杧果Whirly基因鉴定及其在病菌侵染过程中的表达分析
2021-03-10刘志鑫罗睿雄蒲金基
孙 宇,刘志鑫,叶 子,罗睿雄,蒲金基,张 贺*
(1 海南大学 植物保护学院,海口 570228;2 中国热带农业科学院环境与植物保护研究所/农业农村部热带作物有害生物综合治理重点实验室,海口 571101;3 中国热带农业科学院热带作物品种资源研究所,海口571101)
Whirly蛋白是一类植物特异的转录因子,广泛存在于植物细胞内,主要定位于叶绿体和线粒体中,具有既能与单链DNA结合,也能与RNA结合的功能[1]。Desveaux等[2]于2000年从马铃薯中分离出第一个Whirly家族成员PBF-2,后来又称为StWHY1[3],它可以和cDNA编码的蛋白质都以序列特异性的方式与病原响应基因PR-10a启动子上的顺式应答元件ERE(elicitor response element)以单链形式结合,进而诱导该基因的表达并调节由病原菌引起的信号转导途径[2]。Whirly蛋白家族一般包含3个结构域:N端结构域,是叶绿体或线粒体信号肽和转录激活结构域;非常保守的Whirly结构域,是与单链DNA结合的区域,其中还包含一段细胞核定位信号;C端多变区,是具有自我调节的结构域,可调节单链DNA和Whirly蛋白的结合活性[3-4]。随后,在拟南芥[1]、玉米[5-6]和番茄[7]等植物中[8-11]陆续鉴定到Whirly基因家族成员的存在,广泛的分布表明它们可能在植物的生长发育与生理过程中起着非常重要的作用。
Whirly蛋白功能发挥通过依赖于单链DNA的碱基及碱基与疏水氨基酸残基的堆叠和疏水键作用进行结合,还依赖其四倍体结构[4,11]。目前已知Whirly蛋白能与4种核酸序列相结合:端粒末端序列[12]、ERE元件[2,13-14]、AtKP1基因启动子上的KPRE元件[15]、WRKY53基因启动子上的ERE-like基序与AT富集区序列[16]。除了ERE-like和AT富集区的序列以外,其余3种核酸序列之间无明显的相似性。近年来的研究表明,拟南芥和马铃薯中的Whirly蛋白家族具有调节防御基因表达的功能[3],其可能在防御反应之外的过程中发挥作用,也可能在叶绿体与细胞核中发挥作用。Whirly蛋白不仅在细胞核内参与水杨酸依赖的抗病信号转导[2,12]、调节端粒结构稳定[14]和调节叶片衰老[16-18],而且在质体和线粒体内能够维持其基因组稳定[19]、DNA损伤修复[20]、调节质体基因的表达[5,19-22]和控制角果的发育[18],另外在胚胎发育和种子萌发[6,23]等方面也发挥着重要的功能。Whirly基因不仅在非生物胁迫中发挥作用,在生物胁迫中也起着积极调节作用,比如番茄SlWHY2在转基因烟草中过表达可对青枯病菌侵染表现出较强的抗性[7],辣椒中CaWHY2的表达受疫病的诱导,在2 h时其表达量达到最高值,为对照的2.3倍[10]。
杧果(Mangiferaindica),是世界著名的热带水果之一,在热带和亚热带地区的100多个国家和地区中分布,包括中国的华南热区[24],是热区重要的农业经济作物,也是农民脱贫致富的“金杧果”和“致富树”。随着2020年杧果全基因组测序的完成[25],杧果基因功能的研究进程明显加快,然而目前国内外对杧果Whirly基因家族的研究还未见报道。为此,本研究对杧果Whirly基因家族成员的序列特征及其表达特性进行分析,以期为进一步研究Whirly家族成员在杧果生长发育中的功能和作用机制提供理论依据。
1 材料和方法
1.1 杧果病害胁迫处理
杧果嫁接苗由农业农村部海南儋州杧果种质资源圃提供(品种为‘贵妃杧’)。将株高约60 cm均匀一致的杧果嫁接苗移栽到花盆内,以室温25 ℃、相对湿度70%~90%、光/暗周期为12 h/12 h条件下进行病害胁迫处理。设置Cg与Xcm 2个处理,每个处理3株苗,0 h处理作为对照组。以分生孢子浓度为2×106个/mL的胶孢炭疽菌分生孢子悬浮液、浓度为2×107cfu 的细菌性黑斑病菌悬浮液分别对杧果嫁接苗叶片均匀喷雾,取样时间分别为处理后0、3、6、12、24、48、72 h,将杧果叶片剪碎,液氮速冻,保存在-80 ℃下待用。
1.2 方 法
1.2.1 杧果全基因组数据的来源杧果全基因组数据由NCBI(PRJNA487154)公布得知,玉米(Zeamays)、烟草(Nicotianatabacum)、拟南芥(Arabidopsisthaliana)、苹果(Malusdomestica)、番茄(Solanumlycopersicum)、菠萝(Ananascomosus)、水稻(Oryzasativa)、木薯(Manihotesculenta)、毛果杨(Populustrichocarpa)9个物种的基因组和Whirly基因家族信息分别来源于NCBI(https://www.ncbi.nlm.nih.gov/)和PlantTFDB(http://planttfdb.gao-lab.org/)数据库。
1.2.2 杧果Whirly基因家族成员的鉴定从拟南芥数据库获取AtWHY1、AtWHY2和AtWHY3氨基酸序列,并将其作为查询对象在杧果基因组数据库中执行Blastp比较,将E值设置为10-5。使用Pfam工具 (http://pfam.xfam.org/)检测得到的杧果蛋白序列结构域,进一步去除没有典型Whirly结构域的蛋白序列,最终得到所有杧果Whirly基因家族成员[26]。其他9个代表物种中的Whirly基因家族成员采用类似的方法进行筛选。
1.2.3 杧果Whirly基因家族的生物信息学分析通过使用ProtParam在线分析工具(http//web.Expasy.org/protpasam/)预测氨基酸数、蛋白质的相对分子质量、等电点、稳定性指数、脂肪系数、总平均亲水性等理化性质[27]。利用在线软件MEME(http://meme-suite.org/tools/meme)对杧果基因家族的蛋白质保守基序进行分析[28],再通过DNAMAN 6.0软件进行高同源蛋白的氨基酸序列比对。通过在线工具SOPMA(https://www.expasy.org/)预测蛋白质的二级结构,利用NCBI Conserved Domain Search预测基因的保守结构域,SWISS-MODEL(https://swissmodel.expasy.org/)对蛋白质保守域的四聚体结构进行预测。
1.2.4 系统进化树分析利用ClustalW程序对杧果和9个物种中的Whirly基因家族成员的氨基酸序列进行多序列比对。获得Whirly蛋白序列后使用MEGA 7.0软件,通过邻近法构建进化树,其中校验参数Bootstrap值设置为1 000次重复[29]。
1.2.5 RNA提取及cDNA合成对杧果叶片总RNA进行提取,采用了RNAprep Pure多糖多酚植物总RNA提取试剂盒(天根生化科技有限公司)中的方法提取,使用超微量紫外分光光度计(Nano-drop2000C型)测定总RNA的浓度,-80 ℃保存备用。利用RevertAid First Strand cDNA Synthesis Kit试剂盒反转录成第1链cDNA[30]。
1.2.6 杧果Whirly基因家族表达分析为研究两种病原菌胁迫下杧果Whirly家族基因的差异表达情况,利用Quant Studio 6 Flex的实时荧光定量PCR检测系统,采用UltraSYBR Mixture 试剂盒(北京康为世纪)作为荧光试剂进行实时荧光定量PCR操作。建立20 μL的反应体系,每个处理样本设置3个重复。qRT-PCR反应程序为:95 ℃预变性10 min;95 ℃变性15 s,60 ℃退火/延伸1 min,45个循环;在60 ℃时收集荧光信号。根据MiWHY1、MiWHY2和MiWHY3基因序列设计实时荧光定量PCR引物MiWHY1-F / MiWHY1-R、MiWHY2-F / MiWHY2-R和MiWHY3-F / MiWHY3-R,以杧果Mi18S作为内参基因,设计引物Mi18S-F / Mi18S-R(表1)。通过实时荧光定量PCR仪软件(Quant StudioTM 6 Flex)获得各个样品的Ct值,以0 hpi的表达量为对照,运用2-ΔΔCt法进行数据统计,确定MiWHY1、MiWHY2和MiWHY3的相对表达量[31]。最后,将qRT-PCR产物经2%的琼脂糖凝胶进行电泳,分析两种病原菌胁迫下不同时间点的基因表达差异。
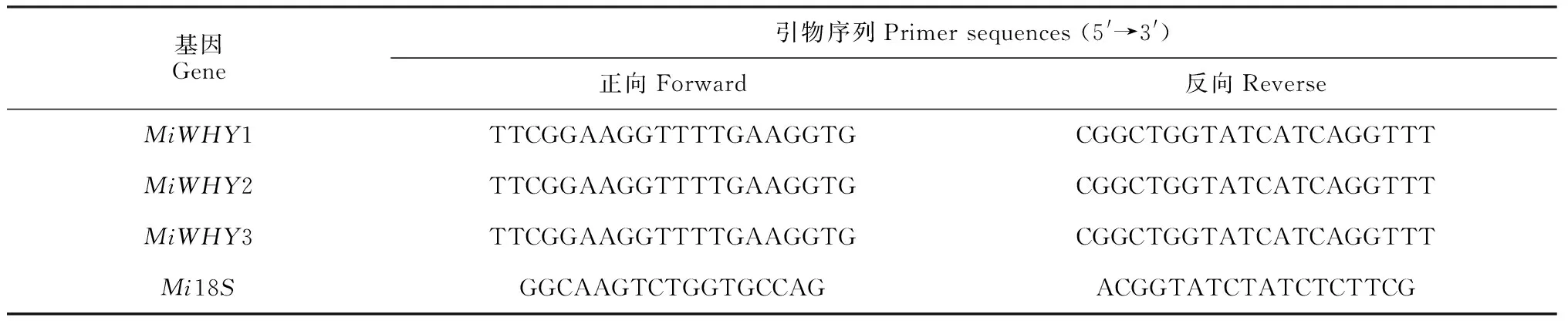
表1 qRT-PCR引物序列
2 结果与分析
2.1 全基因组水平鉴定杧果Whirly基因家族成员
在杧果基因组数据库中进行Blastp检索,去除无典型Whirly结构域的冗余序列和同一基因的非主要转录形式以后,发现杧果Whirly基因家族有3个成员,分别命名为MiWHY1、MiWHY2和MiWHY3。通过生物学在线分析工具ProtParam (http:web.sxpasy.org/protparam/)对杧果Whirly基因家族3个成员MiWHY1、MiWHY2和MiWHY3氨基酸序列的理化性质进行了分析,结果表明(表2),MiWHY1氨基酸数为180个,蛋白分子量为20 294.28 Da,蛋白等电点为9.32,外显子数为6,亲水性平均数为-0.19,脂溶指数为92.06,不稳定指数为45.32;MiWHY2氨基酸数为217个,蛋白分子量为24 512.89 Da,蛋白等电点为9.18,亲水性平均数为-0.447,脂溶指数为68.66,不稳定指数为55.66;MiWHY3氨基酸数为236个,蛋白分子量为26 260.89 Da,蛋白等电点为9.54,外显子数为8,亲水性平均数为-0.285,脂肪系数为75.64,不稳定指数为42.44。因此,预测这3个蛋白均为不稳定亲水碱性蛋白。
2.2 杧果Whirly蛋白系统进化树分析
为了更好地了解杧果Whirly基因家族的系统发育关系,将杧果(3个)、玉米(6个)、烟草(19个)、拟南芥(4个)、苹果(3个)、番茄(2个)、菠萝(1个)、水稻(2个)、木薯(3个)、毛果杨(6个)Whirly基因的蛋白序列通过BLAST工具进行蛋白质序列分析,利用ClustalW程序和MEGA 7.0软件构建3个杧果Whirly基因与其他物种(共9种)植物Whirly基因的蛋白序列系统进化树(图1),基于蛋白序列构建系统发育树显示,Whirly家族蛋白具有多样性,MiWHY1与木薯(Manes.01G189300.1)亲缘关系最近,MiWHY2与毛果杨(Potri.010G092500.1)亲缘关系最近,MiWHY3与番茄(Solyc11g044750.1.1)亲缘关系最近,对于亲缘关系较近的蛋白质,推测它们具有相似或相近的生物学功能。
2.3 杧果Whirly蛋白保守基序分析
基于在线软件MEME对杧果MiWHY1(GWHPABLA011035)、MiWHY2(GWHPABLA032351)、MiWHY3(GWHPABLA034174);番茄PtWHY1(Solyc05g007100.2.1)、PtWHY2(Solyc11g044750.1.1);毛果杨PtWHY1(Potri.003G048700.1)、PtWHY2(Potri.003G048700.2)、PtWHY3(Potri.008G149100.1)、PtWHY4(Potri.008G149100.2)、PtWHY5(Potri.008G149100.3)、PtWHY6(Potri.010G092500.1);木薯MeWHY1(Manes.01G189300.1)、MeWHY2(Manes.02G200100.1)、MeWHY3(Manes.18G108600.1)基因家族的蛋白质保守基序进行分析,发现MiWHY1、MiWHY2和MiWHY3与番茄、毛果杨、木薯的Whirly基因家族中鉴定出3个保守基序,其中共有1个高度保守的基序Motif1,Motif1再通过DNAMAN进行多序列比对(图2)。保守基序Motif1含有49个氨基酸,包含20个百分之百保守的序列,杧果MiWHY1的保守基序位置为34~82;MiWHY2的保守基序位置为125~173;MiWHY3的保守基序位置为91~139。通过基序分析发现,Whirly基因家族在进化上结构具有保守性。

图1 10个Whirly蛋白的系统进化树Fig.1 Phylogenetic tree constructed from 10 Whirly proteins
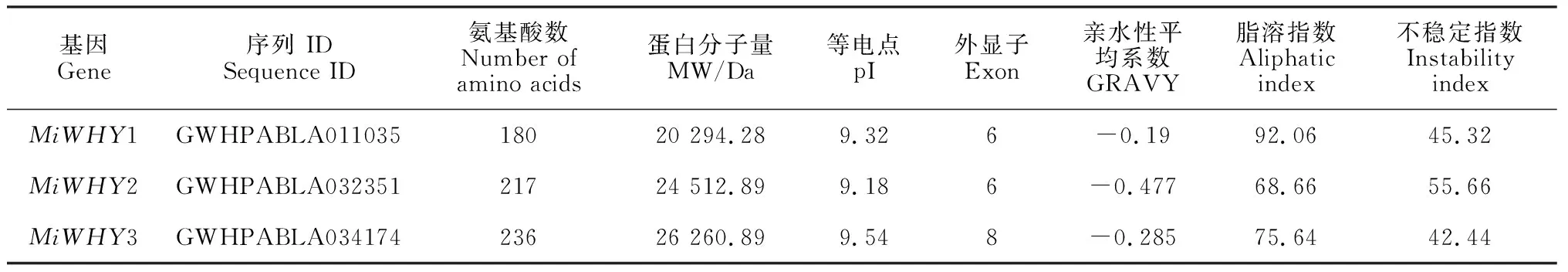
表2 杧果Whirly基因家族成员的基本信息
2.4 杧果Whirly蛋白二级结构预测
通过生物学在线分析工具SOPMA对MiWHY1、MiWHY2和MiWHY3蛋白的二级结构预测分析结果表明(图3),无规则卷曲(45.00%)和α-螺旋(30.00%)是MiWHY1蛋白二级结构主要元件,其次是延伸链(19.44%)和β-转角(5.56%);无规则卷曲(47.47%)和延伸链(26.27%)是MiWHY2蛋白二级结构主要元件,其次是α-螺旋(20.28%)和β-转角(5.99%);无规则卷曲(42.37%)和α-螺旋(29.66%)是MiWHY3蛋白二级结构主要元件,其次是延伸链(22.03%)和β-转角(5.93%)。

MiWHY1~MiWHY3. 杧果;SlWHY1、SlWHY2. 番茄;PtWHY1~PtWHY6. 毛果杨;MeWHY1~MeWHY3. 木薯图2 杧果与其他物种Whirly家族保守基序的多重序列比对MiWHY1-MiWHY3. Mangifera indica;SlWHY1,SlWHY2. Solanum lycopersicum;PtWHY1-PtWHY6. Populus trichocarpa;MeWHY1-MeWHY3. Manihot esculentaFig.2 Multiple sequence alignment of conserved motifs of Whirly family in mango and others

A. MiWHY1;B. MiWHY2;C. MiWHY3;横坐标表示氨基酸位点图3 杧果Whirly家族的二级结构预测The abscissa indicates the position of the amino acid sequenceFig.3 Secondary structure prediction of Whirly family in mango
2.5 杧果Whirly蛋白保守结构域分析
通过NCBI Conserved Domain Search对杧果Whirly家族和马铃薯StWHY1蛋白质序列进行保守结构域分析,发现杧果Whirly家族成员与马铃薯StWHY1含有的保守结构域相同,均含有Whirly超家族保守域(图4)。其中MiWHY1蛋白的Whirly超家族保守域位置为24~139(图4,A);MiWHY2蛋白的Whirly超家族保守域位置为95~212(图4,B);MiWHY3蛋白的Whirly超家族保守域位置为61~196(图4,C); StWHY1蛋白的Whirly超家族保守域位置为98~233(图4,D)。
基于同源建模原理,通过SWISS-MODEL软件对杧果MiWHY1、MiWHY2和MiWHY3蛋白保守结构域的四聚体结构进行预测(图5,A~C),预测结果显示,它们三者的结构基本一致。其中,MiWHY1、MiWHY2和MiWHY3四聚体结构与马铃薯(图5,D)Whirly蛋白的四聚体结构类似,根据结构决定功能的原理,推测MiWHY1、MiWHY2和MiWHY3四聚体在功能上也与马铃薯Whirly蛋白的四聚体具有相似性。
2.6 杧果Whirly基因家族胁迫响应表达分析
以杧果Mi18S作为内参基因,0 h处理作为对照组。利用qRT-PCR检测并分析了两种病原菌侵染过程中杧果Whirly家族基因的表达程度。其中,杧果胶孢炭疽菌(Cg)侵染过程中(图6,A1),MiWHY1、MiWHY2和MiWHY3的相对表达量均显著上调,并在24 h时达到最大值,其中MiWHY1的相对表达量达到对照的74.9倍,MiWHY2的相对表达量达到对照的15.5倍,MiWHY3的相对表达量达到对照的6.8倍,整个侵染过程中MiWHY1的相对表达量均高于MiWHY2和MiWHY3;杧果细菌性黑斑病菌(Xcm)侵染过程中(图6,B1),MiWHY1、MiWHY2和MiWHY3的相对表达量在处理12 h前明显下调,MiWHY1、MiWHY3相对表达量在12 h后显著上调,随后下调。将qRT-PCR产物经2%的琼脂糖凝胶进行电泳(图6,A2、B2),结果表明,在不同病原菌侵染过程中,MiWHY1、MiWHY2和MiWHY3的相对表达量与对照差异性显著。
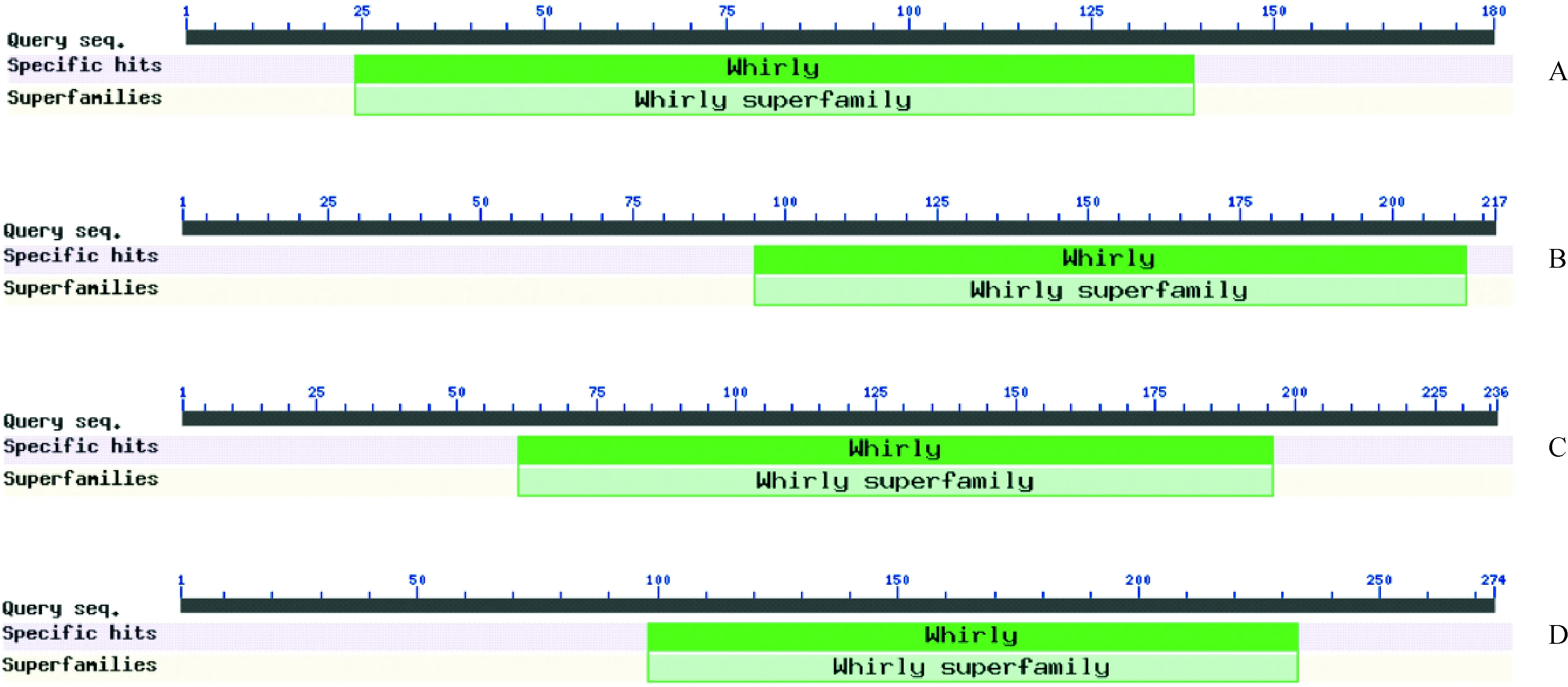
A. MiWHY1; B. MiWHY2; C. MiWHY3;D. StWHY1;Query seq.查询序列;Specific hits.特异位点;Superfamilies.超家族图4 杧果Whirly蛋白保守结构域分析Fig.4 Conserved domain analysis of Whirly proteins in mango
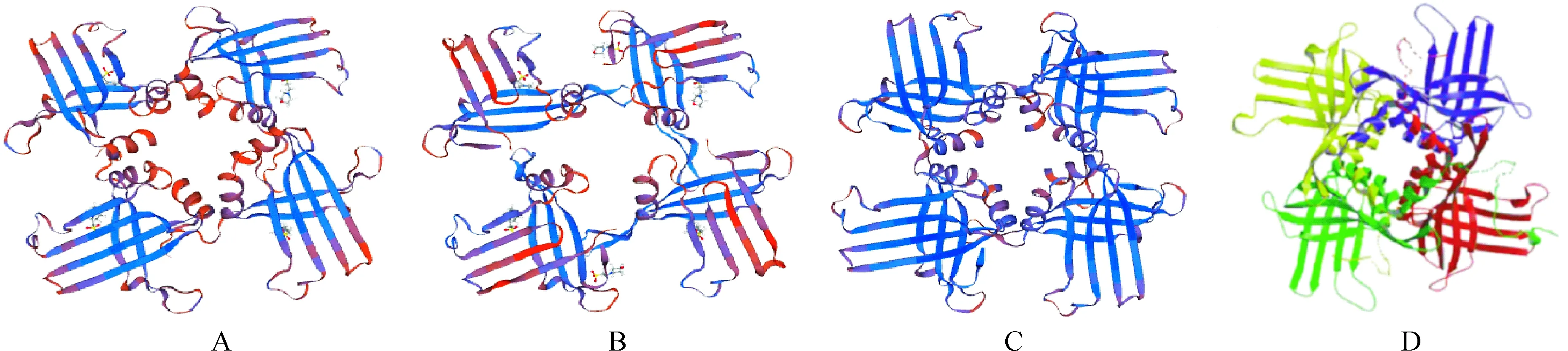
A. MiWHY1;B.MiWHY2;C. MiWHY3;D. 马铃薯 Potato图5 杧果Whirly蛋白保守域四聚体结构预测与马铃薯[4,11]Whirly蛋白四聚体结构比较Fig.5 Comparison of the conserved domain tetramer structure of mango Whirly proteins with the structure of potato Whirly protein tetramer
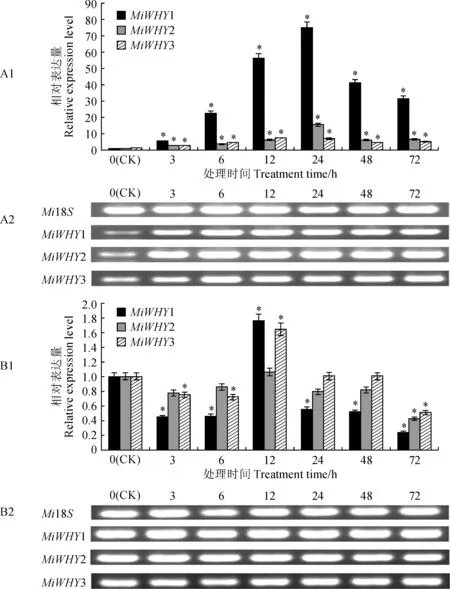
A. 胶孢炭疽菌(Cg);B. 细菌性黑斑病菌(Xcm);*表示处理与对照在0.05水平上差异显著图6 杧果Whirly基因家族在不同病原菌胁迫下的表达与PCR产物电泳图A. Cg;B. Xcm;* Indicates significant difference between treatments and control (CK) at 0.05 levelFig.6 Expression analysis of Whirly gene family in mango under different pathogen stress treatments and PCR product electrophoresis
3 讨 论
基因组测序技术的提高和分子生物学研究的深入,为植物基因家族的鉴定、功能基因的挖掘提供了基础,已经成功对大量植物全基因组进行测序。Whirly家族是一类植物特异的转录因子,广泛存在于植物细胞内。叶绿体、线粒体与细胞核是植物3个具有遗传信息DNA的细胞器,保护DNA与维持遗传信息的有序表达是植物生存与发育的关键,Whirly蛋白既定位于质体又定位于细胞核,在质体与细胞核中发挥着重要作用[4]。Whirly蛋白的结构与生理功能十分复杂,尽管Whirly基因在双子叶草本十字花科的模式植物拟南芥中的分子机制已经得到了阐明[4,11],但对丰富多彩的植物而言,仍很局限,仅仅是家族成员的数量上就差异很大,如CAM植物菠萝有1个成员、茄科烟草有19个成员,家族成员的多样性也暗示着其功能多种多样。本研究的杧果是一种典型的漆树科果树,参照首次报道的马铃薯PBF-2(StWHY1)Whirly蛋白的四聚体结构特点[2-3]和最为保守的Whirly结构域序列,通过全基因组鉴定确定杧果有3个Whirly基因家族成员,均含有1个公认的最为保守的Whirly结构域[3-4],四聚体结构与马铃薯Whirly蛋白四聚体结构具有高度相似性,推测它们在功能上具有一致性。Desveaux等[2]研究发现,马铃薯PBF-2四聚体由4个p24蛋白通过中心的螺旋-环-螺旋连接在一起,C4对称分布形成,形态呈螺旋状。此外,Whirly蛋白四聚体的β-片层结构辐射向外,β-片层的边缘及之间起到与单链DNA结合的作用,而β-片层上部则没有单链DNA的结合,每个蛋白单体只可以结合9个核苷酸;3个α-螺旋向四聚体中心汇聚,从而形成一个直径为0.8 nm且具有疏水功能的中心空穴在蛋白质表面,有害物质有可能被Whirly蛋白利用中心空穴进行储存,最终保护DNA免受胁迫的伤害[4]。
中国杧果病害危害严重,常见的病害有炭疽病、细菌性黑斑病、白粉病、蒂腐病、疮痂病等[27],其中,杧果胶孢炭疽病与细菌性黑斑病危害杧果最为严重[28-29]。为了研究杧果Whirly基因是否受胶孢炭疽菌与细菌性黑斑病菌侵染的诱导表达,本研究分别喷雾接种两种病原菌于杧果叶片上,并通过qRT-PCR技术揭示胶孢炭疽菌与细菌性黑斑病菌侵染过程中杧果Whirly家族基因的相对表达量。发现在不同病原菌侵染过程中,MiWHY1、MiWHY2和MiWHY3的相对表达量与对照差异性显著,如MiWHY1、MiWHY2和MiWHY3在胶孢炭疽菌侵染后的3~72 h内均显著性上调表达,表明杧果Whirly基因响应了病菌的侵染。Whirly基因的研究大多集中在模式植物拟南芥Whirly蛋白的结构和功能上[1]以及干旱[7]、低温[10]、盐[11]等非生物胁迫对基因表达水平的影响方面,也有前人在研究中提到Whirly基因参与转基因烟草青枯病菌[7]、辣椒疫霉菌[10]等生物胁迫的调控。本研究所用的两种病原菌分别属于刺盘孢属真菌、黄单胞菌属细菌,这两个属是生产中较为重要的病原菌,且Whirly基因响应疫霉菌的侵染[10],这暗示着Whirly基因对多种病原菌的侵染均有响应,可作为研究植物抗病机制的候选基因。
