:secY基因与16SrII-A 亚组的各个成员共同聚为一群,而与16SrII 组其他亚组及其他外组亲缘关系相对较远;这也与基于rp基因和16S rRNA基因构建的系统发育树显示出相似的主次分枝顺序。据此结果推断,将GvWB-YNym 划分为16SrII-A 亚组。
2.2.3 小驳骨丛枝植原体(GvWB-YNym)16S rRNA基因在线分类鉴定结果
经iPhyClassifier 在线分类鉴定结果显示:该株系16S rRNA基因序列与16SrII-A 亚组代表性植原体花生丛枝植原体株系(L33765)相似性最高,相似系数高达1.00,且二者间的16S rDNA F2n/R2 序列片段模拟RFLP 图谱完全一致,上述所有分类结果均显示GvWB-YNym 归属为16SrⅡ-A亚组。同时,在线分析还显示:GvWB-YNym 与候选种‘CandidatusPhytoplasma aurantifolia’(U15442)同源性最高,达98.2%,因此可确定GvWB-YNym候选种与‘Ca.Phytoplasma aurantifolia’相关。
2.2.4secY基因编码的蛋白亚基生物信息学分析
经分析工具ProtParam 对GvWB-YNym 的secY 蛋白理化特性进行分析,结果显示其编码了420 个氨基酸,分子量为47 720.21 u,理论等电点9.68,脂肪系数为136.45,不稳定系数31.94,属于稳定蛋白,总平均亲水性数值为0.664,表明该蛋白疏水性较好,属疏水性蛋白。通过在线分析网站Protscale 对secY 蛋白亲疏水性作进一步分析,结果如图3 所示。从整体来看,该蛋白疏水氨基酸(正值)多于亲水氨基酸(负值),表现为疏水性,与物理特性预测结果一致;其在215 和216 位氨基酸疏水性分值最高(3.661),疏水性最强;在238 和239 位氨基酸亲水性分值最低(-2.678),亲水性最强。
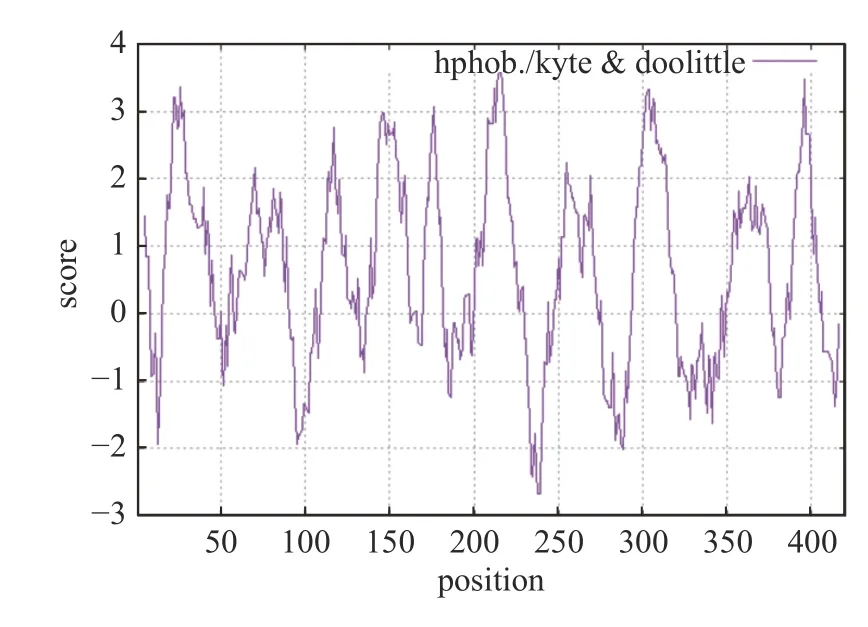
图3 secY 蛋白亲疏水性分析Fig.3 Hydrophilic analysis of secY protein
以在线跨膜结构区预测软件TMHMM 分析secY 蛋白的跨膜域,结果(图4a)显示:该蛋白氨基酸序列具有10 个显著且分布相对均匀的跨膜螺旋区域,表明该蛋白可能定位于细胞中与膜相关的结构,属于跨膜蛋白。信号肽是负责把蛋白引导到不同膜结构的亚细胞器内的短肽链,通过SignalP 在线分析工具对secY基因进行信号肽分析,分析结果如图4b 所示:GvWB-YNym 的secY 蛋白不存在信号肽。
3 讨论
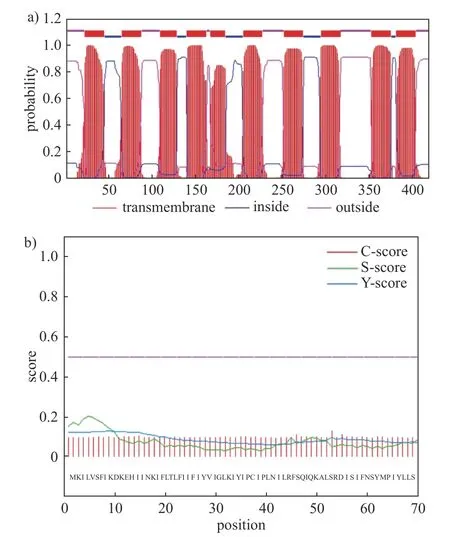
图4 secY 蛋白跨膜结构及信号肽分析Fig.4 TMHMM posterior probabilities and prediction of the signal peptide of secY protein
由于植原体难以体外培养,多方面研究均落后于其他细菌,目前植原体的分类鉴定及其亲缘进化关系的研究领域仍然是热点。高度保守的16S rRNA基因序列常被用作植原体的系统发育进化关系及分类鉴定遗传标记[5]。随着计算机技术及网络系统的加速发展和日渐成熟完善,16S rRNA基因的虚拟RFLP 图谱分析在植原体快速分类鉴定以及流行病学研究的分类学中越来越重要[16]。根据IRPCM (2004)规定[17],16S rRNA基因序列与现有序列间的同源性大于97.5%时可被认为是相同组的植原体。然而,基于植原体中高度保守的16S rRNA基因序列的2.5%不同的任意阈值对植原体株系进行分组的依据,不能满足16S rRNA基因序列同源性虽大于97.5%但在植物寄主范围和介体昆虫等方面具有独特生态学和生物学的植原体株系进行分类的要求。因此,为了能在分子水平上更准确地区分植原体株系,便需要引入额外的分子标记。而作为辅助分类的其他遗传标记如核糖体蛋白(rp)基因、secY、tuf和23S rRNA基因以及16S-23S rRNA基因间隔区序列已被广泛应用。
比较分析给定的PnWB 组(16SrII)各亚组之间的同源性发现:secY基因的序列相似性变化范围最大,其次为rp基因和16S rRNA基因;GvWB-YNym 与16SrII-A 亚组的亲缘关系最为接近,16S rRNA和rp基因序列在16SrII-A 亚组各株系上表现出高相似性,但rp及secY基因序列比16S rRNA基因序列能够呈现出更大的遗传变异性,而secY基因相较于rp基因的序列又具有更大的可变程度,这些信息能够极大地增强对植原体组内遗传进化关系密切但不同亚组的分类鉴定能力,且与前人的研究[12,18-22]具有一致性。通过基于16S rRNA、rp及secY基因构建的系统发育进化树分析,也能够支持这一论断。根据16S rRNA、rp及secY基因序列构建的系统发育进化树,都显示GvWB-YNym 与16SrII-A 亚组的各植原体株系形成单系群,且与其他16SrII 组植原体聚为一大亚枝,但与16SrII 组外的其他亚组植原体明显划分为不同进化枝。这3 个系统进化树分析能够显著区分开植原体株系组与亚组之间的关系,进一步证实了以rp及secY基因作为分类辅助标记的可信度和重要性,是对利用16S rRNA基因对植原体进行分类的补充依据,也表明了GvWB-YNym 归属到16SrII-A 亚组的准确性。然而,分别基于16S rRNA、rp及secY基因构建的系统进化树中的各个分枝顺序并不完全一致,表明16S rRNA、rp及secY基因在其各自的进化史上遗传变异程度的不一致性。16S rDNA序列的RFLP 分析代表不同的系统发育谱系,依据植原体在线分类网站iPhyClassifier 对GvWB-YNym进行快速分类,结果表明GvWB-YNym 与候选种‘Ca.Phytoplasma aurantifolia’相关,且与16SrII-A亚组代表性植原体株系花生丛枝植原体(L33765)的16S rDNAF2n/R2 序列片段模拟RFLP 图谱完全一致,进一步证实了将GvWB-YNym 归属为16SrII-A 亚组分类地位的准确性。这是小驳骨丛枝植原体的首次报道。
原核生物细胞的生长和发育依赖于Sec 分泌蛋白转运机制,多数分泌蛋白和整合膜蛋白都需要经由此途径进行转运,其中起重要作用的是膜蛋白三聚体SecYEG 和动力蛋白secA。在Sec 转运酶中,secY、secE、secG 和secA 构成作为细胞质膜的输出机制中的转位酶复合物,secA、secE 和secY 是蛋白质易位和细胞活力必不可少的部分。在蛋白转运过程中,前体蛋白经由secA 携带的伴胞分子的引导,同时利用ATP 酶的水解活性促进该蛋白质穿过三聚体SecYEG 形成的跨膜通道到达膜外[23]。植原体有2 个分泌系统,YidC 和Sec,后者几乎是绝大多数或所有植原体的共同分泌系统,KAKIZAWA 等[24]发现:植原体中存在Sec 系统,而secA 和secY 蛋白是其中重要组成部分,但其并不清楚Sec 系统在植原体中的作用机制。岳红妮等[25]研究发现:泡桐丛枝(PaWB)植原体中同样存在的Sec 分泌蛋白转运系统,并猜测该系统可能会直接将细菌蛋白质如毒素等转运到宿主细胞质或寄主昆虫细胞中,从而诱发宿主病症。本研究通过对secY 蛋白进行生物信息参数的分析发现:该蛋白作为疏水性稳定跨膜蛋白存在于感病小驳骨植株中;secY 蛋白含10 个显著疏水跨膜区域,与大肠杆菌中的secY 蛋白具有相似的跨膜结构区[26],这种膜包埋的结构可赋予secY 蛋白“转运蛋白”功能,为膜蛋白的分泌提供通道,但secY 蛋白中不存在信号肽。由于植原体缺乏细胞壁,其细胞膜可直接与寄主植物的细胞内环境进行接触,一些与致病性相关的致病因子可能通过Sec 转运系统直接释放到宿主细胞中,导致植株韧皮部功能受损,从而诱发寄主产生抗病性,影响植物的防御响应,进而直接或间接地导致寄主植物或介体昆虫发生互作,诱使寄主植物发生典型植原体病害症状。但secY 蛋白在植原体内转运机制中扮演的具体角色尚不明确,还有待进一步研究和证实,本研究将为了解Sec 转运系统在植原体的分子致病机制及防治植原体病害上提供更大的帮助。
4 结论
本研究是首次对小驳骨丛枝植原体病害的相关报道,明确了小驳骨丛枝病是由植原体所侵染发生的,确定了该植原体株系16S rII-A 亚组的分类地位;同时对secY 蛋白在Sec 分泌蛋白转运系统中的作用进行了初步探讨,为了解Sec 转运系统在植原体的分子致病机理及防治此类病害提供帮助
