2年生云南松实生苗生物量家系间变异研究*
2021-02-24魏巍蔡年辉李根前郑畹许玉兰李孙玲陈诗王磊孙睿
魏巍,蔡年辉,李根前,郑畹,许玉兰,李孙玲,陈诗,王磊,孙睿
(1.云南省林业和草原科学院,云南 昆明 650201;2.西南林业大学西南山地森林资源保育与利用教育部重点实验室,云南 昆明 650224)
植物生长受环境中可用资源的限制和影响,理想的生物量分配理论假定植物将生物量分配到能够获得最受限制资源的器官,以便实现最快的生长[1]。浓密的下层植物或干旱贫瘠的生境可以影响更新层幼苗的生存,而苗木生物量的分配策略,决定了其竞争能力和将来的生长状况[2]。特定条件下,苗木对各构件器官生物量的投入和比例平衡能力,可视作一组苗木质量评价指标或待选择的优良性状。
云南松(Pinusyunnanensis)具有适应性强、耐干旱瘠薄等优良特性,是中国西南地区人工造林的重要先锋树种,该树种占其主要分布区云南省全省林地面积的19.56%、森林总蓄积量的14.28%[3-5]。但是其天然更新的幼苗均存在“蹲苗期”,即幼苗地上部分生长较慢,很大程度影响着造林成活率和保存率[6]。优质壮苗是营造培育优质高产人工林的基础[7],实际林业生产中一般根据苗高、地径或综合的苗木质量指标对苗木进行分级使用[8]。不同层次的选育工作和苗木分类生产需强化具有生物学特性的变异规律研究,摸清遗传变异规律和主要性状的遗传参数,从而选出不同生态区的优良种源、林分和家系[9]。当前,云南松的遗传改良水平仍处于较为初级的阶段,良种使用率尤其优良家系种苗使用率还较低。进行各层次遗传变异的测定势在必行,而家系层次的遗传变异研究是遗传改良工作的重要组成部分。
因此,本研究对20个半同胞家系的云南松实生苗生物量累积与分配在家系层次的变异规律进行研究,以期为科学评价云南松苗木质量和发挥遗传改良的潜在增益提供理论依据。
1 材料与方法
1.1 试验材料
本试验所用半同胞家系云南松种子采自云南省永仁县林业局国营白马河林场云南松母树林基地内20个云南松健康单株,每株之间的最小水平距离大于100 m,家系平均千粒重和标准差为15.036±2.676g。
1.2 试验设计
本研究采用单因素对比设计,除了种子来自不同的母树外,其他条件均相同。田间排列采用完全区组法,并结合顺序错位法进行排列,设3次重复,每次重复由1个区组组成,1个区组划分为20个小区,每个家系为1个小区,完整试验共包含60个小区。
1.3 育苗方法及苗木管理
试验地设置在西南林业大学育苗圃(102°45′41″ E,25°04′00″ N),海拔1 945 m,属北亚热带半湿润高原季风气候,年平均气温14.7 ℃,年降水量700~1 100 mm,年平均相对湿度68.2%。苗床采用高床设计,宽1.0 m,下层生土,播种前2 d喷洒0.5%高锰酸钾溶液及10.0%多菌灵溶液对下层土消毒,上层铺4~5 cm取自昆明树木园的云南松林表土。按照DB53/T249-2008《林木育苗技术规程》[10]进行种子调制处理并用马勃菌拌种。育苗当年的5月按设计进行点播,每个家系播种14行、每行16株,株行距皆为5 cm×5 cm。播种后采用0.5~1.0 cm厚细粒云南松林下土覆盖,喷灌装置雾状浇水20 min,土壤湿透盖塑料拱膜。苗期第1—2月保持苗床土壤湿润无杂草。大部分苗木长出真叶前,为防止苗木灼伤或猝倒,根据膜内温度定期开膜透风降温,此后去除薄膜。为预防常见病害,在苗木完全木质化前每隔20~30 d用0.5%多菌灵溶液喷洒1次,遇虫害则用1%敌百虫溶液喷洒。
1.4 数据调查
到2 a生苗木生长结束时,即播种后第2年12月底,从每个家系中随机抽取20株苗木,采用全挖法测定生物量。挖出后每株分根、茎、叶不同器官按家系和单株编号带回实验室,105 ℃杀青30 min,80 ℃恒温下烘干至恒定质量,精确至0.001 g。
1.5 家系生物量变异及遗传特性
1.5.1 方差分量百分比
方差分量百分比可以体现数据集变异来源,本研究借鉴群体或居群遗传多样性分析手段[11-14],将其应用在家系层面变异特性分析。方差分量百分比Pσf可以表示家系变异占总变异的比例。
Pσf=σf/σt
①
式中:Pσf为方差分量百分比,σf和σt分别为家系间和总体的方差分量。
1.5.2 表型分化系数
表型分化系数Vσf可以表示家系间表型性状变异的值。
Vσf=σf/(σf+σe)
②
式中:σe为家系内的方差分量,即环境方差分量。
1.5.3 变异系数
变异系数(coefficient of variation,下文皆以“CV”表示)是表示一组数据变异程度大小的指标之一,为标准差S与平均数的比值的百分数,可以反映单位均值上的离散程度,常用在2个总体均值不等的离散程度的比较上。由于不同家系表型性状指标的均值有所差异,本研究采用CV来辅助评价不同家系各表型性状的变异程度。根据式③和式④计算不同家系各表型性状的平均值和标准差S,并依式⑤计算CV。
③
④

⑤
式中:x为生物量指标实测值,n为每个家系测定的株数。
1.5.4 半同胞家系遗传力
苗木的表型性状传递给其后代的潜力称为遗传传递力或遗传力,本研究采用的遗传力估算方法,如式⑥:
⑥
式中:Sf为家系间离差平方和,Se为家系内离差平方和,r为供测家系总数,n为单个生物量指标供测样品的总数,F值为家系间和家系内供测生物量的均方之比值。
1.5.5 遗传增益
遗传增益(△G)在林木遗传育种的分析应用中实际上是表示人工选择取得改良效果的指标之一[15],即入选亲本的子代平均表现值距亲本群体平均型值的离差R与亲本群体平均型值的百分比。根据代算可得:
⑦
式中:S′为选择差,hf2为遗传力。本研究借鉴林木遗传育种遗传增益△G这个指标,并假设选择后的选择差为家系表型性状的1倍标准差,估算在此选择强度下的遗传增益,模拟选择后的改良效果。试验数据用Excel 2013进行统计,SPSS 13.0进行家系生物量指标的分析比较。
2 结果与分析
2.1 2 a生实生苗生物量累积、分配与生产力关系
2 a生实生苗生产力与各构件器官生物量的累积密切相关,由图1可知,实生苗各构件器官生物量累积量随总生物量变大而呈现明显的增加趋势。

图1 2 a生苗木不同构件器官生物量累积量与总生物量大小的关系Fig.1 The relations between modular biomass effort and total biomass of two-year P.yunnanensis
除生物量的累积外,不同构件器官的生物量分配比例还随总生物量的变化呈现一定的规律。由图2可知,如总生物量分配于针叶和茎生长的比例分布范围,随苗木总生物量的增大而逐渐减小,生物量较高的实生苗,针叶生物量分配的范围仅在50%附近,茎生物量分配的范围在40%附近;生物量较低的实生苗,其根系的分配比例较小,高生产力的苗木根系分配比例普遍高于生产力较低的苗木;不同生产力苗木的地上与地下生物量之比有所不同,较低生产力苗木的地上地下生物量比普遍高于较高生产力的苗木,较高生产力的苗木,其地上地下生物量比大部分处于较低水平,在5~10 g之内分布广泛。

图2 2 a生苗木不同构件器官生物量分配比例与总生物量大小的关系Fig.2 The relations between modular biomass effort ratio and total biomass of two-year P.yunnanensis
2.2 2 a生实生苗生物量变异概况
2 a生苗木各构件器官生物量累积量分别为,根干重(0.774±0.281)g、茎干重(3.119±1.488)g、叶干重(3.609±1.586)g、地上干重(6.728±2.908)g、全株干重(7.503±3.425)g。多重比较结果表明(表1),在P<0.05的显著水平下各构件器官生物量累积量可分为2组,全株干重最大的为11号、12号、14号、16号家系,最小的为1号、6号、8号家系;地上干重最大的为12号、14号、16号家系,最小的为1号、6号、8号家系;径干重最大的为2号、8号、11号、15号家系,最小的为6号家系;叶干重最大的为2号、10号、11号、12号、14号、16号家系,最小的为1号家系;根干重最大的为12号、16号家系,最小的为1号家系。由此可知,12号家系生物量积累最多,6号家系生物量积累最少。

表1 2 a生云南松各家系生物量累积Tab.1 Biomass effort among families of two-year P.yunnanensis g
2 a生苗木各构件器官生物量分配比例分别为,根分配(9.32±3.86)%、茎分配(41.75±7.62)%、叶分配(48.93±7.68)%、地上地下生物量比(13.96±5.43)。多重比较结果(表2)表明,在P<0.05的显著水平下根系生物量分配比例可分为2组,最大为12号、16号家系,最小为1号家系。在P<0.05的显著水平下,茎分配、叶分配和地上地下生物量比可以分为3组。径分配最大的为1号、3号家系,最小的为6号、12号、16号家系;叶分配最大的为10号家系,最小的为17号家系;地上地下生物量比最大的为5号、8号、9号家系,最小的为12号家系。

表2 2 a生云南松各家系生物量分配Tab.2 Biomass allocation among families of two-year P.yunnanensis
2.3 2 a生实生苗生物量变异来源
如表3所示,各构件器官和总生物量的累积量在家系间的方差分量占总方差的百分比范围在12.770 3%~33.028 5%,表明其生物量累积量在家系间有一定程度的变异,其中叶干重变异受家系层次的影响最小,茎干重受家系层次的影响最大。总生物量对各构件器官生长分配比例的方差分量占总方差的百分比范围在17.7424%~39.0348%,其中,家系对根分配变异的影响最大,对地上地下生物量比的变异影响最小。
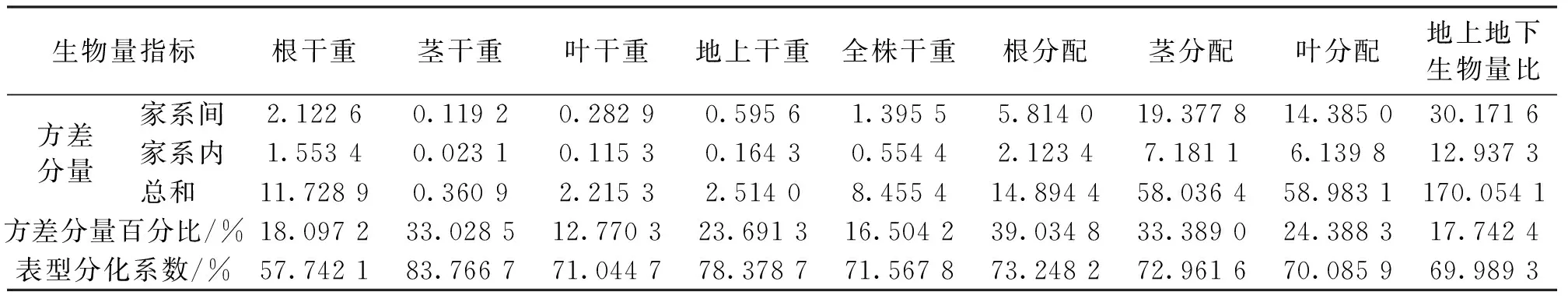
表3 2 a生云南松各家系生物量的方差分量与表型分化系数Tab.3 Variance components and differentiation coefficients of phenotypic traits for biomass among families of two-year P.yunnanensis
2.4 2 a生实生苗生物量变异特征
如表4所示,2 a生各构件器官生物量累积量的平均CV为39.70%~55.40%。其中,根干重变异最为丰富,11号家系根干重的变异程度最小,16号家系的变异程度最大;叶干重的分化较小,CV为39.70%,其2号家系的变异程度最小,9号家系的变异程度最大。

表4 2 a生云南松各家系生物量的变异系数Tab.4 Coefficient of variations for biomass among families of two-year P.yunnanensis %
构件器官生物量分配比例和地上地下生物量比的平均CV依次为39.12%、11.25%、7.78%和44.36%。根分配比例的变异程度最低,12号家系根分配比例的CV最小(4.52%),4号家系最大(22.14%);地上地下生物量比的变异最为丰富,其19号家系的CV最小(18.07%),2号家系CV最大(48.17%)。
2.5 2 a生实生苗生物量遗传特性
如表5所示,2 a生云南松苗木各构件器官生物量累积量的遗传力在0.39~0.45的范围,遗传力都略小于0.50,遗传力低于中等水平,表明变异受遗传因素的影响水平较低,其中根干重的遗传力最高,叶干重最低。

表5 2 a生云南松生物量的遗传参数估算Tab.5 Estimates of genetic parameters for biomass of two-year seedling
生物量分配比例和地上地下生物量比的家系遗传力都较生物量累积指标的遗传力高,在0.46~0.62范围,仅茎分配比例的遗传力低于中等水平,其余指标的变异受遗传因素的控制力较大,叶分配的家系遗传力最高(0.62)。
如果通过人工选择,各构件器官的累积与分配能在不同程度获得遗传增益。选择地上地下生物量比和根干重可以获得的增益较高,分别为14.21%和10.26%;选择根分配能获得的增益最小,仅为5.51%。
3 结论与讨论
2 a生云南松实生苗各构件器官生物量累积与生产力的关系十分明确,即构件器官生物量累积越多,苗木的总生物量累积就越多,生产力也就越大。但总生物量大小与不同构件器官生物量分配比例有着各种微妙的关系。不同构件器官间生物量分配比例的不同会影响各构件器官生物量的累积,也会一定程度影响苗木的形态特征,最终影响苗木的生产力[16]。以往学者也在群落、个体等水平十分关注有关植物形态指标的变异规律,在草本植物、针叶树种的形态指标调控和变异规律等方面开展相关研究[17-19]。作为可以描述植物形态的指标之一,地上地下生物量比可用于对苗木质量、林木生长特点、碳循环分配等问题进行探讨,因此了解这类性状在家系间的变异规律十分有必要。本研究发现绝大部分与苗木生产力和生态适应能力紧密相关的指标,在家系间存在具有统计学意义的变异现象,且家系间变异占主导地位。根生物量、茎生物量、叶生物量、地上生物量、总生物量、根分配比例、茎分配比例、叶分配比例、地上地下生物量比等9个生物量累积与分配指标经多重比较分析,均可以在P<0.05的显著水平分为2组以上,具备家系间选育的差异基础。所有指标中,生物量大小有关的指标及地上地下生物量比指标在家系间的变异较为丰富。但生物量大小有关的指标遗传力较小且普遍低于中等水平,生物量分配比例有关的指标和地上地下生物量比遗传力高于中等水平,将地上地下生物量比作为选育指标的遗传增益最高。本研究发现生产力较高的苗木,地上地下生物量比处于较低水平。同时有研究表明,茎根比、高径比等一些与苗木形态相关的参数,可以作为体现苗木活力水平的指标,如茎根比小的苗木活力水平高,造林后生长表现也较为良好[20]。因此,如以2 a生云南松苗木为材料开展早期选育时,宜选择地上地下生物量比小、生物量较大的家系。
李莲芳等[6]用不同苗龄云南松苗木进行造林效果的研究,认为苗龄较大、高径生长量更大并解除“蹲苗”的苗木,有助于解决造林后一段时间高生长弱、竞争力低的问题,而且苗龄较大苗木的生长性状分化程度也较高,提供更好的选育基础。通过对比其他研究中云南松不同苗龄苗木的生物量情况,发现随着苗木生长,最合理的生物量累积与分配指标可能有所变动。例如陈诗等[21]的研究中,育苗第1年由于苗木尚小,个体间地上部分对空间的竞争较小,苗木个体和家系将主要在争夺水肥等生态限制因子的能力上产生更多的分化。而随着苗龄和密度的增大,2、3 a生苗木为获取生存资源,对地上部分的分配比例要比1 a生苗木更大,随之将带来对光资源竞争能力的强烈分化[22-23]。因此,建议继续深入地摸清生物量各指标在时间上的遗传变异规律,以便科学筛选不同苗龄苗木的选育指标,确定部分优良性状的最佳考察时间[24]。
本研究中2 a生云南松苗根系生物量分配比例均低于蔡年辉等[25]、汪梦婷等[26]研究中2 a生苗木,而地上部分分配比例则较其他报道大。这可能与苗木种植密度、光水肥等管理措施有关。苗木的地上部分负责吸收太阳能进行光合作用,地下部分吸收水分和养分,两个构件器官共同作用了植物与外界的交流和平衡,生物量的大小与比例变化,是植物形态对生态因子的可塑性具体体现[27]。本研究的播种密度为5 cm×5 cm,小于其他研究的5 cm×10 cm,因此,可能分配更多的生物量用于对地上空间的争夺。
作为西南地区重要的阳性先锋树种,云南松的天然更新是森林演替和可持续管理的重要环节,本研究初步探讨了苗木生物量在田间管理情况下家系间的变异规律。继续挖掘自然状态下云南松幼苗、幼树的个体或群体生物量分配等生理生态规律,或能为该树种的森林经营、苗木生产和遗传改造提供更加丰富的基础参考[28-32]。
