亚洲金猫(Catopuma temminckii Vigors and Horsfield)的研究进展及威胁分析
2021-01-05刘健瑜黄建肖凌云冯杰
刘健瑜, 黄建, 肖凌云, 冯杰*
1. 北京市海淀区山水自然保护中心,北京 100871;
2. 北京大学生命科学学院自然保护与社会发展研究中心,北京 100871
全球37 个现存野生猫科物种中,已有36 个被列为稀有物种[1]。中国分布着13 种猫科动物,是世界上猫科动物多样性最高的国家[2]。全球对于野生猫科的调查研究主要集中在豹亚科如老虎Panthera tigris和雪豹Panthera uncia等大型猫科动物,而中小型猫科动物如云豹Neofelis nebulosa、亚洲金猫Catopuma temminckii和 云 猫Pardofelis marmorata的关注度相对较少。中小型的猫亚科动物在森林系统扮演着控制小型哺乳类和鸟类数量的重要角色,但是对于它们分布和生态学的了解甚少,基线生态数据的欠缺,以致保护状况未明[3]。
亚洲金猫(以下简称金猫),学名Catopumatemminckii, 异 名Felis temminckii和Paradofelis temmickii。英文名 Asiatic Golden Cat, Golden Cat,Temminck’s Cat[4]。金猫在中国古语称为“彪”。中国对各地和对不同毛色的金猫有不同的俗称,如芝麻豹、乌云豹、红椿豹和黄虎等[5]。金猫为中小型猫科动物的代表性物种之一,体长75~100 cm,尾长35~60 cm,体重8~16 kg[2],是除云豹外中小型猫科中体型最大的[3]。全球对金猫的研究主要集中在东南亚和南亚地区的种群[3],而中国金猫种群的研究较少,主要作为大熊猫伴生动物的调查纳入其中[6,7]。虽然缺乏可靠的金猫种群评估,但根据已有研究可推测出金猫的种群数量在持续下降中[4]。本研究综述了全球的金猫研究现状,分析了金猫种群的分布和数量变化趋势,并分析了金猫面临的威胁因素,为金猫保护、监测和研究提供建议。
本文基于1984 至2019 年间发表,以金猫为研究对象的生态学和保护生物学中英文文献,综述金猫的研究状况。
在谷歌学术搜索引擎中(Google)使用关键词“Asiatic Golden Cat”“Catopuma temminckii”。同时,在中国知网搜索引擎中(http://www.cnki.net),使用关键词“亚洲金猫”“金猫”(“Catopuma temminckii”“Asiatic Golden Cat”),来 分 别 识 别相关中英文引文。未囊括未发表论文和会议论文。本次共识别出了64 篇从1984 年至2019 年发表的且明确关注亚洲金猫的文献。
1 分布与生境
1.1 分布情况
金猫分布于中国华南、西南地区,向西延伸至喜马拉雅南麓,向南延伸至东南亚。分布国家包括中国、越南、老挝、柬埔寨、泰国、马来西亚、印度尼西亚、缅甸、印度、孟加拉国、不丹与尼泊尔[4]。金猫分布范围虽然广泛,但在印度东北部至孟加拉和尼泊尔东呈稀少零星散落的分布[8-11],在柬埔寨、老挝、越南和中国南部的记录较少[4,12-14],而广泛分布于不丹、缅甸、泰国和马来西亚[4,15]。除了印度尼西亚群岛以外,苏门答腊半岛也有广泛的记录[4,16-18]。
在国内,金猫的历史分布区曾广布全国14 省。如今金猫在国内的分布呈孤岛状和高度破碎化[19],广东、江西、福建、浙江、湖南、湖北、安徽等地区已基本没有金猫的野外记录。近期仍有记录的地区是四川北部和西部、甘肃东南部、宁夏西南部、陕西西南部、西藏东南部、云南西部和南部[19-22]。
1.2 生境
金猫分布状况共搜集到17 篇,其中包括全球分布情况(n=15)和国内分布状况(n=2);金猫生境相关文献共搜集5 篇。
金猫主要分布在热带和亚热带常绿林、混合常绿林和干燥落叶林的森林生境[4],栖息地的选择还包括高海拔的以冷杉为主的针叶林、阔叶混合针叶林和草甸等[3]。开阔的环境,如灌丛和草地,或者是裸露的岩石区,乃至退化或不连续的森林景观都有分布记录。金猫的海拔分布跨度大,最高海拔记录于不丹达4 282 m[23],但总体上还是更偏好低山森林[4]。在苏门答腊半岛,发现一只佩戴无线项圈的金猫,常跑到保护区外一片被种植的咖啡豆而破碎化的林地里[16]。而在泰国有更多出现在密闭林地的记录[24]。可见,金猫的栖息地并不单一固定。
2 种群动态
金猫种群状况搜索到的文献共28 篇,其中有金猫种群总体描述(n=1),皮毛调查反映的部分野外种群(n=4),国内红外相机监测反映金猫野外种群状况文献(n=)3 篇,金猫与其他同分布野生猫科动物的相对丰富度文献共12 篇,以及人工种群状况(n=8)。
2.1 野外种群动态
目前仍没有金猫野外种群的评估,种群密度和种群数据匮乏,难以准确评估该物种真实的种群状况。但根据现有数据指出,近几年大量的栖息地丧失和盗猎威胁,金猫的整体种群数量呈现20%~30%的下降幅度,而且这个下降趋势很有可能会持续,尤其在中国和越南,其种群数量持续急剧下降,金猫在中国和越南的大部分地区可能已经在面临局部灭绝[4]。
根据1995—2013 年越南通过红外相机和市场调查可知,金猫的种群数量呈下降趋势,推测种群数量减少的主要原因是盗猎[25]。中国浙江金华的哺乳动物资源调查可知,20 世纪60—70 年代以金猫为主的皮毛产量稀少且呈下降趋势[26],广西也呈相似的变化,踏入20 世纪80 年代后金猫皮毛更难发现[27],由于豹Panthera pardus已近灭绝,金猫的皮毛需求量增加,后者的猎杀随之更为突出。然而,在中国江西的调查中显示出不一样的趋势[28],金猫被捕量从20 世纪50 年代中到80 年代的140 头,增加到1980 年的240 头,数量增长明显。由此可见,由毛皮调查反映出的金猫野外种群的历史变化趋势,呈地区差异。
近年来,在秦岭、岷山和邛崃山脉内开展了不少利用红外相机的调查。通过红外相机的拍摄率——单位时间内物种被红外相机捕捉到物种的频率(拍摄率(%)=有效照片数/相机有效工作日×100)可以反映区域内金猫的种群情况,在位于陕西的长青保护区华阳保护站内,开展2009—2010 年对金猫的种群密度研究,利用红外相机监测,得到金猫在长青保护区的红外相机拍摄率为0.43%,研究者计算出当地金猫的种群密度是7.2~10.8 只·100 km−2[29]。在108 国道秦岭生物走廊,2009—2010 年间共布设红外相机45 架次,得到金猫的样线遇见次数5 次[7]。2002—2009 年间对大熊猫栖息地的五大山脉11 个自然保护区的猫科动物进行调查,仅有秦岭长青保护区和岷山唐家河保护区有金猫的记录,其拍摄率分别为0.7%和0.2%[6]。在岷山老河沟自然保护区,研究人员在2011—2014 年间共设置125 个调查点位,70 个1 km × km 有效调查方格,有效探测金猫34 次。可见,中国已发表的金猫野生种群相关的调查集中于秦岭和岷山地区。
另外,在岷山北部的老河沟、唐家河以及甘肃的白水江保护区 3 个地区都有连续的金猫记录,李晟指出中国西南部(秦岭、岷山、甘孜、藏东南等)有较稳定和一定规模的金猫分布(李晟通信确认)。
虽然金猫的整体数量在下降,但有研究表明,金猫比同区域分布的其他野生猫科动物(云豹、马来云豹Neofelis diardi和云猫)的记录更多[4,8,25,30-33]。在不丹Jigme Dorji 自然保护地对6 种猫科动物—金猫、老虎、豹、云豹、云猫和豹猫Prionailurus bengalensis种进行相对丰富度(Relative Abundence Index, RAI)的分析,该研究的RAI 是每100 天拍摄到的独立照片数,其中金猫的相对丰富度最高(RAI=0.8)[34]。然而,也有金猫相对少见的记录[4,35,36],Yongdrup[37]发现不丹的Jomotsangkha 野生动物保护区,在一定强度的森林砍伐和人为干扰栖息地中,根据金猫红外相机拍摄到的记录,其相对丰富度(RAI=0.17)不如云豹(RAI=0.68),可见,金猫与同分布的其他猫科动物的相对丰富程度因地区而异,人为干扰对金猫及其他野生猫科动物的相对丰富度的影响,有待进一步研究。
2.2 人工种群动态
由于工授精成功率极低和人工繁殖的死亡率高,全球动物园圈养的金猫情况不容乐观[38-40]。根据《国际动物园年鉴》的统计,欧洲、亚洲、澳洲、北美洲的动物园都曾尝试建立稳定的金猫人工种群,但饲养金猫的动物园逐年减少,截止到2013 年的统计数据可知,在北美洲和澳洲动物园中已无金猫人工种群,仅有51 只金猫饲养于欧洲和亚洲的动物园[41]。金猫全球人工种群数少的另一个原因是,国际动物园界按照3 个亚种的理论来管理,彼此不能杂交。欧洲动物园维持着两个亚种的金猫种群—西藏亚种(C. t. tristis)和指名亚种(C. t.temminckii),近亲繁殖的情况严重,迫切需要亚洲人工种群的加入(D.B.Sandra Reichler, 2018)。2018 年最后的1 只人工饲养的金猫华南亚种(C. t.dominicanorum)在杭州动物园死去后,亚洲现存的人工种群也仅维持到两个亚种。
金猫的人工繁殖成功率低,表现为一胎仅产1 崽,少数产2 崽[27],且有雌性产崽后情绪不稳定或只关注1 崽,幼崽有不少人工哺育的记录[40,42]。另外,圈养条件下,雄雌金猫配对时极易产生暴力行为,甚至有一方致死的记录[38]。可见,金猫人工繁育难度大,建立和维持稳定的金猫人工种群难度大。
3 食性和活动节律
3.1 食性
金猫食性相关的文献共搜集到6 篇文献。金猫主要捕食小型的脊椎动物,如鸟、蜥蜴、蛇、啮齿类,大到如小麂Muntiacus reevesi、毛冠鹿Elaphodus cephalophus等小型有蹄类食草动物,在老河沟通过分子生物学技术发现金猫还会捕食体型较大的野猪Sus scrofa、羚牛Budorcas taxicolor等大型有蹄类动物幼体[43],在马来西亚捕食最大的猎物为郁乌叶猴Trachypithecus obscurus,在印度还有猎杀水牛的记录[44]。金猫白天和夜间都有活动记录,表明其猎物可能是昼行性和夜行性动物的组合[24,31,44]。金猫的食物网络结构有待未来进一步研究。
3.2 活动节律
金猫日活动节律的相关文献共搜索到11 篇,季节性活动节律的文献有3 篇。
3.2.1 日活动节律
金猫日平均活动距离1 597 m/d(Grassmanet al.,2005),在中国藏东南、马来半岛、缅甸、泰国和印度的研究发现,金猫偏昼行性或黄昏性(crepuscular),活动高峰为黎明、中午和黄昏,半夜期间也有小活动高峰[4,32,34,45-48]。2013—2018 年在藏东南部开展红外相机调查,发现金猫的活跃期为10:00—24:00[2]。然而,也有金猫昼伏夜出的记载[27](吴名川,1986),在秦岭长青保护区[49]监测到的金猫是夜行性的(晚上拍摄率80%)。有泰国和不丹的研究指出,金猫的日活动节律与人类活动有关,栖息地非原始森林或人为干扰大的林地,金猫则更偏向夜间活动[46,50],在保护地与有一定人类活动强度的非保护地的金猫,其日活动节律无明显差异,记录到的金猫皆为白天活动更为显著。
金猫曾经一度被认为是夜行性动物,越来越多的研究得到金猫的日活动节律是偏昼行性的,期间至少有两个活动高峰,但也有秦岭地区和人为干扰强度较大的地区,金猫显示出夜行性活动的记录,昼夜活动节律的影响的因素有待进一步研究。
3.2.2 季节性活动节律
金猫呈现不同程度的季节性迁移,冬春季常分布于海拔较低的林地,而夏秋季则会上迁到海拔较高的区域[2,49]。迁移可能与猎物的迁徙、生理及降雨有关[49]。不丹Jigme Dorji NP,虽然在秋季和冬季的红外相机捕捉率无明显差异,但金猫在秋季的空间覆盖度高于冬季[34]。然而,金猫在藏东南金猫的活动高峰期—不同季节(月份)的活动强度与红外相机的拍摄率呈正相关,拍摄率越高,该物种活动强度也越高,表明其在此时间段内更活跃—集中在2 月、5 月、7 月和10—11 月,对应可能是该物种的繁殖周期和雨季[2]。
4 色型研究
金猫以其不同色型的变异在猫科动物中显得尤其突出,常见的有红棕色型、麻褐色、灰色型和黑色型,以及罕见的花斑色型。根据已有研究发现,红棕色和灰色型的金猫较广泛分布于亚热带的栖息地。花斑色型在干燥落叶林、热带草原、草地,甚至灌木地更为常见,主要分布华东、四川、喜马拉雅山脉东南-高黎贡山、缅甸北部。黑色型的金猫则更容易发现于热带和亚热带森林,印度东北边的温带亚高山和高海拔栖息地的藏东南和横断山脉、喜马拉雅山脉东段南坡[2,4]。各地的色型比例差异较大,总体变化趋势呈克拉地峡南北部及过渡:中国南部有最多样的4 种色型且比例均衡,往巽他群岛方向的种群,花斑色型和黑色型逐渐减少,到黑色型极少和花斑色型消失,只剩红色和灰色2 种色型。但其色型分布、形成机制、不同色型在不同森林类型的分布,仍有待进一步研究[51]。
有研究提出过不同色型与特定海拔分布有关[3,22],但不丹也有监测到多种色型出现在同一海拔跨度内[47]。通过红外相机发现,不同色型的金猫,有混居的情况出现,不丹监测到4 种色型金猫的栖息地重叠,甚至还发现两种不同色型的金猫出现玩耍性较量的社交互动[47]。
IUCN 根据金猫的体型和毛色将其分为3 个亚种,分别是指名亚种(C. t. temminckii),通体红棕色或灰棕色,胁部没有明显的斑点和条纹,体型最小最瘦,分布于苏门答腊、马来半岛、中南半岛、滇南、缅甸直达喜马拉雅山南麓;华南亚种(C. t.dominicanorum),毛色和体型介于指名亚种和西藏亚种之间,分布于中国东南部;西藏亚种(C. t.tristis),则是体型最大,毛厚,胁部有稀疏的蛛网状白色细纹,颈下、胸口、腿部有黑色斑点,体色以灰色调为主,但越往背部越偏红,而且常出现花斑色型,其分布于青藏高原东南缘的高海拔地区,包括西藏东部、青海南部、四川、甘肃、陕西、重庆、滇西北和缅甸北部等[2,4]。
然而,其他研究则表明,多样的金猫色型与3 个亚种没有明显关系,有人质疑依据体色来划分亚种的方法。期间有研究者提出云南的金猫应独立成另一亚种C. t. bainesiSowerby, 1924;而尼泊尔、中国西藏南部、云南西北部和四川西部的金猫则列为C. t. moormensisHodgson, 1831。在2016 年,Riddhi P. Patel[51]在结合了分子生物学、古地理学和形态学的研究,提出金猫只是分2 个亚种的理论,克拉地峡南部的苏门答腊和马来半岛种群C. t. temminckii,以及克拉地峡北部亚洲大陆种群C. t. moormensis。此理论能部分解释金猫南北部种群体型的差异和整体色型比例变化趋势,并指出色型比例的减少,可能是金猫巽他种群为适应当地差异更大的栖息地环境而经过长时间的选择结果。
搜索到金猫色型主要的生境类型、海拔分布和地理位置变化趋势文献6 篇,国内色型分布的文献1 篇,以色型与亚种分类的讨论有4 篇文献。
5 金猫与其他猫科动物的关系
5.1 金猫在猫科系统发育的位置
金猫在猫科系统发育位置的文献搜集到1 篇文献。王金凤等[1]对猫科物种之间的系统发育关系从体型、毛皮斑纹模式的变化、骨骼和软组织等的形态学特征,染色体数的细胞学,白蛋白免疫距离、蛋白电泳、同工酶、化学信号的、线粒体基因、核基因的分子生物学方面进行概述,由于不同学者在选材, 研究方法, 依据理论和研究目的等方面各不相同,其研究结果会有差异,也有达成一定共识的:37 种猫科动物至少可以分成3 个亚科(Subfamilies)或谱系(Lineages):家猫谱系、豹谱系和虎猫谱系,以及8 个谱系:虎猫谱系、家猫谱系、豹谱系、美洲狮谱系、猞猁谱系、豹猫谱系、狞猫谱系和金猫谱系。其中亚洲金猫属于3 谱系分类方法的豹谱系,以及8 谱系分类方法的金猫谱系(见表1)。
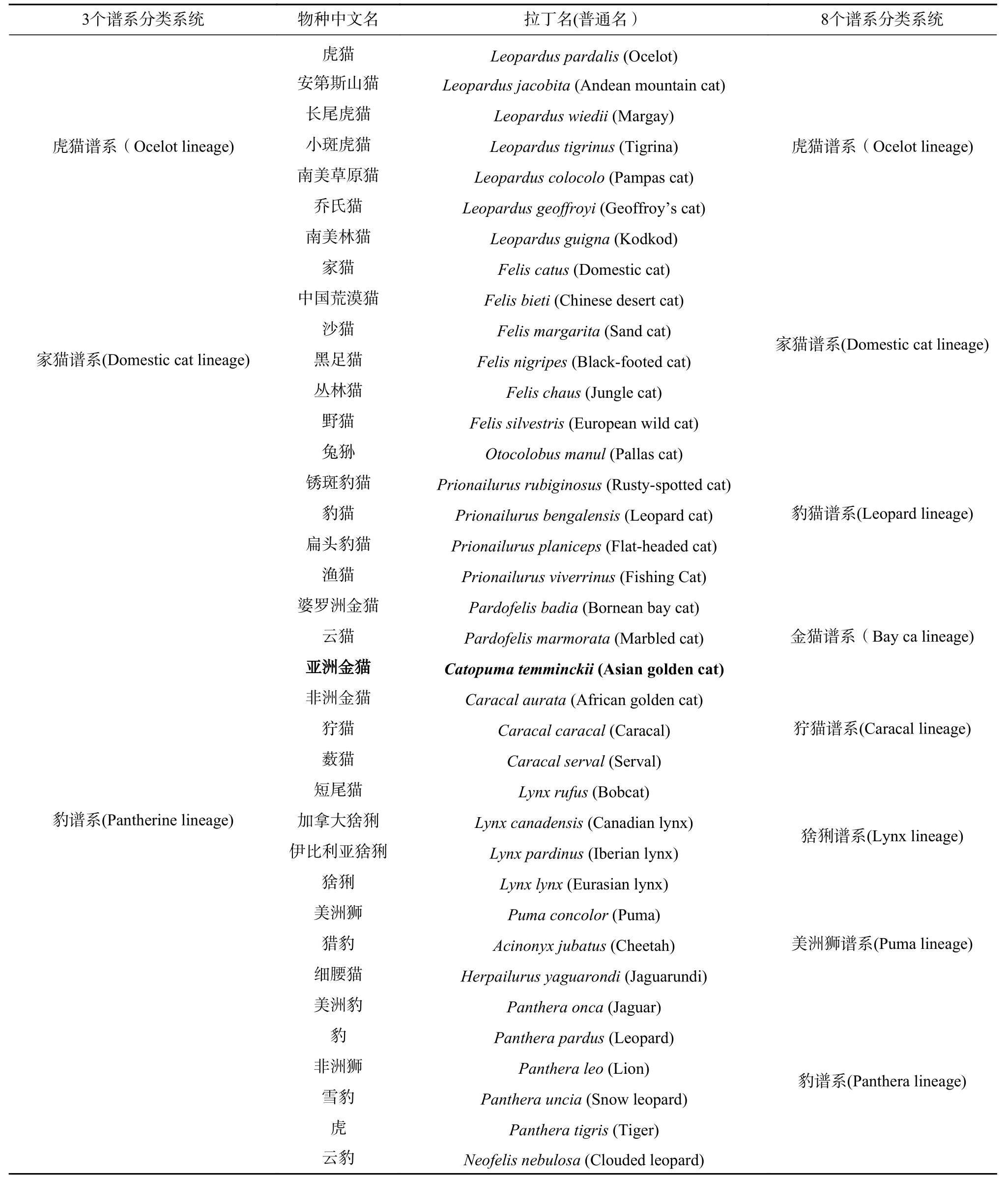
表 1 现存猫科 37 个物种及分类系统Tab. 1 37 species of existing Felidae and taxonomic classification system of existing cats
5.2 金猫与其他猫科动物生态位分化
金猫与其他同分布的猫科动物的日活动节律分离的文献收集有5 篇,其中有金猫日活动节律与其他同分布猫科动物的重叠度分析(n=5),讨论了不丹JDNP 的金猫与老虎、豹、云豹、豹猫和云豹的季节性活动范围、空间、栖息地类型和海拔分离(n=1),另外,在老河沟也有对金猫和豹猫进行食性的对比讨论(n=1)。
5.2.1 日活动节律分离
很多学者通过对比不同猫科动物之间的日活动节律以及日活动节律重合度来讨论它们之间的日活动节律分化程度。同分布猫科动物日活动节律重合度(以下简称“重合度”)通过使用统计学方法的核密度估计Kernel Density Estimation(KDE)[17]或拟合三角函数(trigonometric sum)分布[52],分别计算同分布猫科动物的日活动节律,然后得出两个焦点物种之间的分布情况[17]。偏向使用重叠系数△,当△=1,显示出活动密度,而△=0,没有共同活动时期。
东南亚对金猫与其他猫科动物的活动节律重合度研究发现,金猫、老虎及豹的活动节律高度重叠[17,50],可能会增加金猫被老虎和豹捕食的概率。由于在东南亚分布的金猫、云豹、豹猫和云猫栖息地类型相似,4 个物种分布区常见于同一片区域[53],而其中与金猫体型相近的云豹,两者却只显示出小程度的日活动节律重叠,不显示明显的种间竞争,可能是云豹更长的身体比例,较金猫捕食的猎物要大;金猫与云猫的重叠度也低[17,24],然而在印度分布的金猫与云猫显示出较高的重叠度[45]。但在中强度的人为活动干扰下,金猫、云豹和云猫活动节律的重叠度增加,而金猫与豹的重叠度则有所下降[17, 37, 50]。
5.2.2 季节性活动空间利用、栖息地类型和海拔分离
同时Thinley[34]提到在不丹的Jigme Dorji 国家公园(JDNP),金猫与老虎、豹、豹猫分布于同一片栖息地,金猫在秋季活动范围是冬季的2 倍,豹的活动范围也呈现类似的秋季大于冬季,而老虎相反,冬季的活动范围更广,还有豹猫在冬季和秋季的活动范围则无明显差异。
JDNP 主要有7 种栖息地类型,金猫、云豹、豹和云豹仅发现于阔叶林和混合针叶林中,老虎则主要栖息于混合针叶林。
JDNP 研究区域的最高海拔4 105 m 有老虎和豹猫的记录,而金猫、豹、云豹和云猫被记录于的该区域内的最高海拔为3 488~3 810 m。
5.2.3 食性分离
在对四川省老河沟自然保护区中的金猫与豹猫的食性分析文献中[54],提到金猫与豹猫食物组成中,主要的都是啮齿类和鼠兔类,但是前者的食物宽度比后者要广,且在金猫的食物组成比例中,体型较大的猎物也比豹猫的要大,有利于降低种间竞争。
可见,金猫与其他同分布的猫科动物之间,生态位分化程度呈现地域的差异。而在人为干扰下,金猫与云豹和云猫的种间竞争有待进一步研究。
5.3 毛发对比
老虎、豹和金猫毛发的显微镜特征研究发现,金猫与老虎和豹的髓质指数差异大,金猫头、颈上、胸、腹、前肢等5 个部位的毛髓质指数区间为50.0%~70.3%,老虎的毛髓质指数区间为22.5%~50.0%,豹的毛髓质指数区间为27.3%~49.3%,可以通过计算髓质指数将金猫和老虎及豹的毛发区别[55]。
有学者发现金猫和豹猫的针毛在傅立叶红外光谱(FT-IR)有区别,可以用于毛发无损的种间识别。两者针毛中部二阶导数谱均具有明显的种间差异,在648 cm−1和654 cm−1波数处金猫出现明显M 形双峰吸收峰,而豹猫没有;金猫的二阶导数谱在709~763 cm−1范围内以峰强较大的单峰为主,豹猫则以峰强较小的M 形双峰为主,这均能说明两种动物毛发的蛋白侧链的构成上则存在较大的差异,同时也存在种间的相似性[56]。
6 威胁
金猫目前发现的威胁主要分为两类,分别是疾病及寄生虫的影响和人为直接和间接干扰的影响。
6.1 寄生虫、细菌及病毒情况
关于金猫疾病的文献共搜集到7 篇,其中有寄生虫(n=5)、细菌(n=1)和病毒(n=1)。对金猫寄生虫的研究主要来源于动物园人工的金猫个体,目前为止,发现有6 种寄生虫寄生于金猫的不同部位和器官。有发现于金猫的腹部和背部的猫虱子Ctenocephalides felis felis记录[57],常见于家猫及家畜的。通过对金猫的驱虫和尸检记录发现:肺部—卫氏并殖吸虫,胃部—有鞘泡翼线虫,十二指肠—犬弓首蛔虫、曼氏裂头绦虫,空肠—狮弓蛔虫,心室—心丝虫。其中较为严重的是鞘泡翼线虫,能引起严重的炎症和器官内出血,以及苏州动物园有心丝虫病致死的情况[58-60]。宁波记录到一只感染多种寄生虫的野生金猫非常健康,提出寄生虫对野生金猫健康状况的影响较圈养金猫小[61]。
另外,犬瘟热是能通过犬瘟热病毒能感染狐狸、家狗等犬科动物,以及包括金猫、老虎、豹和狮等猫科动物,并引起大量死亡的疾病[62]。
放线菌病,由放线菌(Actinomyces)感染引起金猫脓毒败血症和胰腺、甲状腺等病变的病例[63]。
6.2 人类活动影响
人类活动中导致的生境破坏和破碎化,以及非法盗猎是金猫种群急剧下降的重要因素。其中工程建设,毁林(包括农用地开拓、放牧及薪柴采集等)和砍伐活动等造成金猫分布区的栖息地破坏和丧失,对金猫种群密度的影响至关重要[24,37]。
有研究指出每个有金猫分布的国家均存在不同程度的盗猎情况,中国和越南的盗猎压力非常大[4],尤其是越南,导致了金猫在越南几乎灭绝。在越南2 个国家公园内和边缘区就记录到500 个陷阱[25],这些都是“无选择性”的绳套陷阱,其设计和放置的位置对像金猫这样的中小型猫科动物是致命的,其次,这些陷阱对金猫的潜在猎物的种群数量影响也非常大,其中的猕猴、野猪、鼷鹿科、野鸡常见在野生动物贸易中。
发现关于金猫贸易的论文有6 篇。金猫在非法贸易中主要以活体、冷藏的尸体、毛皮、骨骼和药酒的形式出售[25,27,64],其骨骼在中药店被称为“豹骨”的药材[28]。金猫是非法贸易调查中最常见到的物种之一,在缅甸1991—2013 年的12 次野生动物贸易调查中,就有发现10 次售卖金猫的情况[65]。随着老虎和豹的野外种群变少,金猫的猎杀情况变得更为突出,不时发现被冒充老虎皮和身体部位的,以及豹皮的替代品[4,25,26,65]。缅甸-泰国边界、苏门答腊和印度东北部部分地区也有兽皮交易的记录[4]。1997 年对越南-云南边界城市的非法贸易调查,记录到金猫都是从中国云南境内出售,有2~5 次记录[64]。
7 保护行动及其研究需求
金猫是中国国家二级保护动物,在IUCN 红色名录被列为近危物种(NT),以及被列进CITES 的附录I 中,金猫在大部分有分布的国家都受到法律保护。但是,大多数中小型猫科动物,很少成为主流野生动物保护的焦点,它们种群密度低,监测难度大的原因,极少有针对它们的野外调查,为将来更好支持和推进金猫的保护和研究工作,提出以下几点建议:
(1)加强关注和研究。除了种群数量和分布的研究,对于金猫的空间和时间活动规律也需要进一步研究。金猫分布范围广,各个不同分布地区的空间和时间活动规律不完全相同。作为森林生态系统中的残存的顶级捕食者之一,金猫有着重要的生态价值。但现阶段金猫还没有得到相应的关注度。建议在未来宣传和研究当中,将金猫作为重要物种之一,提高金猫的关注度,吸引更多研究者进行研究。
(2)采取相应的保护措施。金猫的生境破碎化严重、野外种群呈现出下降趋势,保护行动迫在眉睫。在我国,还没有专门的金猫自然保护区,在国外,金猫也一直存在严重的人为干扰。所以,要在金猫分布的地区调查金猫受到的威胁,建立专门的保护地;已发现辖区内有金猫存在的自然保护区应该制定专项的金猫保护行动;恢复栖息地,建立生态廊道,缓解金猫生境的破碎化。最终,防止金猫种群数量的进一步下降。
(3)利用新技术对金猫野外种群进行研究。在野外,因为金猫独居、有些区域的夜行性等习性,导致了金猫的野外数据收集很困难,进而导致了金猫的野外种群评估工作不好开展。可参考其他食肉动物的研究技术和方法,非损伤方法如空间标记重捕法等,为金猫的研究和保护创造新的机遇,对其的生态学发展具有重要的意义。基于这些方法,应尽快制定系统性大尺度区域调查的标准,确保目标区域广泛涵盖金猫各地的分布区,有助于识别关键区域,开展金猫密度和动态等更深入和长期的研究。
(4)加强金猫的病理研究,包括对该物种有潜在威胁的寄生虫、病毒和细菌对人工种群和野外种群的了解。另外,对金猫有致死性的犬瘟热病毒,可以通过家养犬传播感染,而村庄的家养犬一般没有得到规范管理,能够自由进出保护区或有金猫分布的分布区域,可能会对金猫构成潜在威胁,因此可以开展犬瘟热对金猫影响程度的调查,以及制定相应的防治措施。
(5)加强金猫盗猎和非法贸易的打击力度,对有金猫分布的保护区,加强反盗猎专项巡护,在保护区以外的金猫分布区可以建立保护地实现有效管理;其次,确立清楚金猫的动物制品,无论是尸体、装饰品还是药材等都属于违禁品,都要得到严格管理;而对于非法贸易,可以从法律法规、执法、执法人员管理、销售渠道、运输方式、公众保护意识等来有效结合,减少甚至消除盗猎和非法贸易对金猫野外种群的威胁。
