6个匍匐剪股颖种质对干旱胁迫的生理响应与耐旱性评价
2021-03-15刘南清林绍艳周兴元
刘南清 林绍艳 周兴元

摘要:采用盆栽模拟干旱胁迫试验的方法,研究5个野生匍匐剪股颖种质(AgrostisstoloniferaL.)生态型与1个进口品种在干旱胁迫期間的生理响应,测定与分析叶色(Color)、相对含水量(RWC)、最大光化学效率(Fv/Fm)、渗透调节(OA)、蒸散率(ER)、相对电导率(EL)、丙二醛(MDA)含量等指标的变化,并用隶属函数法进行耐旱性综合评价。结果表明,随着干旱胁迫时间的增加,匍匐剪股颖的Color、RWC、Fv/Fm及ER持续降低,而叶片的EL、MDA含量则持续升高,这些生理指标在变化上存在显著的生态型差异。干旱胁迫16d,匍匐剪股颖生态型A-LY和A-LS种质与生态型A-DY、A-HZ和A-HS种质之间生理指标对比发现,其中生态型A-LY和A-LS种质的指标Color、RWC、Fv/Fm和OA显著低于生态型A-DY、A-HZ和A-HS种质;生态型A-LY和A-LS种质的指标EL和MDA显著高于生态型A-DY、A-HZ和A-HS种质。采用隶属函数法对6个匍匐剪股颖种质的耐旱性进行耐旱性综合评价,耐旱性排序为A-HS>A-HZ>A-DY>Pen-A4>A-LY>A-LS。
关键词:匍匐剪股颖种质;野生生态型;耐旱性;生理响应;隶属函数综合评价
中图分类号:S688.401文献标志码:A
文章编号:1002-1302(2021)01-0118-08
作者简介:刘南清(1979—),男,广西平南人,博士,副教授,主要从事草坪生产与管理及草坪草育种研究。E-mail:Liunanqing@jsafc.edu.cn。
通信作者:周兴元,博士,教授,主要从事林草植物逆境生理研究。E-mail:Zhouxingyuan@jsafc.edu.cn。
干旱是最为普遍的自然灾害,我国地处东亚地区,是典型的季风气候,季风年际的不稳定性导致我国干旱灾害频发,其发生频次约占总气候灾害频次的1/3,为各项灾害之首[1]。干旱胁迫是植物生长过程中遭遇的最主要的环境抑制因子之一[2],草坪草尤其是冷季型草坪草因其耐旱性不强,因而容易受到干旱胁迫的不利影响[3]。
匍匐剪股颖(AgrostisstoloniferaL.)作为世界上最重要的冷季型草坪草之一,其原产于欧亚大陆。匍匐剪股颖具有耐寒、耐阴、耐低修剪等优点,并兼具柔软细腻、色泽鲜绿、成坪品质高等优质的坪用性状,是当今世界高尔夫球场果岭草坪的最优秀的建坪草种[4]。然而,匍匐剪股颖的耐旱性较差,为获得高质量的匍匐剪股颖草坪,在草坪养护管理过程中须要频繁灌溉,消耗大量的淡水资源,养护成本高。
我国华东地区经济发达,对草坪的质量标准要求高,匍匐剪股颖被广泛应用于园林绿化及高尔夫球场的建造中。当前,我国匍匐剪股颖草种基本从国外进口,耐旱性与耐热性较差,在华东地区因夏秋季干旱胁迫而萎蔫干枯严重,极大降低了草坪的使用价值与使用效果。我国华东地区有着丰富的野生匍匐剪股颖种质资源,对其进行耐旱性研究,明确其耐旱机制,并进行耐旱性综合评价,对挖掘具有强耐旱性的野生匍匐剪股颖种质资源,选育与推广耐旱型匍匐剪股颖新品种,降低草坪养护管理中水资源与能源的消耗,降低养护成本,提倡生态节约型草坪建植与养护理念,都具有重要的意义。
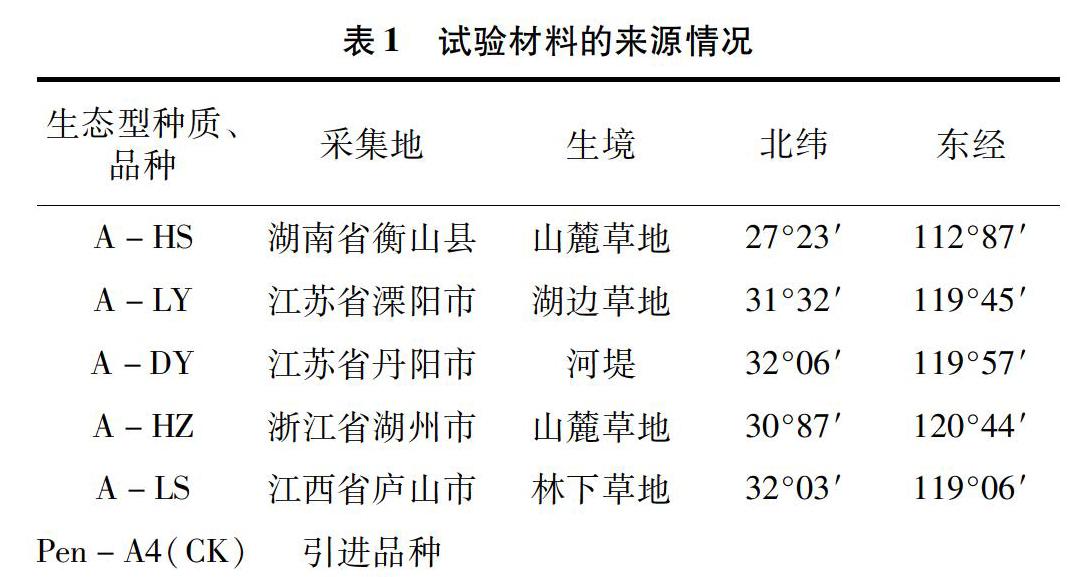
本试验以5个采集于华东地区各省(市)的野生匍匐剪股颖生态型,以及国外引进的品种Pen-A4为材料,采用干旱处理,通过测定叶色(Color)、相对含水量(RWC)、最大光化学效率(Fv/Fm)、渗透调节(OA)、蒸散率(ER)、相对电导率(EL)和丙二醛含量(MDA)等指标,研究干旱胁迫下匍匐剪股颖的生理响应,运用模糊数学隶属函数法综合评价6个匍匐剪股颖的耐旱性,旨在明确匍匐剪股颖的耐旱机制,挖掘耐旱性强的种质资源,为选育耐旱型匍匐剪股颖乡土草种提供依据。
1材料与方法
1.1试验区概况
试验地位于江苏省句容市边城镇江苏农博园草坪种质资源圃(32°01′34.76″N,119°14′25.85″E),海拔为50m。年均气温为15.5℃,全年无霜期为220~240d,土壤为黄棕壤,pH值为7.6,试验地土壤肥力中等,地力均匀。
1.2试验材料
供试材料共6份,2014年采集于华东各省的野生匍匐剪股颖种质资源5份,引进品种1份(表1)。供试材料采集或引进后,于2014年11月种植于种质资源圃,正常水肥管理,所有供试材料均完全成活并生长良好。
1.3试验方法
2017年3月,从种质资源圃挖取6个匍匐剪股颖草块,用水清洗干净根部土壤,随后立刻移栽到充满纯净细河沙的聚氯乙烯(PVC)种植管中(深40cm,管径为12cm),放进江苏农博园的日光温室中栽培,每份供试材料设置浇水对照组与干旱处理组,3次重复。3—5月期间,供试材料放置于温室条件下(10~12h/d的自然光照,昼夜平均温度分别为21、13℃)栽培3个月,期间正常水肥管理,每隔3d修剪1次,修剪高度为6cm。3个月后,把栽培好的试验材料转移进人工气候模拟箱中进行适应性栽培。2017年8月6日,在人工气候模拟箱中开始对植物进行干旱胁迫处理。生长室保持20/15℃的昼夜温度,70%的相对湿度,光照时长为12h/d,以及650mmol/(m2·s)的草坪草冠际高度光照度。
干旱胁迫开始之前,每周给草坪草浇水3次,浇水时,当发现水分从种植管底部流出时即停止浇水,以确保足够的土壤湿度。每周使用50%的霍格兰氏全营养液进行施肥1次,每隔3d用剪刀对草坪进行修剪1次,保持6cm的修剪高度。试验设计2个处理,分别是干旱处理组和对照组。干旱处理组采用自然干旱(即不浇水)的方法,分别于干旱处理0、4、8、12、16d进行各项指标测定,并剪取长势一致的完全展开叶进行指标的分析测定。对照组(即正常浇水)保持每周3次的浇水频率,确保根系土壤水分充足。
1.4观测项目与生理指标的测定
1.4.1叶色
采用目测法测定匍匐剪股颖的叶片色泽,评分标准采用草坪草叶片色澤9级评分法。其中,1级代表分值最低,表示草坪草完全枯黄;9级代表分值最高,表示草坪草叶片含水量充足,色泽墨绿[5]。3次重复。
1.4.2相对含水量[JP3]
RWC计算公式为RWC=100×[(FW-DW)/(TW-DW)][6],FW为叶片的鲜质量;DW指样品叶片在80℃的烘箱内烘72h后叶片的干质量;TW是把叶片完全浸泡在去离子水中,放置于4℃冰箱中,24h后吸饱水后的质量。3次生物学重复。
1.4.3相对电导率
采用李合生的相对电导率测定方法[7]。取样后将叶片用去离子水冲洗3次,并用洁净滤纸吸干表面水分。叶片剪成0.5cm的小片段,准确称取0.5g,装入具塞试管。加30mL去离子水,真空抽气10min,振荡,加塞,在室温下静置1h。3次生物学重复。用电导率仪测得电导率(E1),沸水浴10min,冷却后测得电导率(E2)。计算公式:EL=E1/E2×100%。
1.4.4渗透调节
通过完全复水法来测定OA,把失水的叶片完全浸泡于水中,使叶片完全吸饱水复水,用压汁器压出细胞液,测定叶片饱和渗透压[8]。把吸饱水后的叶片样品放置于液氮中低温急冻处理,然后储存于-20℃的低温冰箱中。测定叶片饱和渗透压时,从低温冰箱中取出叶片样品,用压汁器(FredS.Carver,Wabash,IN)压出叶片细胞液,用与渗透压仪配套的专用滤纸吸足细胞液,把吸足了细胞液的滤纸放在渗透压测定仪(Vapromodel5520;Wescor,Logan,UT)上测定叶片细胞的渗透压(C,单位为mmol/kg)。用公式1MPa=-C·2.58×10-3把单位mmol/kg换算成MPa。测定的浇水对照的叶片渗透压与干旱胁迫处理的叶片的渗透压之差,即为渗透调节OA。3次重复。
1.4.5最大光化学效率
采用叶绿素荧光仪(Fim1500型,美国德克萨斯州休斯敦Dynamax公司生产)测定法测定光化学效率。测定时,用叶绿素荧光测定仪的叶片夹分别夹住各处理的匍匐剪股颖的叶片,关闭叶片夹的透光孔,避光30min处理,读取Fv/Fm测定值[9],3次重复。
1.4.6蒸散率
蒸散率的测定采用质量平衡法进行测定,每隔2d称量1次不同处理的总质量,即不同生态型的匍匐剪股颖的总生物量与其种植柱的质量,计算出2次称量的质量差值,用此质量差值作为蒸散量,蒸散率为单位面积及单位时间内的蒸散量[10]。3次重复。
1.4.7丙二醛含量
MDA含量采用硫代巴比妥酸法测定[11]。剪取叶片鲜样0.5~1.0g,在-80℃的液氮中冷冻。解冻后,加7mL浓度为50mmol/L的磷酸盐缓冲溶液(pH值为7.0),并立即在冰浴中研磨。以上所得提取液放置于4℃于20000g高速离心25min。待所得的上清液与三氯乙酸和硫代巴比妥酸充分反应后,用分光光度计(SpectronicInstruments,NewYork)在532nm与600nm波长下分别测定其吸光度。使用Dhindsa的计算公式[12]计算出MDA的含量。3次重复。
1.5数据处理
不同处理间的方差分析采用SAS软件(9.0版)进行。用Fisher检验法确定处理间差异的显著性,P<0.05表示差异显著。
采用公式X(μ)=(X-Xmin)/(Xmax-Xmin)计算模糊数学隶属函数,公式中的X(μ)为匍匐剪股颖某一耐旱性相关指标的隶属函数值;X为该指标的平均值;Xmax和Xmin分别为该指标的最大值、最小值。最后将匍匐剪股颖各生态型所有性状的具体隶属值加和,加和平均值即代表各生态型各自的耐旱性综合值[13]。
2结果与分析
2.1叶色与叶片相对含水量的变化
2.1.1叶色的变化
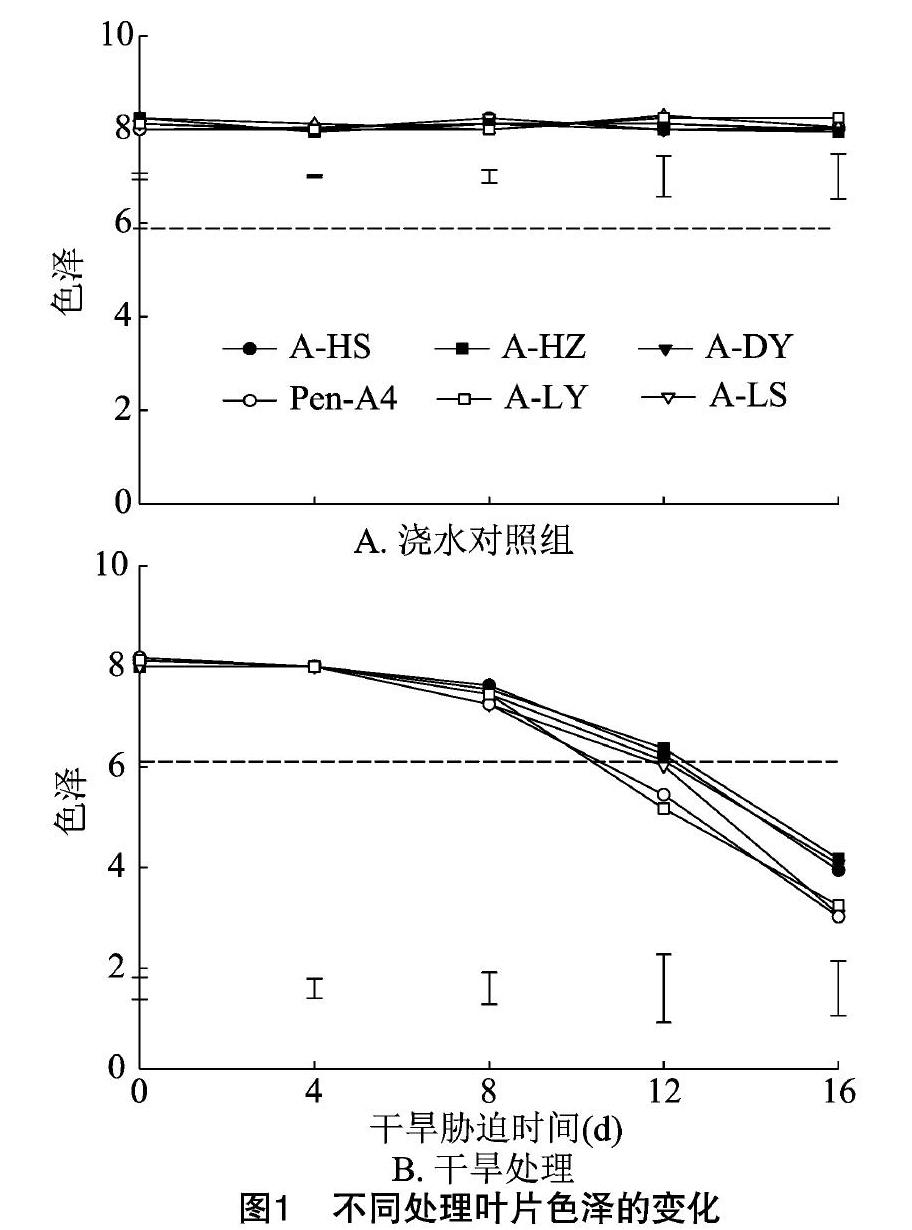
随干旱处理时间的延长,浇水对照组匍匐剪股颖的叶色都保持在7.95~8.30的较高水平,不同生态型种质间无明显差异(图1-a)。
在干旱处理的0~4d,各生态型匍匐剪股颖叶色降低不明显;干旱胁迫至8d时,匍匐剪股颖的叶色开始出现明显的下降,各生态型间的下降幅度差异明显;干旱胁迫12d时,6个匍匐剪股颖叶色都急剧下降,叶色的降低程度在不同生态型间差异明显,野生生态型A-LY与Pen-A4种质的叶色明显低于其他生态型,降低到6.0以下;干旱胁迫16d,A-HS、A-HZ、A-DY、Pen-A4、A-LY和A-LS种质的叶色分别下降47.81%、51.38%、49.85%、61.56%、63.00%、60.00%;在干旱胁迫16d后,A-LY与A-LS种质的叶色一直明显低于A-HS、A-HZ、A-DY种质(图1-b)。
2.1.2叶片相对含水量的变化
浇水对照组在整个试验期间叶片的相对含水量维持在85%~94%之间,不同生态型种质和品种间无明显差异(图2-a)。
干旱胁迫开始的0~4d,匍匐剪股颖叶片的相对含水量下降不明显;干旱胁迫4d至试验结束,匍匐剪股颖各生态型与品种的叶片相对含水量持续下降,在下降速度与下降幅度上,不同生态型种质与品种间差异明显;干旱胁迫16d时,A-HS、A-HZ、A-DY、Pen-A4、A-LY和A-LS种质相对含水量分别下降了46.95%、49.87%、50.75%、55.38%、60.70%、62.39%;野生生态型种质A-HS、A-HZ和A-DY的叶片相对含水量明显高于A-LY、A-LS与Pen-A4种质(图2-b)。
2.2叶片相对电导率与丙二醛含量的变化
2.2.1叶片相对电导率的变化
干旱胁迫期间,浇水对照组匍匐剪股颖的叶片相对电导率维持在较低的水平,不同生态型种质间和品种间差异不明显(图3-a)。
随着干旱胁迫加重,匍匐剪股颖各生态型和品种的叶片相对电导率持续上升,但是在升高速度与程度上各生态型和品种间差异明显;干旱胁迫16d时,A-HS、A-HZ、A-DY、Pen-A4、A-LY和A-LS种质的叶片相对电导率分别为0d的3.21、2.98、3.24、3.22、3.85、4.83倍;干旱8d至试验结束,匍匐剪股颖野生生态型种质A-LY、A-LS与Pen-A4的叶片相对电导率都明显高于A-DY、A-HZ与A-HS种质(图3-b)。
2.2.2叶片丙二醛含量的变化
浇水对照组匍匐剪股颖叶片的MDA含量较低,不同生态型种质与品种间差异不明显(图4-a)。
干旱胁迫期间,匍匐剪股颖叶片的MDA含量持续增加,但是不同生态
型和品种在MDA含量的增加幅度上差异明显;干旱胁迫16d时,A-HS、A-HZ、A-DY、Pen-A4、A-LY和A-LS种质叶片的MDA含量分别是初始的3.76、4.39、5.21、4.83、4.41、5.22倍;匍匐剪股颖野生生态型种质A-LY与A-LS的叶片MDA含量明显高于Pen-A4、A-DY、A-HZ与A-HS种质(图4-b)。
2.3叶片渗透压的变化
干旱胁迫期间,浇水对照组所有匍匐剪股颖叶片的OA稳定保持在0.150~0.242MPa之间,不同生态型种质与品种间差异不明显(图5-a)。
干旱胁迫发生时,6个匍匐剪股颖叶片内的渗透调节都表现出先轻微下降,后快速升高,再快速下降的趋势,在12~16d的干旱胁迫期间,野生生态型种质A-HS、A-HZ与A-DY的OA都明显大于品种Pen-A4与A-LY、A-LS种质(图5-b)。
2.4蒸散率的变化
在浇水对照组中,匍匐剪股颖的蒸散率存在明显的生态型和品种间差异。野生生态型种质A-LY、A-LS、A-DY与品种Pen-A4的蒸散率较高,而A-HS与A-HZ种质的蒸散率则相对较低(图6-a)。
干旱胁迫过程中,6个匍匐剪股颖的蒸散率持续降低,在10~12、14~16d,品种Pen-A4的蒸散率明显高于其他5个匍匐剪股颖野生生态型种质之外;干旱胁迫18~20d,A-HS、A-HZ、A-DY、
Pen-A4、A-LY和A-LS種质蒸散率分别下降至2~4d蒸散率的8.56%、9.13%、8.72%、7.98%、6.22%、6.75%;在其他时期内,匍匐剪股颖的蒸散率并未表现出生态型和品种间的差异(图6-b)。
2.5最大光化学效率的变化
干旱胁迫期间,浇水对照组6个匍匐剪股颖种质的最大光化学效率保持在0.76~0.80的高水平,不同生态型种质和品种间无明显差异(图7-a)。
干旱胁迫下,匍匐剪股颖的最大光化学效率持续下降。在0~8d下降速度缓慢,6个匍匐剪股颖种质间无明显差异。在干旱胁迫的8~16d,最大光化学效率急剧下降,试验结束时,最大光化学效率最低的生态型种质A-LS降到0.35;干旱胁迫16d后,A-HS、A-HZ、A-DY、Pen-A4、A-LY和A-LS种质的最大光化学效率分别下降了38.43%、38.92%、41.31%、41.66%、54.89%、56.87%;匍匐剪股颖各生态型种质和品种间最大光化学效率差异明显,A-HS、A-HZ、A-DY种质和对照品种Pen-A4的最大光化学效率均显著高于A-LY与A-LS种质(图7-b)。
2.6干旱胁迫期间耐旱生理指标的隶属函数法分析
对5个匍匐剪股颖野生生态型种质与品种Pen-A4的叶色、最大光化学效率、相对含水量、渗透调节、相对电导率与丙二醛含量等指标进行隶属函数法计算综合耐旱性,结果表明,5个匍匐剪股颖种质野生生态型与品种Pen-A4间的综合耐旱性差异明显,综合耐旱性排序为A-HS>A-HZ>A-DY>Pen-A4>A-LY>A-LS(表2)。
2.7干旱胁迫期间生理指标相关性分析
相关性分析结果(表3)表明,干旱胁迫下,综合耐旱性X(μ)与Color、Fv/Fm、RWC及OA之间存在显著或极显著正相关,相关系数分别为0.70、0.96、0.95、0.82;X(μ)与EL及MDA之间存在极显著的负相关,相关系数分别为-0.87、-0.94;X(μ)与ER之间呈负相关性,但是相关性不显著。
3讨论与结论
在逆境胁迫发生时,植物细胞膜系统的通透性与膜脂过氧化程度会增加,植物受到逆境胁迫的伤害。在干旱胁迫下,草坪草细胞膜系统的通透性与膜脂过氧化程度升高,膜系统加速解体,草坪草遭受干旱胁迫伤害[14-16]。当前,相对电导率与丙二醛含量作为衡量细胞膜通透性与膜脂过氧化程度的重要指标而被广泛接受[17]。干旱胁迫下,草地早熟禾(PoapratensisL.)[18-20]、高羊茅(FestucaarundinaceaSchreb.)[21-24]、黑麦草(LoliumperenneL.)[24-25]与匍匐剪股颖[26]等草坪草的相对电导率与丙二醛含量都持续升高。光合作用在调节植物耐旱性方面起着至关重要的作用。相关研究表明,当受到干旱胁迫时,草坪草的叶绿素含量快速降低,最大光化学效率与光合作用也因而大幅度降低,草坪草在干旱胁迫下的存活率降低[27-29]。
保持适宜的叶片水分状态对维持叶片的正常生理生化功能尤为重要。当干旱胁迫发生时,能在较长时间内维持叶片适宜的相对含水量的植物,保持其正常生理代谢的可能性更大,存活率也更高,耐旱性更强。渗透调节被认为是诸多植物的耐旱生理机制[30],在草坪草中同样如此[31-32]。在缺水状态下,提高渗透调节有利于维持细胞膨压[33-35]。相关对匍匐剪股颖的耐旱研究发现,干旱胁迫下,一方面,匍匐剪股颖因缺水加剧,相对含水量下降,另一方面,细胞内的多糖、脯氨酸、甜菜碱、多胺等渗透调节物质在干旱胁迫时合成增加,从而维持细胞的渗透调节,降低干旱胁迫伤害[15-16]。研究发现,草坪草植物的叶片蒸散率与草坪草植物的水分利用率具有密切相关性,蒸散率因物种或品种的不同而存在明显的差异[36-37]。相关研究认为,低蒸散率是草坪草节水、耐旱的一个重要性状[38]。然而,相关研究发现,匍匐剪股颖不同品种间的蒸散率仅在水分充足时存在明显差异,而在干旱胁迫下,则品种间的蒸散率差异不明显[39-40]。
本试验中,干旱胁迫发生时,6个匍匐剪股颖种质的叶色、相对含水量、最大光化学效率与蒸散率都持续降低,相对电导率与丙二醛含量则出现持续升高,表明6个匍匐剪股颖种质的外部形态与内部细胞都受到了干旱胁迫的伤害。这可能由于干旱胁迫使得匍匐剪股颖无法从缺水的根系土壤获取足够的水分,植物体内的细胞含水量下降,植株相对含水量降低,细胞膜的流动性降低,膜透性和膜脂过氧化程度均持续上升,因而导致细胞膜系统与细胞解体的加速,伴随着膜系统与细胞的解体,匍匐剪股颖叶片细胞内的叶绿体开始大量解体,叶色快速失绿,叶片最大光化学效率急剧降低。本研究发现,渗透调节在干旱胁迫期间出现了先升后降的变化趋势,在干旱胁迫发生8d后,6个匍匐剪股颖种质的渗透调节同时升高,野生生态型A-HS、A-HZ与A-DY的渗透调节持续升高至干旱胁迫发生12d时。这可能是匍匐剪股颖适应干旱胁迫的一种机制,通过提高渗透调节,阻止水分由细胞内快速流失,保持一定的水合能力,降低干旱伤害。
本研究发现,在干旱胁迫下,匍匐剪股颖的叶色、相对含水量、最大光化学效率、渗透调节、蒸散率、相对电导率、丙二醛含量等7个生理指标的变化存在明显的生态型或品种间的差异。通过隶属函数法对6个匍匐剪股颖种质进行耐旱性综合评价发现,6个匍匐剪股颖种质的耐旱性存在明显的生态型与品种间的差异,综合耐旱性与相关生理指标的相关性分析结果表明,匍匐剪股颖的综合耐旱性与叶色、最大光化学效率、相对含水量、渗透调节等存在显著或极显著的正相关关系,而与相对电导率及丙二醛含量存在极显著的负相关关系,但与蒸散率的相关性不明显[13],这些指标可以作为衡量匍匐剪股颖耐旱性的生理指标。
6个匍匐剪股颖种质的综合耐旱性存在差异,排序为A-HS>A-HZ>A-DY>Pen-A4>A-LY>A-LS。说明在野生匍匐剪股颖种质中存在着比引进品种耐旱性更高的生态型,匍匐剪股颖种质生态型A-HS、A-HZ与A-DY对选育强耐旱型匍匐剪股颖具有较高的价值,可以作为匍匐剪股颖耐旱种质育种的材料。
参考文献:
[1]黄会平.1949—2007年全国干旱灾害特征、成因及减灾对策[J].干旱区资源与环境,2010,24(11):94-98.
[2]曲涛,南志标.作物和牧草对干旱胁迫的响应及机理研究进展[J].草业学报,2008,17(2):126-135.
[3]马祎,王彩云.几种引進冷季型草坪草的生长及抗旱生理指标[J].草业科学,2001,18(2):57-61.
[4]TurgeonAJ.TurfgrassManagemen[M].UpperSddleRiver,NJ:PrenticeHall,2008:84-86.
[5]TurgeonAJ.TurfgrassManagemen[M].4thed.Englewood:PrenticeHall,1996:207-211.
[6]BarrsHD,WeatherleyPE.Are-examinationoftherelativeturgiditytechniqueforestimatingwaterdeficitsinleaves[J].AustralianJournalofBiologicalSciences,1962,15(3):413-428.
[7]李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.
[8]BüssisD,HeinekeD.Acclimationofpotatoplantstopolyethyleneglycol-inducedwaterdeficit:Ⅱ.Contentsandsubcellulardistributionoforganicsolutes[J].JournalofExperimentalBotany,1998,49(325):1361-1370.
[9]MerewitzEB,GianfagnaT,HuangB.Proteinaccumulationinleavesandrootsassociatedwithimproveddroughttoleranceincreepingbentgrassexpressinganiptgeneforcytokininsynthesis[J].JournalofExperimentalBotany,2011,62(15):5311-5333.
[10]MccannSE,HuangB.Evaluationofdroughttoleranceandavoidancetraitsforsixcreepingbentgrasscultivars[J].HorticulturalScience,2008,43(2):519-524.
[11]刘祖祺,张石城.植物抗性生理学[M].北京:中国农业出版社,1994:372.
[12]DhindsaRS.Inhibitionofproteinsynthesisbyproductsoflipidperoxidation[J].Phytochemistry,1982,21(2):309-313.
[13]李俊庆,芮文利,齐敏忠,等.水分胁迫对不同抗旱型花生生长发育及生理特性的影响[J].中国农业气象,1996(1):11-13.
[14]AbrahamEM,HuangBR,BonosSA,etal.EvaluationofdroughtresistanceforTexasbluegrass,Kentuckybluegrass,andtheirhybrids[J].CropScience,2004,44(5):1746-1753.
[15]LiuNQ,ShenYX,HuangBR.Osmoregulantsinvolvedinosmoticadjustmentfordifferentialdroughttoleranceindifferentbentgrassgenotypes[J].JournaloftheAmericanSocietyforHorticulturalScience,2015,140(6):605-613.
[16]LiuNQ,LinSY,HuangBR.Differentialeffectsofglycinebetaineandspermidineonosmoticadjustmentandantioxidantdefensecontributingtoimproveddroughttoleranceincreepingbentgrass[J].JournaloftheAmericanSocietyforHorticulturalScience,2017,142(1):20-26.
[17]RachmilevitchS,DacostaM,HuangBR.Physiologicalandbiochemicalindicatorsforstresstolerance[M].BocaRaton:CrcPress,2006.
[18]李寿田,韩建国,毛培胜.26个草地早熟禾品种苗期抗旱性综合评价[J].草业科学,2012,29(7):1114-1119.
[19]白利国,俞玲,马晖玲.野生草地早熟禾对干旱胁迫的生理响应[J].草原与草坪,2014,34(2):86-91.
[20]王昕慧,孙晓阳,李勋,等.钾素增强干旱胁迫下草地早熟禾生理耐性的机制[J].中国草地学报,2018,40(4):23-29.
[21]HuangBR,FryJD.Rootanatomical,physiological,andmorphologicalresponsestodroughtstressfortallfescuecultivars[J].CropScience,1998,38(4):1017-1022.
[22]DuHM,WangZL,HuangBR.Differentialresponsesoftallfescueandzoysiagrasstoheatanddroughtstress[J].ActaHorticulturae,2008,783:207-213.
[23]謝晓蓉,李唯,刘金荣,等.21个高羊茅品种抗旱抗热性的生理评价[J].草地学报,2009,17(2):202-205.
[24]杨有俊,郑明珠,秦伟志,等.干旱及复水处理下坪用黑麦草和高羊茅抗旱特性比较[J].草业科学,2012,29(3):370-376.
[25]OraeeA,TehranifarA,NezamiA,etal.Effectsofdroughtstressoncoldhardinessofnon-acclimatedviola(Viola×wittrockiana‘IonaGoldwithBlotch)incontrolledconditions[J].ScientiaHorticulturae,2018,238:98-106.
[26]MaXQ,ZhangJ,BurgessP,etal.Interactiveeffectsofmelatoninandcytokininonalleviatingdrought-inducedleafsenescenceincreepingbentgrass(Agrostisstolonifera)[J].EnvironmentalandExperimentalBotany,2018,145:1-11.
[27]刘美君,隋晓青,安沙舟.干旱脱水对狗牙根叶片光合机构的影响[J].草地学报,2018,26(2):441-446.
[28]常红军,秦毓茜.干旱和盐胁迫对草地早熟禾草坪质量及其叶绿素荧光参数的影响[J].西北植物学报,2008,28(9):1850-1855.
[29]杜建雄,师尚礼,刘金荣,等.干旱胁迫和复水对草地早熟禾3个品种生理特性的影响[J].草地学报,2010,18(1):73-77.
[30]BohnertHJ,JensenRG.Strategiesforengineeringwater-stresstoleranceinplants[J].TrendsinBiotechnology,1996,14(3):89-97.
[31]QianYL,FryJD,UphamWS.Rootinganddroughtavoidanceofwarm-seasonturfgrassesandtallfescueinKansas[J].CropScience,1997,37(3):905-910.
[32]DacostaM,HuangBR.Changesincarbonpartitioningandaccumulationpatternsduringdroughtandrecoveryforcolonialbentgrass,creepingbentgrass,andvelvetbentgrass[J].JournaloftheAmericanSocietyforHorticulturalScience,2006,131(4):789-802.
[33]HuangBR,FryJD.Osmoticpotential,sucroselevel,andactivityofsucrosemetabolicenzymesintallfescueinresponsetodeficitirrigation[J].JournaloftheAmericanSocietyforHorticulturalScience,2010,135(6):505-510.
[34]BurgessP,HuangBR.Effectsofsequentialapplicationofplantgrowthregulatorsandosmoregulantsondroughttoleranceofcreepingbentgrass[J].CropScience,2013,54(2):837-844.
[35]黄鑫,杨健,罗新宁.调亏灌溉对棉花生长及渗透调节物质的影响[J].江苏农业科学,2020,48(7):96-101.
[36]郭君,向佐湘.五种草坪草的蒸散量研究[J].作物研究,2009,23(3):197-200.
[37]潘全山,韩建国,王培.五个草地早熟禾品种蒸散量及节水性[J].草地学报,2001,9(3):207-212.
[38]AlvesAAC,SetterTL.Responseofcassavatowaterdeficit:leafareagrowthandabscisicacid[J].CropScience,2000,40(1):131-137.
[39]McCannSE,HuangBR.Evaluationofdroughttoleranceandavoidancetraitsforsixcreepingbentgrasscultivars[J].HortScience,2008,43(2):519-524.
[40]MccannSE,HuangBR.Effectsoftrinexapac-ethylfoliarapplicationoncreepingbentgrassresponsestocombineddroughtandheatstress[J].CropScience,2007,47(5):2121-2128.
