黄瓜发育早期耐旱突变体筛选与鉴定
2021-03-15陈皓炜刘志宇安宇宁王茹宋蒙飞陈洁娄群峰
陈皓炜 刘志宇 安宇宁 王茹 宋蒙飞 陈洁 娄群峰
摘要:以863份黄瓜突变体和长春密刺野生型为材料,进行PEG-6000溶液(200g/kg)模拟干旱发芽试验,通过比较发芽势、相对发芽率并综合胚根生长情况,初步筛选得到耐旱材料,再对筛选出的材料和野生型材料长春密刺进行苗期干旱(PEG-6000,200g/kg)处理,并测定形态指标和生理指标,综合比较筛选得到耐旱新材料。结果表明,干旱发芽试验下,初步得到11份耐旱突变体材料。幼苗期干旱胁迫处理下11份突变体材料与长春密刺野生型各项形态指标和生理指标都呈现不同程度的变化,株高、茎粗和地下鲜质量均降低,而超氧化物歧化酶(SOD)活性、丙二醛(MDA)含量、脯氨酸(Pro)含量及可溶性蛋白质含量上升。隶属函数法比较筛选得到7份高耐旱新材料。
关键词:黄瓜;发育早期;突变体;耐旱;筛选;鉴定
中图分类号:S642.201文献标志码:A
文章编号:1002-1302(2021)01-0112-06
作者简介:陈皓炜(1999—),男,浙江义乌人,主要从事葫芦科作物遗传育种与种质创新研究。E-mail:2629015119@qq.com。
通信作者:娄群峰,博士,教授,主要从事葫芦科作物遗传育种与种质创新研究。E-mail:qflou@njau.edu.cn。
干旱是世界性的主要自然灾害之一,占所有非生物危害的首位,是限制植物生长发育和产量的重要环境因素[1]。干旱半干旱区约占全球陆地总面积的30%,我国干旱半干旱区约占国土面积的40%[2],而根据模型预测,随着气候变化的加剧,干旱区的面积还将继续扩大[3]。此外,季节性干旱还时常影响我国非干旱的主要农业地区,如何提高农作物的耐旱性一直是研究的热点[4]。
黄瓜(CucumissativusL.)是世界性的重要蔬菜作物,我国黄瓜种植面积约126.67万hm2,占世界黄瓜栽培面积的60%,年产量达6487万t,规模和年产量都位居世界第一[5-6]。黄瓜根系较浅,且叶片大,耗水量大,生长发育对于水分的要求高,干旱极易影响黄瓜的生长发育、果实品质和产量[7]。
黄瓜优质材料筛选已在耐盐、低温、高温等耐逆方面得到实现,而耐旱方面研究仍集中于黄瓜生理指标对干旱的响应[8-13],对耐旱黄瓜种质选育的相关研究较少,现有黄瓜耐旱种质资源缺乏。同时,黄瓜遗传背景狭窄,现有可用来表型筛选的种植资源缺乏,而EMS(甲基磺酸乙酯)诱变突变体库为筛选目标材料提供了丰富的资源,可作为本研究的筛选源[14-15]。但用常规方法对植株苗期形态和生理指标鉴定来筛选耐旱材料在本研究需要花费大量的人力和时间,因此简便有效的筛选方法显得十分必要[16]。种子萌发期筛选操作简单且周期较短,可快速过滤大量耐旱性较差的材料,缩小筛选范围,苗期则对粗筛出的材料进一步筛选,得到较可靠的优质耐旱材料。
研究表明,干旱胁迫会抑制种子对水分的吸收,从而使细胞丧失膨压,种子生活力下降,影响种子中与碳氮代谢相关酶的活性及其代谢产物,进而显著抑制种子的萌发[17]。由于缺乏水分,植株在营养生长过程中表现为地下鲜质量变小、株高和茎粗下降,不利于后期的生殖生长[18]。此外,干旱导致外界环境拥有较大的渗透压,植物细胞失水,导致可溶性蛋白和脯氨酸含量上升[19];且干旱会导致细胞膜氧化,引起超氧化物歧化酶(SOD)活性和丙二醛含量上升[20]。由于植株耐旱性是多项生理指标共同作用的结果,以某一指标的结果判断材料耐旱性过于片面,且无法对所有参试材料的耐旱性进行合理的定量比较,隶属函数正好可将互相独立的数值转换成综合指标,减小单个指标存在的片面性和误差,增强评价的说服力[21]。
本研究利用聚乙二醇6000(PEG-6000)模拟干旱对黄瓜甲基磺酸乙酯(EMS)突变体库材料进行萌发试验筛选耐旱突变体材料,再分析盐胁迫下耐旱突变体材料的形态和生理指标特征,利用平均隶属函数值综合评价不同材料耐旱性,筛选出高耐优良材料,并阐明黄瓜耐旱生理机制,为黄瓜耐旱的品种选育提供优良材料,同时为耐旱生理机制的进一步研究提供理论基础。
1材料与方法
1.1试验材料与处理
本试验使用的材料由南京农业大学葫芦科作物遗传与种质创新实验室提供。前期已获得用EMS诱变处理的华北型黄瓜材料长春密刺,自交3代获得突变体库。试验于2019年5—11月于南京农业大学作物遗传与种质创新国家重点实验室进行。
取上述的863份突变体材料进行编号,每个编号取5粒饱满整齐的种子,用清水冲洗后放入装有55℃清水的纸杯中浸泡4h,将种子取出吸去多余的水,放入垫有2层滤纸的培养皿中,加入20mL的200g/kgPEG-6000溶液,置于光培箱中黑暗发芽,温度为(28±1)℃,重复3次。以胚根突破种皮2mm为准,分别于2d后统计种子发芽数,4d后统计种子发芽数、胚根长度,最后计算出各个编號种子的发芽率、发芽势、相对发芽率。根据种子的各项发芽指标,对比筛选出耐旱型、中间型、旱敏型3种突变体材料。
将发芽期初步筛选出来的耐旱材料和长春密刺野生型进行浸种催芽,待种子出芽后,播入挖有小孔的泡沫板,放入装有清水的塑料盘中进行水培,每天更换清水。在幼苗的子叶展开后,换用1/2Hogland营养液水培;待幼苗长至3叶1心后,每份材料挑选12株长势整齐一致的黄瓜苗平均分为2组:处理组和对照组;处理组营养液中加入一定量的PEG-6000使之浓度达到100g/kg,预处理1d,再加PEG-6000使其浓度达到200g/kg,处理2d后测定植株形态指标和叶片生理指标,每个指标设3个重复。
1.2测定指标与方法
1.2.1发芽指标发芽势=2d后种子发芽数/供试种子数×100%;发芽率=4d后种子发芽数/供试种子数×100%;相对发芽率=供试种子发芽率-对照长春密刺种子发芽率。
1.2.2形态指标用游标卡尺于子叶部位向下0.5cm处测量茎粗(mm);用直尺测量从泡沫板表面到幼苗最高生长点部位的距离(cm);将幼苗在泡沫板以下的地下部与植株分离,用吸水纸吸干表面残留的营养液,电子天平称地下部鲜质量(g)。
1.2.3生理指标超氧化物歧化酶(SOD)采用氮蓝四唑(NBT)还原法[22]测定;丙二醛(MDA)含量采用硫代巴比妥酸法[23]测定;脯氨酸(Pro)测定采用酸性茚三酮法[24]进行;可溶性蛋白质含量用考马斯亮蓝法[25]测定。
1.2.4隶属函数值模糊集合中的隶属函数,可将互相独立的数值转换成1个综合的指标,现在综合评判耐逆性常用此方法。
耐旱性隶属函数值算法如下:
若测得指标与耐旱性正相关,则U=(X-Xmin)/(Xmax-Xmin);
若测得指标与耐旱性负相关,则U=1-(X-Xmin)/(Xmax-Xmin)。
式中:X表示某指标的一个测定值;Xmin为该指标测定值中的最小值;Xmax为该指标测定值中的最大值;U表示指标该测定值的隶属函数值。计算各指标隶属函数值的均值,均值越大,则表明耐旱性越强。
1.3数据处理
数据处理分析采用Excel2016和SPSS25.0软件,显著水平α=0.05。
2结果与分析
2.1发芽期黄瓜耐旱突变体材料的筛选
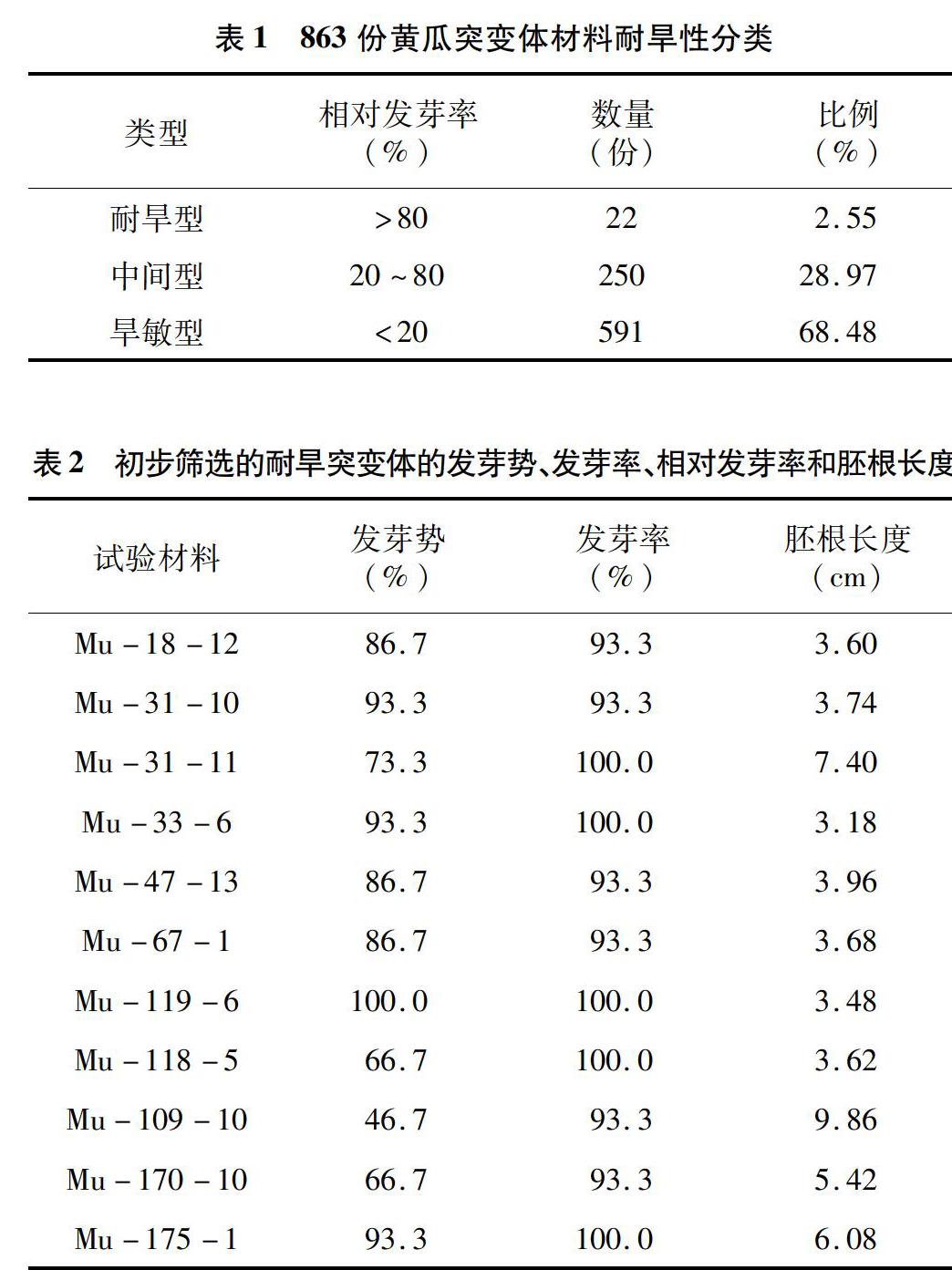
对863份黄瓜突变体材料进行发芽指标统计,数据显示,不同编号种子之间发芽指标差异较为显著。绝大部分突变体材料发芽指标弱于或近似长春密刺野生型(发芽率17.7%),发芽率低甚至为0、胚根很短甚至不生长,表明这些材料耐旱性较差;少部分材料表现出发芽指标显著优于野生型,大部分发芽甚至全发芽、胚根长度较长。根据相对发芽率将所有材料分为耐旱型、中间型和旱敏型3种材料(表1),其中旱敏型相对发芽率小于20%,数量最多,有591份,占总材料数的68.48%;中间型相对发芽率为20%~80%,有250份,占总材料数的28.97%;耐旱型相对发芽率大于80%,仅有22份,占总材料数的2.55%。
根据这22份材料种子的发芽指标,以高发芽势、发芽率和较长胚根长度,从中选出11份表现优异的材料进行下一步鉴定(表2)。
2.2幼苗期黄瓜耐旱材料的鉴定
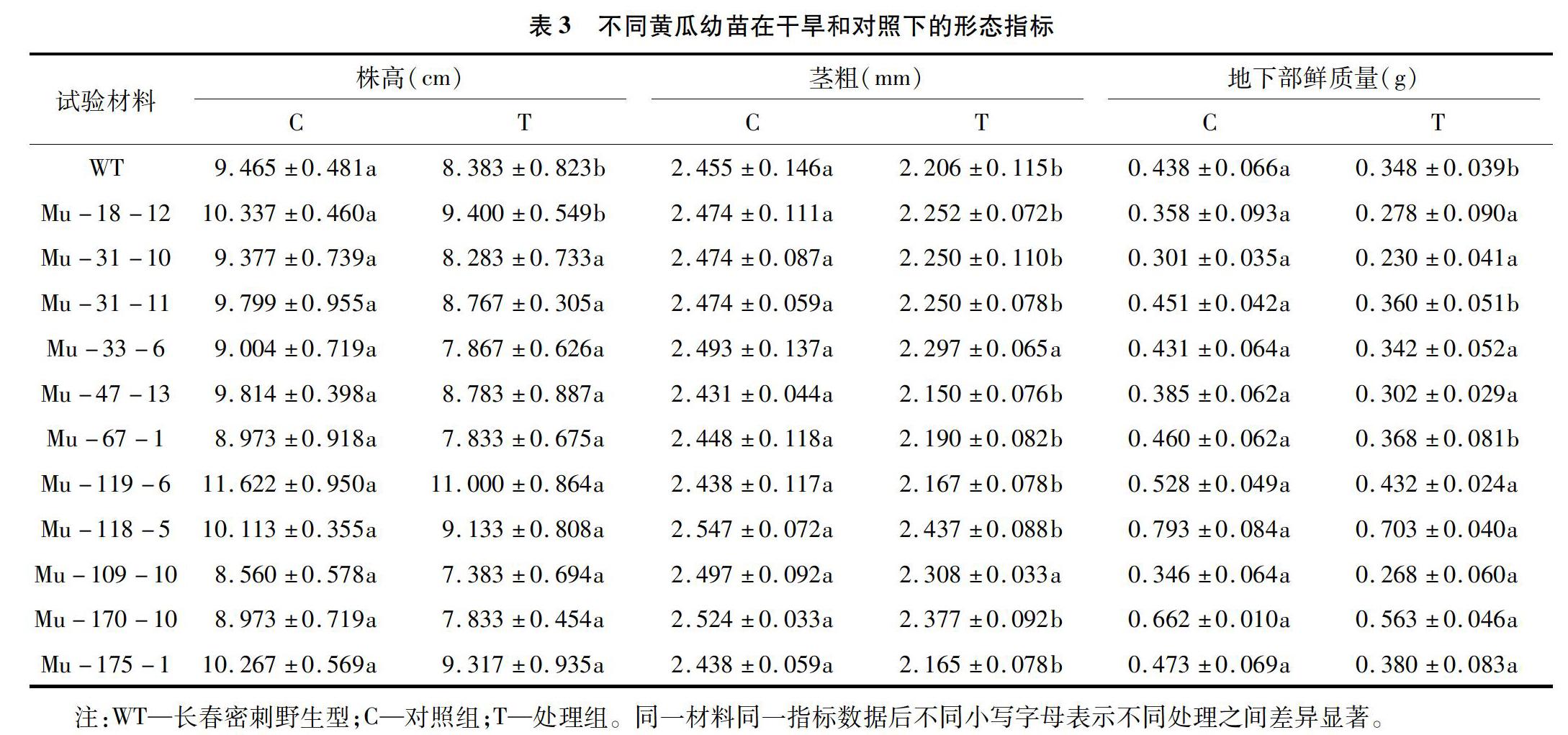
2.2.1形态指标鉴定干旱胁迫下11份黄瓜突变体材料和长春密刺野生型在各个形态指标上均呈现不同程度的下降(表3)。除Mu-18-12和野生型外,其余材料的株高在干旱处理和对照间均无显著差异,且共6份材料(Mu-119-6、Mu-18-12、Mu-175-1、Mu-118-5、Mu-47-13、Mu-31-11)株高降低的百分比(5.35%、9.06%、9.25%、9.69%、10.51%、10.53%)小于野生型(11.43%);仅Mu-33-6和Mu-109-10这2份材料的茎粗在干旱和对照间无显著差异,且7份材料(Mu-118-5、Mu-170-10、Mu-109-10、Mu-33-6、Mu-18-12、Mu-31-10、Mu-31-11)茎粗降低的百分比(4.32%、5.82%、7.57%、7.86%、8.97%、9.05%、9.05%)小于野生型(10.14%);在干旱处理和对照间,除Mu-31-11、Mu-67-1和野生型外,其余材料的地下鲜质量无显著差异,且6份材料(Mu-118-5、Mu-170-10、Mu-119-6、Mu-175-1、Mu-67-1、Mu-31-11)地下鲜质量降低的百分比(11.35%、14.95%、18.18%、19.66%、20.00%、20.18%)小于野生型(20.47%)。
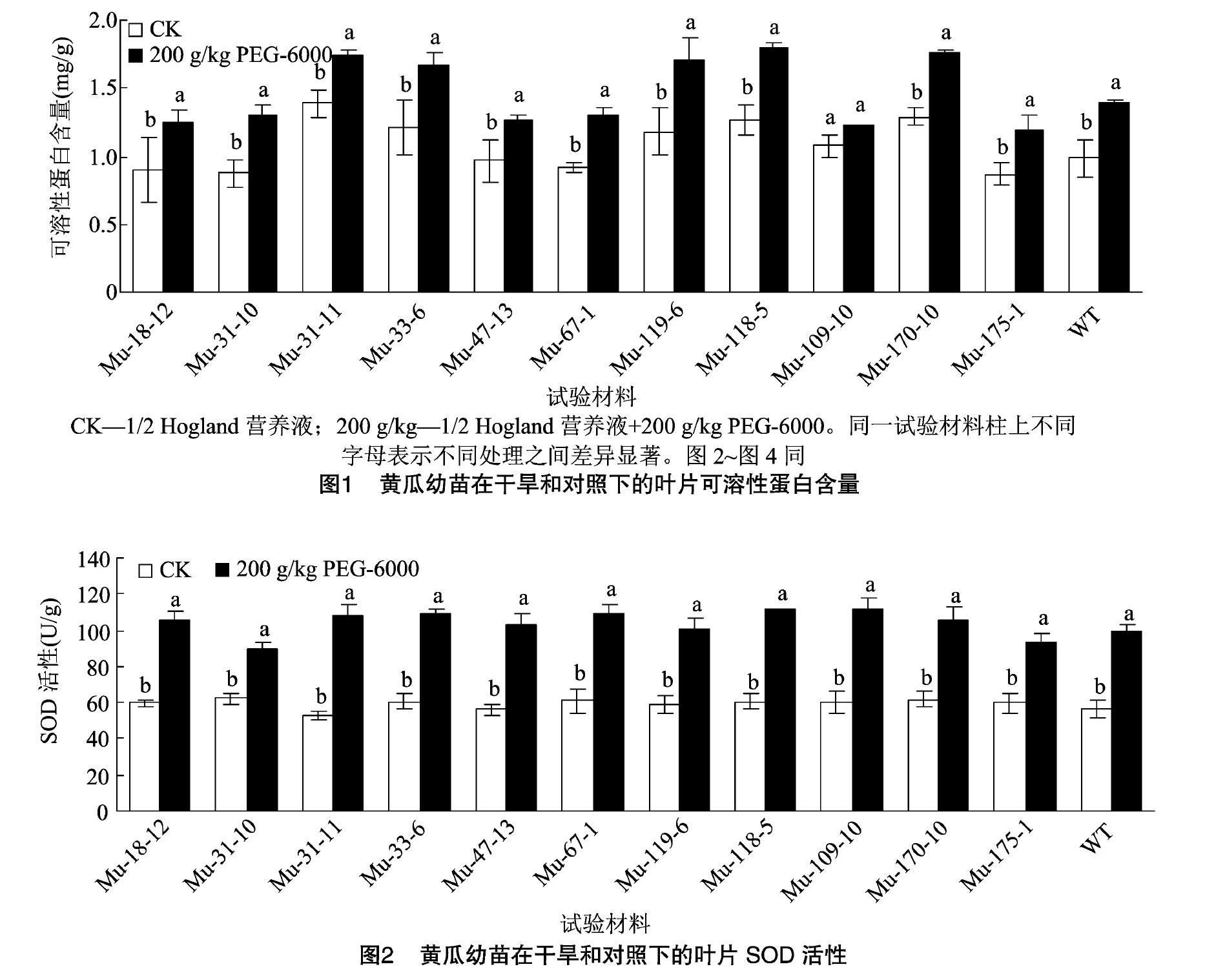
2.2.2可溶性蛋白含量鉴定干旱胁迫使12份材料的可溶性蛋白含量均呈现不同程度的上升趋势(图1),除Mu-109-10外,其余材料在干旱和对照之间的可溶性蛋白含量均存在显著差异。其中,4份材料(Mu-31-10、Mu-119-6、Mu-67-1、Mu-118-5)的可溶性蛋白上升的百分比(48.55%、44.33%、43.35%、42.86%)大于野生型(41.42%)。
2.2.3SOD活性鉴定干旱胁迫能显著提高黄瓜幼苗的超氧化物歧化酶(SOD)活性(图2),每份材料的SOD活性均与对照有显著差异。其中,6份突变体(Mu-31-11、Mu-109-1、Mu-47-13、Mu-118-5、Mu-33-6、Mu-67-1)的SOD活性上升百分比(104.07%、86.58%、83.79%、84.14%、82.51%、79.87%)大于野生型(77.43%)。
2.2.4脯氨酸含量鉴定干旱导致不同材料的脯氨酸含量上升(圖3),除Mu-119-6以外,每份材料的脯氨酸含量在干旱胁迫和对照间均存在显著差异,其中2份材料(Mu-118-5、Mu-31-10)的脯氨酸含量上升百分比(460.00%、412.50%)大于野生型(402.17%)。
2.2.5丙二醛含量鉴定干旱使不同材料的丙二醛含量呈现不同程度的上升(图4),除Mu-119-6、Mu-47-13和Mu-170-10以外,其他材料的丙二醛含量在干旱胁迫和对照间均存在显著差异。突变体中有9份材料(Mu-170-10、Mu-47-13、Mu-67-1、Mu-119-6、Mu-31-10、Mu-118-5、Mu-175-1、Mu-33-6、Mu-31-11)的丙二醛含量上升百分比(13.60%、13.29%、14.42%、16.04%、25.67%、36.92%、39.39%、45.79%、47.68%)小于野生型(54.87%)。
2.3黄瓜耐旱突变体筛选结果鉴定
部分突变体在不同指标表现差异较大,而单一指标无法评判植株的耐旱性,同时不同突变体在正常生长环境下同一生理指标的结果也不尽相同,若直接取干旱胁迫下的指标进行隶属函数值计算,不可避免地存在起始误差,笔者利用盐胁迫下同一时期黄瓜幼苗不同指标上升/下降百分比的隶属函数值的平均值来综合评价不同材料的耐旱性,最终得到12份突变体和长春密刺野生型材料的耐旱性隶属函数值(表4),以长春密刺野生型的表现为分界,根据平均隶属度最终筛选出7份优质耐旱材料,即Mu-118-5、Mu-119-6、Mu-170-10、Mu-67-1、Mu-31-11、Mu-33-6和Mu-47-13,与上述分析结果基本保持一致。
3讨论
干旱严重影响种子萌发、植株的形态结构和生理生化[26-27]。种子的发芽指标和胚根长度可直接反映其对逆境的耐受性,且操作简单、试验周期短,适合大批量的种子筛选试验。聚乙二醇6000(PEG-6000)是不能透过细胞壁的一类大分子渗透调节物,其对细胞毒性小[13]。本试验选用200g/kg聚乙二醇6000模擬干旱环境进行萌发试验,筛选得到11份相对耐旱材料,仅占供试材料的1.27%,表明黄瓜EMS突变体中仅有极少数突变体拥有较好的耐旱性,而绝大部分材料耐旱性较差。
由形态指标结果可见,12份材料在干旱胁迫后指标都下降,但下降比例较小,且不同材料之间差异也较小,形态指标虽能在一定程度上反映植株受干旱危害的程度,但由于形态指标属于表型,不只受种质本身调控,也受外界各种环境影响,再加上处理前后指标变化幅度不大,所得结果容易存在误差,但大体指向仍有参考价值,此时需要内部生理指标来进一步补充。
可溶性蛋白和脯氨酸是植物细胞中的渗透调节物质,在干旱胁迫下大量积累,降低细胞渗透势,促进细胞在干旱环境下吸收外界的水分,以维持细胞正常的形态结构[28],12份材料的可溶性蛋白呈现10%~50%的增幅,其中增幅为40%~50%的材料耐旱性较好。参试材料脯氨酸含量均呈现成倍增长,甚至有2份材料出现4倍以上的增幅,表明其拥有较强渗透调节能力和耐旱性。除此以外,所有材料在干旱下SOD活性都有40%~100%的显著提高,研究表明,干旱下细胞膜脂过氧化,产生大量的活性氧,而SOD是植物细胞重要的耐氧化酶,在接收到活性氧大量增加的信号后,耐氧化系统中SOD活性增强,清除过量的活性氧,缓解干旱可能造成的膜损伤[29],SOD活性越高,说明对干旱胁迫的调节能力越强。SOD虽然可以缓解膜损伤,但在较强的干旱胁迫下,SOD不足以清除所有增加的活性氧,导致细胞膜过氧化、MDA合成增加,故MDA含量是评价细胞膜受损伤程度的指标[30],MDA含量增幅越小,说明细胞膜受损程度越小,植株耐旱性越强,故突变体中9份MDA含量增幅小于野生型的材料拥有较强的耐旱性。综上所述,可溶性蛋白、脯氨酸含量和超氧化物歧化酶活性与黄瓜幼苗耐旱性呈正相关,而MDA含量与耐旱性呈负相关。
本研究以PEG处理模拟干旱,通过发芽期+苗期的筛选形式对长春密刺黄瓜突变体材料进行耐旱性评价,最终由形态和生理指标的平均隶属度筛选得到7份优质耐旱材料:Mu-118-5、Mu-119-6、Mu-170-10、Mu-67-1、Mu-31-11、Mu-33-6和Mu-47-13,研究结果为后续黄瓜耐旱种质的选育和耐旱机制的深入研究提供理论指导。
参考文献:
[1]CellierF,ConéjéroG,BreitlerJC,etal.Molecularandphysiologicalresponsestowaterdeficitindrought-tolerantanddrought-sensitivelinesofsunflower:accumulationofdehydrintranscriptscorrelateswithtolerance[J].PlantPhysiology,1998,116(1):319-328.
[2]王涛,杨保,BraeuningA,等.近0.5ka来中国北方干旱半干旱地区的降水变化分析[J].科学通报,2004,49(9):883-887.
[3]FuQ,FengS.Responsesofterrestrialariditytoglobalwarming[J].JournalofGeophysicalResearch(Atmospheres),2014,119(13):7863-7875.
[4]MediciLO,ReinertF,CarvalhoDF,etal.Whataboutkeepingplantswellwatered?[J].EnvironmentalandExperimentalBotany,2014,99:38-42.
[5]中国农业科学院农业信息研究所.黄瓜2016年市场分析及2017年市场预测[EB/OL].(2017-01-22)[2020-10-23].http://jiuban.moa.gov.cn/zwllm/jcyj/201701/t20170122_5461548.htm.
[6]许天颖.我国黄瓜种植规模世界第一,亩产却只有荷兰的1/5[EB/OL].[2020-10-23].http://www.njdaily.cn/2018/1013/1730030.shtml.
[7]李阳,秦智伟,周秀艳,等.黄瓜主要种质资源耐旱性评价[J].北方园艺,2016(20):5-8.
[8]潘俏,张舒玥,沈迪,等.黄瓜耐盐突变体材料的筛选与鉴定[J].南京农业大学学报,2018,41(1):64-70.
[9]顾少涵,潘俏,李子昂,等.黄瓜耐低温突变体材料的筛选与鉴定[J].南京农业大学学报,2017,40(2):219-224.
[10]孟令波.黄瓜(CucumissativusL.)耐高温种质资源筛选及耐高温特性的研究[D].哈尔滨:东北农业大学,2002.
[11]肖凡,蒋景龙,段敏.干旱和复水条件下黄瓜幼苗生长和生理生化的响应[J].南方农业学报,2019,50(10):2241-2248.
[12]梁玉芹,刘子会,曹彩霞,等.水分胁迫下黄瓜生理生化的变化与抗旱性的关系[J].华北农学报,2010,25(增刊1):105-108.
[13]张占军.PEG-6000模拟干旱胁迫下秋地黄瓜萌芽期抗旱性评价[J].甘肃农业科技,2014(5):16-18.
[14]王晶,崔晓锁.温室黄瓜节水耐旱品种筛选试验[J].安徽农业科学,2017,45(1):28-30.
[15]StaubJE,ChungSM,FazioG.Conformityandgeneticrelatednessestimationincropspecieshavinganarrowgeneticbase:thecaseofcucumber[J].PlantBreeding,2005,124(1):44-53.
[16]周玉麗,朱平,胡能兵,等.不同大豆品种发芽期耐旱性评价及耐旱种质筛选[J].大豆科学,2015,34(4):616-623.
[17]郁飞燕,张联合,李艳艳,等.干旱胁迫对水稻种子萌发的影响[J].山东农业科学,2011(8):36-39.
[18]AnjumSA,XieX,WangL,etal.Morphological,physiologicalandbiochemicalresponsesofplantstodroughtstress[J].AfricanJournalofAgriculturalResearch,2011,6(9):2026-2032.[HJ1.6mm]
[19]李丹丹,梁宗锁,杨宗岐,等.外源-5氨基乙酰丙酸对干旱胁迫下紫花苜蓿生理特性及次生代谢物含量的影响[J].西北植物学报,2019,39(10):1827-1834.
[20]马少薇,刘果厚,王蕾,等.干旱胁迫对黄柳雌雄扦插苗生长和生理特性的影响[J].西北植物学报,2019,39(7):1250-1258.
[21]钱瑭璜,雷江丽,庄雪影.华南地区8种常见园林地被植物抗旱性比较研究[J].西北植物学报,2012,32(4):759-766.
[22]王建华,刘鸿先,徐同.超氧物歧化酶(SOD)在植物逆境和衰老生理中的作用[J].植物生理学通讯,1989,1(1):1-7.
[23]赵世杰,许长成,邹琦,等.植物组织中丙二醛测定方法的改进[J].植物生理学通讯,1994,30(3):207-210.
[24]职明星,李秀菊.脯氨酸测定方法的改进[J].河南科技学院学报(自然科学版),2005,33(4):10-12.
[25]曲春香,沈颂东,王雪峰,等.用考马斯亮蓝测定植物粗提液中可溶性蛋白质含量方法的研究[J].苏州大学学报(自然科学版),2006,22(2):82-85.
[26]BaghalianK,AbdoshahS,Khalighi-SigaroodiF,etal.Physiological[KG*2/3]and[KG*2/3]phytochemical[KG*2/3]response[KG*2/3]to[KG*2/3]drought[KG*2/3]stress[KG*2/3]of
[KH*4D]
[HT8.]Germanchamomile(MatricariarecutitaL.)[J].PlantPhysiologyandBiochemistry,2011,49(2):201-207.
[27]ChenCH.Physiologicalresponseofseedlingsofthreeazaleaspeciesofdroughtstressandevaluationofdroughtresistance[J].ScientiaSilvaeSinicae,2011,47(6):48-55.
[28]IraniS,MajidiMM,MirlohiA,etal.Assessmentofdroughttoleranceinsainfoin:physiologicalanddroughttoleranceindices[J].AgronomyJournal,2015,107(5):1771-1781.
[29]姚觉,于晓英,邱收,等.植物抗旱机理研究进展[J].华北农学报,2007,22(增刊1):51-56.
[30]张弢.干旱胁迫对黄瓜幼苗生理指标的影响[J].南方农业学报,2011,42(12):1466-1468.
