微电极离子流技术在植物逆境生理研究中的应用及展望
2021-03-15赵怡琳吕铭汪晓丽柏彦超单玉华
赵怡琳 吕铭 汪晓丽 柏彦超 单玉华

摘要:植物在遭遇盐碱、干旱、重金属、低温、酸、机械刺激等非生物胁迫时,细胞膜电化学特性的变化和调节往往是最早发生的植物细胞反应之一,并与细胞内的生理代谢活动之间存在复杂的联系。基于此,在过去的30年间,微电极离子流技术以其非损伤性、实时性、灵敏性和高分辨率等特有的技术优势,成为了研究逆境胁迫条件下植物的生理响应及调节机制常用的技术手段。从跟踪监测界面反应、解析基因功能、进行抗逆育种和研究信号物质等方面综述该技术在植物逆境胁迫生理研究中的应用,旨在为研究植物功能基因组学和调节植物对环境的适应性提供参考。
关键词:微电极离子流技术;逆境胁迫;离子流;活体;基因功能
中图分类号:Q945.78;Q-3文獻标志码:A
文章编号:1002-1302(2021)01-0043-06
作者简介:赵怡琳(1998—),女,江苏常州人,硕士研究生,主要从事植物逆境生理研究。E-mail:1324822006@qq.com。
通信作者:汪晓丽,博士,副教授,主要从事植物营养电生理研究。E-mail:xlwang@yzu.edu.cn。
非损伤微测技术(non-invasivemicro-testtechnique,NMT),别称微电极离子流检测(microelectrodeionfluxestimation,MIFE),最早由神经科学家Jaffe于1974年提出原初概念[1],如今开始广泛应用于动植物科学、微生物学、神经科学、医学生理学、分子遗传学等生物学领域,是近年来发展起来的可进行生理特征、生理功能动态测量的研究新手段[2]。在植物抗逆方面,NMT以其特有的优势被众多研究者用于干旱、高温、低温、盐渍、重金属、病原菌等非生物胁迫和生物胁迫的研究中[2]。研究受逆境胁迫后植物组织或细胞表面附近的离子流变化,对于揭示植物在遭遇逆境时的物质跨膜转运特征、生理响应机制和调节机制都具有重要意义。
1非损伤微测技术的原理和特点
当细胞或组织不论以主动还是被动方式吸收离子和分子时,样品表面的离子或分子浓度会发生变化,与环境浓度形成浓度差,从而产生电化学势梯度。非损伤微测技术通过离子/分子选择性微电极技术,玻璃电极在临近样品表面的一段微小的距离内来回移动,同时测量电化学势梯度,通过Nernst方程得到该距离内的离子或分子浓度差,并通过Fick第一扩散定律计算出流速,流速的正负符号反映了离子流或分子流移动的方向[3]。
NMT具有活体、实时、动态、直观的优点,并可以长时间多维度地进行离子/分子流测量与扫描[3]。该技术可用于各种生物组织或细胞的研究,可单独或同时测量H+、Ca2+、K+、NH+4、Cd2+、NO-3、O2、H2O2等离子/分子,为生物学研究提供了良好的试验系统平台[4]。相比于传统的微电极技术,其优势在于高灵敏度,两者相差约6个数量级,而且在研究植物生理生态学过程中不须要破坏组织或细胞,能直接进行活体测定,可以实时反映进出植物体组织或细胞的离子/分子的动态变化规律。
2非损伤技术在植物抗逆中的应用
2.1界面反应
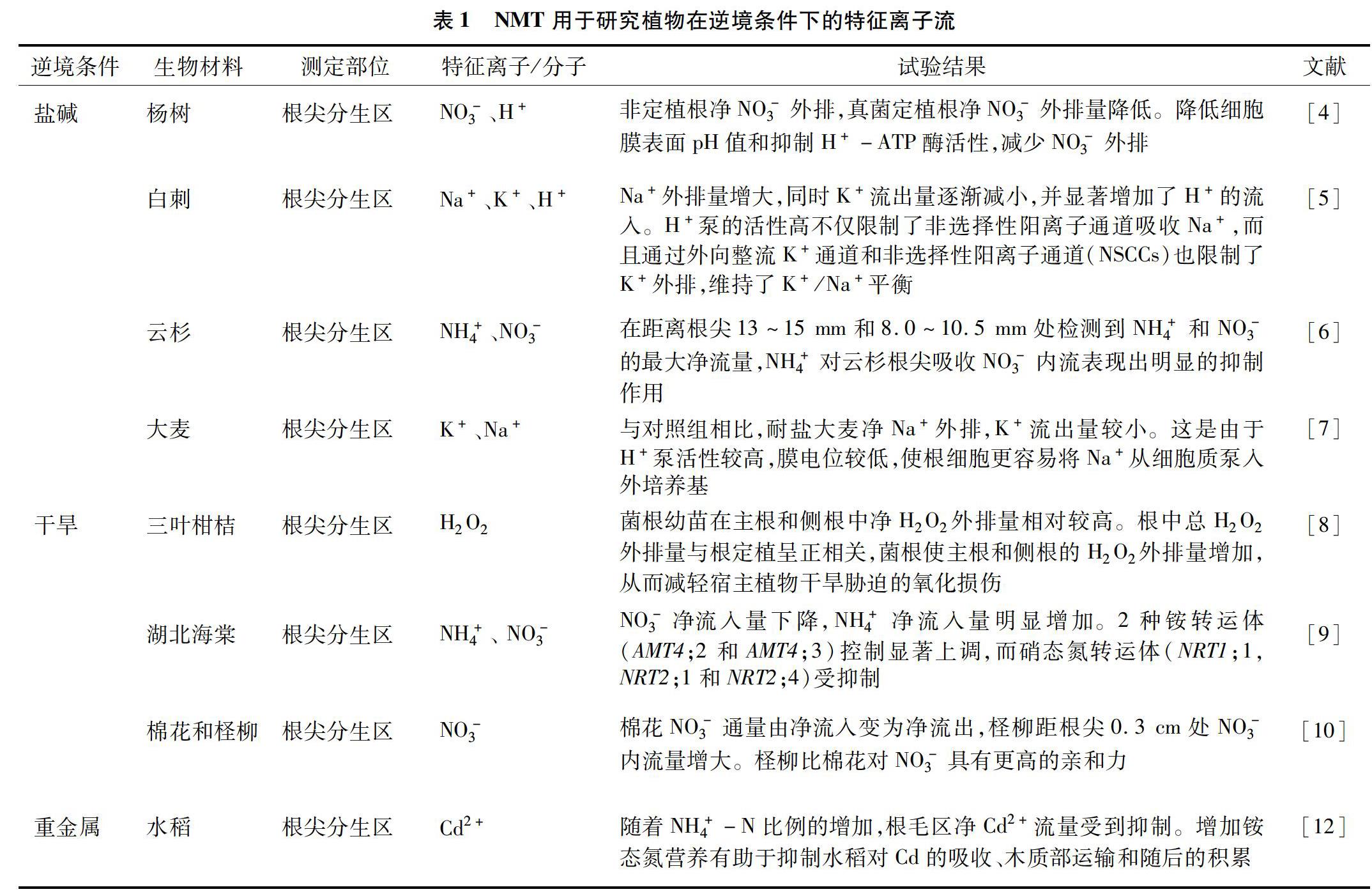
植物在遭遇逆境时会有一系列的生理生化反应,从而表现出各种特有的响应和调节机制。细胞膜透性和功能的改变是植物对逆境条件最为直接和快速的响应[4],进而才是在细胞器或细胞质中进行的同化、代谢等生理过程。因此,从逆境条件下植物与外界环境之间的界面反应着眼,探索界面上特征离子或关键离子的流动速度和方向变化,对研究植物的逆境生理具有极为重要的意义。研究者们已对盐渍、干旱、重金属的胁迫方面进行了广泛的研究,所采用的生物材料、特征离子和测定部位等列于表1。
Sa等研究了杨树在盐胁迫下根表NO-3和H+的净通量,非定植根表现出较强的净NO-3外排,而真菌定植通过降低细胞膜表面pH值和抑制H+-ATP酶活性,阻止了NO-3的损失[5]。Tang等采用NMT研究了不同NaCl浓度下小果白刺(NitrariasibiricaPall.)根幼苗分生组织中Na+、K+和H+的流速,NaCl处理后,根系稳定的K+流出量逐渐减小,并显著增加了H+的流入[6]。Tang等研究了云杉根尖NH+4和NO-3的流速,在距离根尖13~15mm和8.0~10.5mm处检测到NH+4和NO-3的最大净流量,NH+4对云杉根尖吸收NO-3内流表现出明显的抑制作用[7]。Chen等采用NMT测定大麦根部K+和Na+的流速,发现具有耐盐性的大麦H+泵活性较高,膜电位较低,使根细胞更容易将Na+从细胞质泵入外培养基[8]。Huang等对三叶柑桔根系H2O2流速进行研究,发现菌根使主根和侧根的H2O2外排量增加,从而减轻宿主植物干旱胁迫的氧化损伤[9]。Huang等使用NMT研究发现,湖北海棠(MalushupehensisRehd.)细根表面NO-3净流入量下降,NH+4净流入量明显增加,说明NH+4在提高海棠耐旱性方面可能发挥着更重要的作用。在干旱胁迫下,2种铵转运体(AMT4;2和AMT4;3)均显著上调,而大多数与硝酸盐吸收、还原和氮代谢相关的基因均下调。在正常氮水平下,5%聚乙二醇处理植株的生物量产量、根系生长和氮素吸收/减少值均高于低氮处理植株。这些结果表明,干旱胁迫对湖北海棠的不利影响可能随着氮素的增加而减轻[10]。Zhang等的研究结果表明,利用聚乙二醇(PEG)模拟的干旱条件下,棉花NO-3通量由净流入变为净流出,在距根尖0.3cm处有1个明显的峰值[11]。对照条件下,柽柳(Tamarixramosissima)距根尖0.3cm处未观察到明显的NO-3流入信号,而PEG处理显著增强了柽柳距根尖0.3cm处NO-3的流入。Lv等首次制备了一种同时测定3种离子的氮化碳超灵敏微电极,用于测定水稻根表面不同位点Cu2+、Pb2+、Hg2+的流速[12]。Wu等研究了不同NO-3/NH+4比值对水稻根系吸收镉(Cd)、木质部转位以及随后Cd积累的生理和遗传机制,结果表明,随着NH+4-N比例的增加,根毛区净Cd2+流量受到抑制[13]。此外,2种剂量Cd处理下,木质部汁液中Cd浓度也呈现出随NH+4-N比值增加而下降的趋势。增加铵态氮营养有助于抑制水稻对Cd的吸收、木质部运输和随后的积累。
2.2解析基因功能
随着转基因技术的发展,从植物的分子机制研究其抗逆机制也更为普遍。从分子学角度,植物抵御逆境是基因调控的结果。植物遭遇逆境时,通过启动或关闭某些相关基因,改变生理代谢活动使植物适应逆境[13]。通过基因工程,将可能具有抗性的基因组导入拟南芥等模式植物中,研究基因组对植物抗逆的作用和机制。基因组学方法可以全面研究抗逆基因的功能和表达调控,并且把模式植物的抗逆信息推广到基因组复杂的农作物上去[14]。同时可以克隆抗逆基因,在没有抗性的植物中通过基因工程手段导入,改良作物的抗逆性[15]。
NMT在研究植物抗逆基因中的应用见表2。Fan等将海马齿(Sesuviumportulacastrum)的SpAHA1基因轉录入拟南芥中,使用NMT测定质膜中Na+、H+和K+的流速,结果显示,SpAHA1蛋白在胞质膜上具有H+-ATPase的功能,SpAHA1蛋白通过增加质子的电化学梯度来降低Na+的积累,促进质膜上Na+/H+反向运输体的交换活性,在植物抵抗盐碱逆境中起着十分重要的作用[16]。拟南芥CYSTM3在耐盐胁迫方面起着负调控作用是Xu等运用NMT发现的,CYSTM3基因可以抑制拟南芥根系Na+外排,抑制一系列活性氧清除酶的活性[17]。Zhang等同样研究了转基因拟南芥,来自胡杨的PeJRL基因在NaCl胁迫下转基因植株保留了K+,限制了Na+的积累[18]。PeJRL转基因株系增加了Na+的外排量,这与编码细胞膜SOS1、AHA1和AHA2基因的上调有关。PeJRL过表达植物中活化的H+-ATPases抑制了由NaCl诱导的去极化激活的K+通道介导的损失。此外,Cd2+借用Cu2+通道、Cu2+激活Ca2+通道等,均是Cd2+吸收增加的潜在机制。Zhang等的研究表明,MhMAPK4基因过表达降低了烟草根中Cd2+的净流入量,转基因烟草中Cd2+流入到稳定状态的恢复时间也比野生型烟草短,MhMAPK4过表达降低了烟草根细胞的死亡和凋亡,MhMAPK4通过调节根对Cd2+的吸收来调节Cd的积累,通过调节液泡加工酶(VPE)的活性来控制Cd引起的细胞死亡[19]。Ma等鉴定了粳稻抗寒性的数量性状位点为COLD。过表达COLD1基因可显著提高水稻的抗寒性,而缺乏或低表达COLD1基因的水稻对寒冷敏感。COLD1编码定位于细胞膜,是内质网的G蛋白信号调节因子。它与G蛋白相互作用,激活Ca2+通道感知低温,加速G蛋白GTPase活性,使水稻具有抗寒性[20]。Zhou等使用NMT分析显示,转基因拟南芥根在冷冲击下Ca2+内流增加。在冷胁迫下,胞质Ca2+作为信号分子的增加激活了下游COR基因的表达。因此,Ca2+的流入可能导致转基因拟南芥COR基因表达增加[21]。
2.3抗逆育种
传统方法筛选具有抗性的植物品种较为耗时费力。非损伤微测技术的活体动态测量,使试验周期缩短,能较为快速地进行选种育种。由于不同品种的植物在逆境下的生理生化反应不同,所表现出的离子流特征也各有不同。将具有耐性的品种与没有耐性的品种作对比试验,了解其抗逆机制,总结具有耐性品种的离子流特征,建立抗性辨识模型。通过测定植物组织或器官具有特征性的一种或多种离子/分子流,来判断该植物是否具有抗性,从而进行抗逆育种。
Liu等对马铃薯二倍体和六倍体根系K+、H+、Ca2+和Na+的净通量进行研究,结果表明在盐胁迫下,在根和叶组织中六倍体比二倍体保留更多的K+,积累更少的Na+。六倍体由于质膜Ca2+通道对H2O2的高敏感性,有效地将Na+区隔在伸长和成熟根区[22]。Chen等为进一步了解大麦耐盐性性状的遗传行为,在6个大麦品种间进行半双列杂交,以已知耐盐性品种为对照,研究在盐胁迫条件下植物根系K+损失为基础的耐盐性配合力。耐盐品种CM72和Numar表现出较高的一般配合力(GCA),有较高的耐盐性(在盐胁迫下K+损失量较小)[23]。毛桂莲等研究灌木叶片净Na+、K+和Ca2+流速,发现在NaHCO3胁迫下,3种灌木通过不同的策略来消除Na+毒害。宁夏枸杞叶片通过将Na+外排来降低钠毒害,而2种滨藜是将Na+区隔化,在叶表皮形成盐腺[24]。Zhang等测定茶树叶片K+流速,模拟干旱胁迫诱导的茶叶细胞中K+外排与12个茶树品种的整体耐旱性呈较强的负相关。与此相一致的是,耐旱品种叶片K+的残留量明显高于敏感品种。外源施用K+显著减轻了茶树的干旱诱导症状,说明K+的保留是茶树耐旱机制的重要组成部分。药理试验结果表明,耐干旱和敏感茶树品种间,K+向外整流通道和非选择性阳离子通道对PEG诱导的叶肉细胞K+外排的控制作用是不同的[25]。
2.4信号物质
植物感受胁迫信号,Ca2+在信号传递过程中发挥不可缺少的作用。Ca2+调节植物对环境胁迫的反应,在胁迫条件下,胞内Ca2+常常显著增加,可以启动相关基因激活一系列生化反应,使植物能够适应环境胁迫[26]。Ca2+还能诱导相关蛋白合成,激活酶活性,参与活性氧的产生,使植物抗性增强[26]。使用非损伤微测技术检测Ca2+流,能较为方便直观地观察到逆境下Ca2+流速和方向的变化,可以进一步研究Ca2+流对其他离子/分子流的影响,探究钙信使在植物抗逆过程中起的作用。
Lang等通过研究甘草(Glycyrrhizauralensis)根部Na+和K+流速发现,盐胁迫会提高在调节K+/Na+平衡中起信号分子作用的Ca2+、H2O2、NO、胞外ATP水平,促进Na+的外流,抑制K+的损失[27]。Chao等研究了铀胁迫下鬼臼菌根(Syngoniumpodophyllum)根部Ca2+流速,结果表明,黑曲霉能抑制根细胞的钙外排,从而减弱了铀对植物生长的毒害作用[28]。在干旱胁迫下,H2S介导离子通量诱导拟南芥气孔关闭是Jin等通过测定拟南芥叶片H+、Ca2+、K+和Cl-流速发现的,此外还发现内源性H2S在不影响H+流量的情况下,可诱导跨膜K+流出以及Ca2+、Cl-流入[29]。Rodrigo-Moreno等研究了铜胁迫下不同拟南芥根部的离子转运情况,发现低浓度的Cu2+刺激根尖Ca2+内流,但未刺激成熟区,铜诱导K+外排并且在铜暴露后,根尖的基础过氧化积累有所减少。铜会在胞质中产生羟自由基,从而调控细胞膜羟自由基敏感性的Ca2+和K+转运系统[30]。
3离子流与植物抗逆的联系
在逆境中植物往往会受到渗透胁迫、离子失调、膜透性改变、生理代谢紊乱等危害,植物对逆境的生理方面的适应变化包括改变生物膜透性、形成逆境蛋白、产生抗氧化防御系统、进行渗透调节等[31],其中渗透调节物质包括细胞与外界进行无机离子交换,如K+、Na+[31];细胞内合成有机物质,如脯氨酸、甜菜碱等[31]。因此在植物抗逆生理过程中进行渗透调节会伴随着H+、Ca2+、K+、Cl-、Na+、H2O2等离子/分子流入或流出植物液泡或细胞。非损伤微测技术的试验结果直接反映的是离子流速,从吸收或外排的离子动态变化角度可以体现植物细胞的渗透调节,表现出植物的抗逆能力。
植物抵抗盐碱胁迫,可以通过细胞质膜上的H+-ATP酶将H+泵到胞外,形成跨膜的H+浓度梯度从而驱动Na+/H+反向运输体将细胞内的Na+排出,同时也抑制了K+外流。Chen等对具有较强耐盐碱胁迫能力的坛紫菜进行了研究,结果表明,与对照海草相比,坛紫菜具有较高的K+/Na+比值。坛紫菜通过Na+/H+反载体将Na+外排,同时降低了海盐藻脱极化激活通道K+的损失,在盐胁迫下维持Na+/H+的稳态,使坛紫菜具有抗高盐胁迫能力[32]。Ca2+在植物干旱胁迫中,是植物细胞内重要的调节生理功能的信号物质。郎涛等研究发现,胞外ATP、H2O2、Ca2+、NO等盐胁迫信号是通过上调红树根系细胞质膜Na+/H+逆向转运体系活性,在促进Na+和H+逆向跨膜转运的同时,抑制K+外流[33]。
植物抵抗干旱胁迫主要通过细胞进行渗透调节来保持吸水能力[34],在根尖细胞的无机渗透调节物质,如K+、Cl-能保持渗透压力和水分梯度,有利于细胞在干旱下维持水分。植物叶片的气孔开闭也与K+密切相关,保卫细胞中K+外排使水势上升,气孔关闭,减少植物蒸腾作用。Mak等研究发现,PEG诱导的短期离子通量响应不同于植物在长期真实干旱条件下的离子通量,长期干旱导致H+流入量更小,K+和Ca2+的流出量更大[35]。聚乙二醇诱导的干旱和实际干旱条件在调节这些化学信号和膜传输系统方面可能是不同的。在PEG短期诱导的大豆基因表达和蛋白质组学研究中,没有发现负责K+、H+和Ca2+通量的转运体发生变化,说明短期胁迫时离子的流入流出是植物的应激反应。
植物对重金属的排斥包括分泌化合物降低有效性、回避摄入、限制运输和排出体外[36],研究者们通过NMT研究植物对重金属的吸收机制。Ma等研究发现,有硅培养的细胞与无硅培养的细胞相比,净Cd2+内流显著减少[37]。因为细胞壁中64%的硅在细胞壁分离后与半纤维素成分结合,从而抑制了镉的吸收。有硅培养的细胞壁表面电位的异质性高于无硅培养的细胞壁表面电位的异质性,且在添加镉后均质化。
4问题与展望
由于全球气候变化加剧,盐碱化、干旱、洪澇、低温、高温等环境问题愈发严重,对植物的生存生长不利。多数植物通过调节自身生理机能对逆境有不同的抵抗机制,在生理调节过程中往往伴随着H+、Ca2+、K+、Cl-、Na+、H2O2等离子/分子流入或流出植物体的过程,通过NMT进行离子/分子流的研究能够从全新角度阐述植物的抗逆机制,获得常规方法难以获得的全新发现[4]。NMT直接在根、叶片细胞等活体植物样品上,在不损伤细胞膜的情况下获得离子/分子流速,不仅能够研究植物抵抗逆境的生理生化反应、微观上基因调控的机制,还能够通过对植物耐性机制的研究,筛选具备优良性状的生物品种。但非损伤微测技术在植物抗性研究方面也存在不足,多数植物都具有抵抗逆境的能力,所能获得的植物材料有很多,检测的离子或分子也多种多样,不同的离子/分子流速在不同的植物材料中有不同的变化趋势,但建立抗性辨识模型的比较少,没有具体的评价指标。目前的研究大多仅是单方面采用NMT在植物某一生长时期进行离子或分子流速的测定,而研究植物的长期演化过程是动态连续的,各要素间存在复杂的互作关系,须要将NMT应用于植物整个生命周期的不同发育阶段的研究中[4]。
非损伤微测技术在理论研究和试验实践中,研究不断深入,探测范围不断扩大,技术不断完善,非损伤微测技术必将更准确、更全面地揭示植物体离子跨膜运动与植物生理功能之间的关系[2]。关于非损伤微测技术本身的改进,可以增加可检测的离子/分子种类,探究植物体生命活动规律中不同离子/分子所起的作用,发现更多植物特定的离子/分子流速的变化趋势,形成特征性图谱。随着可检测的离子/分子种类增多,非损伤微测技术须加快技术革新,每次单一测定一种离子/分子已经不能满足要求,须要发展多通道离子检测技术,更便捷地同时测定多种离子/分子,有效发挥实时、活体测定离子/分子的优势。对于该技术的应用技巧方面,完整活体植株固定在操作平台上测定时根系易晃动,使用滤纸和小瓷片压住样品根系;植物材料相同品种间根系活力差异较大,在准备样品时选择较为强盛的根系,并多做平行试验;仪器反应太灵敏,容易产生基线漂移,须在测定同种离子流不同品种时进行多次校准;试验环境要求较高,由于样品容易晃动,数据容易受环境影响而产生波动。还须进一步改进试验条件。非损伤微测技术也应与其他先进检测技术结合研究,如荧光显微、分子生物学、微域分析等,从不同层次不同角度全方位揭示植物的生理生化过程,逐步完善植物长期演化过程的应用研究体系。拓展非损伤微测技术的应用领域需要丰富样品的品种、类型以及不同的测试部位,如微管形成层、木质部等。同时新的逆境条件出现在不断出现,如纳米物质、病虫害等,了解植物在新逆境条件下的适应性与植物的生理生化机制,从而推广非损伤微测技术在植物抗逆以及各领域研究中的应用。
参考文献:
[1]JaffeLF,NuccitelliR.Anultrasensiticevibratingprobeformeasuringsteadyextracellularcurrents[J].JournalofCellBiology,1974,63(2):614-628.
[2]刘科,张丙林,张文英,等.非损伤离子流检测技术在作物逆境研究中的应用[J].应用生态学报,2018,29(2):678-686.
[3]李静,韩庆庆,段丽婕,等.非损伤微测技术在植物生理学研究中的应用及进展[J].植物生理学报,2014,50(10):1445-1452.
[4]贾代东,刘爱琴,李惠通,等.非损伤微测技术在植物生理生态学研究中的应用进展[J].应用与环境生物学报,2017,23(1):175-182.
[5]SaG,YaoJ,DengC,etal.AmeliorationofnitrateuptakeundersaltstressbyectomycorrhizawithandwithoutaHartignet[J].NewPhytol,2019,222(4):1951-1964.
[6]TangX,YangX,LiH,etal.MaintenanceofK+/Na+balanceintherootsofNitrariasibiricaPall.inresponsetoNaClstress[J].Forests,2018,9(10):601.
[7]TangB,YinCY,LiuQ.CharacteristicsofammoniumandnitratefluxesalongtherootsofPiceaasperata[J].JournalofPlantNutrition,2019,42(7):772-782.
[8]ChenZH,PottosinII,CuinTA,etal.RootplasmamembranetransporterscontrollingK+/Na+homeostasisinsalt-stressedbarley[J].PlantPhysiology,2007,145(4):1714-1725.
[9]HuangYM,ZouYN,WuQS.AlleviationofdroughtstressbymycorrhizasisrelatedtoincreasedrootH2O2effluxintrifoliateorange[J].ScientificReports,2017,7:42335.
[10]HuangLL,LiMJ,ZhouK,etal.UptakeandmetabolismofammoniumandnitrateinresponsetodroughtstressinMalusprunifolia[J].PlantPhysiologyandBiochemistry,2018,127:185-193.
[11]ZhangL,LiGJ,DongGQ,etal.CharacterizationandcomparisonofnitratefluxesinTamarixramosissimaandcottonrootsundersimulateddroughtconditions[J].TreePhysiology,2018,39(4):628-640.
[12]LvH,TengZ,WangS,etal.VoltammetricsimultaneousionfluxmeasurementsplatformforCu2+,Pb2+andHg2+nearricerootsurface:Utilizingcarbonnitrideheterojunctionfilmmodifiedcarbonfibermicroelectrode[J].SensorsandActuatorsB-Chemical,2017,256:98-106.
[13]WuZC,ZhangWJ,XuSJ,etal.Increasingammoniumnutritionasastrategyforinhibitionofcadmiumuptakeandxylemtransportinrice(OryzasativaL.)exposedtocadmiumstress[J].EnvironmentalandExperimentalBotany,2018,155:734-741.
[14]高銀.植物抗逆机制与基因工程研究进展[J].内蒙古农业科技,2007(5):75-78.
[15]杨柳,张振乾,宋继金,等.植物抗逆基因研究进展[J].作物研究,2010,24(2):126-129.
[16]FanYF,WanSM,JiangYS,etal.Over-expressionofaplasmamembraneH+-ATPaseSpAHA1conferredsalttolerancetotransgenicArabidopsis[J].Protoplasma,2018,255(6):1827-1837.
[17]XuY,YuZP,ZhangSZ,etal.CYSTM3negativelyregulatessaltstresstoleranceinArabidopsis[J].PlantMolecularBiology,2019,99(4/5):395-406.
[18]ZhangHL,DengC,YaoJ,etal.PopuluseuphraticaJRLmediatesABAresponse,ionicandROShomeostasisinArabidopsisundersaltstress[J].InternationalJournalofMolecularSciences,2019,20(4):815.
[19]ZhangWW,SongJF,YueS,etal.MhMAPK4fromMalushupehensisRehd.decreasescelldeathintobaccorootsbycontrollingCd2+uptake[J].EcotoxicologyandEnvironmentalSafety,2019,168:230-240.
[20]MaY,DaiXY,XuYY,etal.COLD1conferschillingtoleranceinrice[J].Cell,2015,160(6):1209-1221.[HJ2mm]
[21][JP3]ZhouAM,LiuEH,LiH,etal.PsCor413pm2,aplasmamembrane-localized,cold-regulatedproteinfromPhloxsubulata,conferslowtemperaturetoleranceinArabidopsis[J].InternationalJournalofMolecularSciences,2018,19(9):2579.
[22]LiuY,YuYC,SunJY,etal.Root-zone-specificsensitivityofK+-andCa2+-permeablechannelstoH2O2determinesionhomeostasisinsalinizeddiploidandhexaploidIpomoeatrifida[J].JournalofExperimentalBotany,2019,70(4):1389-1405.
[23]ChenZH,ShabalaS,MendhamNJ,etal.CombiningabilityofsalinitytoleranceonthebasisofNaCl-inducedK+fluxfromrootsofbarley[J].CropScience,2008,48(4):1382-1388.
[24]毛桂莲,李国旗,许兴,等.NaHCO3胁迫下3种灌木Na+、K+、Ca2+的吸收及转运[J].应用生态学报,2014,25(3):718-724.
[25]ZhangXC,WuHC,ChenLM,etal.Mesophyllcellsabilitytomaintainpotassiumiscorrelatedwithdroughttoleranceintea(Camelliasinensis)[J].PlantPhysiologyandBiochemistry,2019,136:196-203.
[26]耶興元.Ca2+与植物抗逆性研究概况[J].信阳农林学院学报,2008,18(1):124-126.
[27]LangT,DengSR,ZhaoN,etal.Salt-SensitivesignalingnetworksinthemediationofK+/Na+homeostasisgeneexpressioninGlycyrrhizauralensisroots[J].FrontiersinPlantScience,2017,8:1403.
[28]ChaoZ,ShaYH,DingDX,etal.Aspergillusnigerchangesthechemicalformofuraniumtodecreaseitsbiotoxicity,restrictsitsmovementinplantandincreasethegrowthofSyngoniumpodophyllum[J].Chemosphere,2019,224:316-323.
[29]JinZP,WangZQ,MaQX,etal.HydrogensulfidemediatesionfluxesinducingstomatalclosureinresponsetodroughtstressinArabidopsisthaliana[J].PlantandSoil,2017,419(1/2):141-152.
[30]Rodrigo-MorenoA,Andres-ColasN,PoschenriederC,etal.Calcium-andpotassium-permeableplasmamembranetransportersareactivatedbycopperinArabidopsisroottips:linkingcoppertransportwithcytosolichydroxylradicalproduction[J].Plant,CellandEnvironment,2013,36(4):844-855.
[31]黄绢.转JERF36基因银中杨的抗旱性评价及生理机理研究[D].北京:中国林业科学研究院,2016.
[32]ChenTX,WangWL,XuK,etal.K+andNa+transportcontributetoK+/Na+homeostasisinPyropiahaitanensisunderhypersalinestress[J].AlgalResearch,2019,40:101526.
[33]郎涛.盐胁迫下泌盐与非泌盐红树离子平衡调控信号网络研究[D].北京:北京林业大学,2014.
[34]张海娜,鲁向晖,王瑞峰,等.稀土尾砂干旱胁迫对2种牧草种子萌发与幼苗生理特性的影响[J].江苏农业科学,2019,47(13):204-208.
[35]MakM,BablaM,XuSC,etal.LeafmesophyllK+,H+andCa2+fluxesareinvolvedindrought-induceddecreaseinphotosynthesisandstomatalclosureinsoybean[J].EnvironmentalandExperimentalBotany,2014,98:1-12.
[36]王学东,周红菊,华珞.植物对重金属的抗性机理及其植物修复研究进展[J].南水北调与水利科技,2006,4(2):43-46.
[37]MaJ,CaiHM,HeCW,etal.Ahemicellulose-boundformofsiliconinhibitscadmiumionuptakeinrice(Oryzasativa)cells[J].NewPhytologist,2015,206(3):1063-1074.
