腺苷及腺苷A1受体在小鼠肾小球系膜细胞收缩中的作用
2020-12-15姜世敏许世清高红梅李文歌
刘 琳,姜世敏,卓 莉,徐 波,许世清,向 青,高红梅,李文歌★
(1.中日友好医院 肾病科,北京 100029;2.中日友好医院 临床研究所,北京 100029)
腺苷和腺苷A1 受体(adenosine A1 receptor,A1AR)介导了管球反馈(tubular golmerular feedback,TGF),A1AR 的阻断能够消除TGF。但近年来进一步的研究显示,选择性阻断入球小动脉平滑肌上的A1AR 仍有约36%的TGF 现象存在,提示肾小球其他细胞表达的A1AR 也在参与TGF调节[1]。系膜细胞作为肾小球毛细血管襻的支撑,被血管活性物质刺激后可发生收缩,参与调节肾小球滤过率,但其A1AR 表达情况以及A1AR 究竟是否参与系膜细胞收缩的调节,尚缺乏确切的证据。本研究在原代肾小球系膜细胞中观察A1AR 的表达情况,使用腺苷、A1AR 激动剂及抑制剂刺激系膜细胞,动态观察细胞收缩能力的变化,为系膜细胞在TGF、肾小球滤过率调节中的作用及机制提供更多依据。
1 材料和方法
1.1 材料
SPF 级野生型6~8 周雄性C57BL/6 小鼠购自北京维通利华公司。胎牛血清(fetal bovine serum,FBS)、2.5g/L 胰蛋白酶-EDTA、DMEM 培养基(含葡萄糖5.5mmol/L)购于GIBCO 公司。Ⅰ型胶原蛋白购于Corning 公司(354236),Ⅳ型胶原酶购于Gibco 公司(17104-19)。CCK-8 试剂盒购于上海东仁化学科技有限公司。抗小鼠desmin 抗体购于Proteintech 公司 (16520-1-AP),抗小鼠synaptopodin 抗体购于Progen 公司 (61094),抗小鼠A1AR 受体抗体购于Abcam 公 司(ab82477)。FITC 标记的山羊抗小鼠二抗、辣根标记的山羊抗小鼠二抗购于北京中杉金桥公司。血管紧张素Ⅱ(AngⅡ)、腺苷、N6-cyclohexyladenosine(CHA)、8-Cyclopentyl-1,3-dipropylxanthine (DPCPX) 购于Sigma 公司。
1.2 原代小鼠系膜细胞的提取、培养与鉴定
参照文献[2]中的分样筛法,断颈处死3 只C57BL/6 小鼠,于75%酒精浸泡1min。无菌条件下剖开腹部取出两侧肾脏,用冰冷的无菌PBS 清洗2 遍。剪下肾皮质,放入少量Ⅳ胶原酶中(1mg每个肾脏),将皮质剪成碎末,使用研钵一定程度的研磨后转移至50ml 离心管中,37℃水浴消化40min 后加适量含血清的培养基终止消化。收集肾组织混悬液体,用大量PBS 充分冲洗同时使其先后通过孔径为70μm 和40μm 筛网,使用无菌吸管收集40μm 筛网上的肾小球悬液,移入15ml离心管。以1000r/min 离心5min,弃上清,加入含20%FBS 的DMEM 培养基制成肾小球悬液,随后转移入1 型胶原蛋白包被过的无菌培养皿,放入37℃,5%CO2培养箱孵育。每3d 换液(3d 内避免移动培养皿),7~10d 后肾小球明显缩小甚至消失,爬出的细胞长满瓶皿后进行传代,前三代使用20%FBS 的DMEM 培养基,此后使用15%FBS 的DMEM 培养基。5 代后细胞形态一致性良好,肾小球完全消失,经免疫荧光染色Desmin(+)、synaptopodin(-)鉴定为系膜细胞,8 代以内的系膜细胞用于实验。
1.3 CCK-8 法检测腺苷刺激系膜细胞生长曲线
以2000 个细胞/100μl/孔铺96 孔板,每个浓度3 个复孔,置于37℃,5%CO2培养箱培养24h后吸出孔内培养液,加入100μl 无血清培养基和10μl CCK-8/孔,孵育2h 后于酶标仪检测OD 值,此点设置为0 点。向培养板加入10μl 不同浓度的腺 苷,使 其 分 别 达 到0.1μM、1μM、10μM 及100μM 浓度,继续培养,分别在24h、48h、72h 使用上述方法加入CCK-8 试剂进行检测,并与不加腺苷的对照组进行对比。
1.4 细胞免疫荧光染色
以1×104个细胞/孔铺24 孔板,待细胞生长至60%~80%融合时4%多聚甲醛固定液室温固定15min。PBS 漂洗,室温下用0.5%Triton/PBS 透化处理10min,3%BSA/PBS 37℃封闭30min。在细胞爬片上分别滴加约50μl 1:200 稀释的desmin抗体、synaptopodin 抗体或A1AR 抗体,密封于湿盒中,37℃孵育30min,避免震荡。漂洗后在细胞爬片上加约50μl 1:50 稀释的FITC 标记的驴抗兔荧光二抗,37℃孵育30min。漂洗后加入1:2000稀释的DAPI,室温孵育5min。漂洗后取10μl 封片剂滴在载玻片上,将细胞爬片有细胞的一面朝下封片,尽快在激光共聚焦显微镜下观察并照相。
1.5 Western blot
使用SDS 裂解液提取细胞总蛋白,每孔加入20μg 细胞总蛋白进行10%SDS-PAGEs 凝胶电泳分离,之后将蛋白转印至PVDF 膜,5%脱脂牛奶室温封闭1h,分别加入抗1:500 稀释的A1AR 抗体、1:1000 稀释的β-actin 抗体,4℃过夜。TBST洗涤后加入HRP 标记的羊抗兔IgG 二抗,室温孵育1h。洗膜后加入ECL 化学发光剂,于凝胶成像系统显影。
1.6 细胞收缩功能的检测
以5×104个细胞/孔将小鼠肾小球系膜细胞铺于直径为35mm 的免疫荧光专用培养皿中,待细胞贴壁后(约4h),将细胞在不同条件下培养。正常对照组使用普通DMEM 培养基培养;腺苷刺激组在DMEM 培养基中加入终浓度为100μmol/L的腺苷;A1AR 激动剂刺激组,DMEM 培养基中加入终浓度为1μmol/L 的A1AR 激动剂CHA;A1AR 抑制剂刺激组,DMEM 培养基中加入终浓度为1μmol/L 的A1AR 抑制剂DPCPX。培养48h后使用Olympus IX83 活细胞显微成像系统观察细胞收缩情况。将培养皿放入操作体系中,0 点拍照后原位更换为含1μM AngⅡ的培养基,关闭培养体系,维持温度37℃,5% CO2浓度的条件,在40 倍物镜同一视野下,连续观察30min,每2min拍照1 张,最后制作为动态影像。应用Image J 软件计算每个细胞在0 时和30min 时的面积变化绝对值,对比各组之间的差异。
1.7 统计学方法
应用SPSS23.0 统计软件,数据以χ±s 表示,先行单因素方差分析,方差齐性检验后,组间比较采用LSD-t 检验。
2 结果
2.1 原代系膜细胞的培养及鉴定
提取出的肾小球使用20%FBS 的DMEM 培养基在37℃,5%CO2条件下孵育3d 后,相差显微镜下观察可见肾小球内有细胞爬出(图1A,见封二)。10d 后肾小球明显缩小甚至消失,爬出的细胞可长满瓶皿(图1B,见封二)。第一次传代后每3d 传代1 次,传至第5 代,细胞一致性良好,肾小球完全消失。此时行Desmin 染色为阳性(图2,见封二),Synaptopodin 染色阴性,鉴定为系膜细胞。

图1 小鼠原代系膜细胞生长过程(×100)。A.提取小鼠肾小球并培养3d后细胞从肾小球内爬出;B.提取小鼠肾小球培养10d后细胞长满瓶皿。图2 小鼠系膜细胞desmin免疫荧光染色(×400)。图3 免疫荧光染色观察到A1AR于系膜细胞胞浆内呈颗粒状表达(×200),箭头所示为A1AR阳性区域。A.绿色为FITC标记阳性区域;B.蓝色为DAPI染色阳性区域。
2.2 腺苷对系膜细胞生长的影响
表1 示,使用CCK-8 试剂盒检测细胞生长,观察到0.1μM、1μM、10μM 及100μM 浓度的腺苷在24h、48h、72h 内,与不加腺苷的正常对照组相比,细胞数量均无明显差异,提示腺苷对小鼠原代系膜细胞的增殖无明显影响(均P>0.05)。
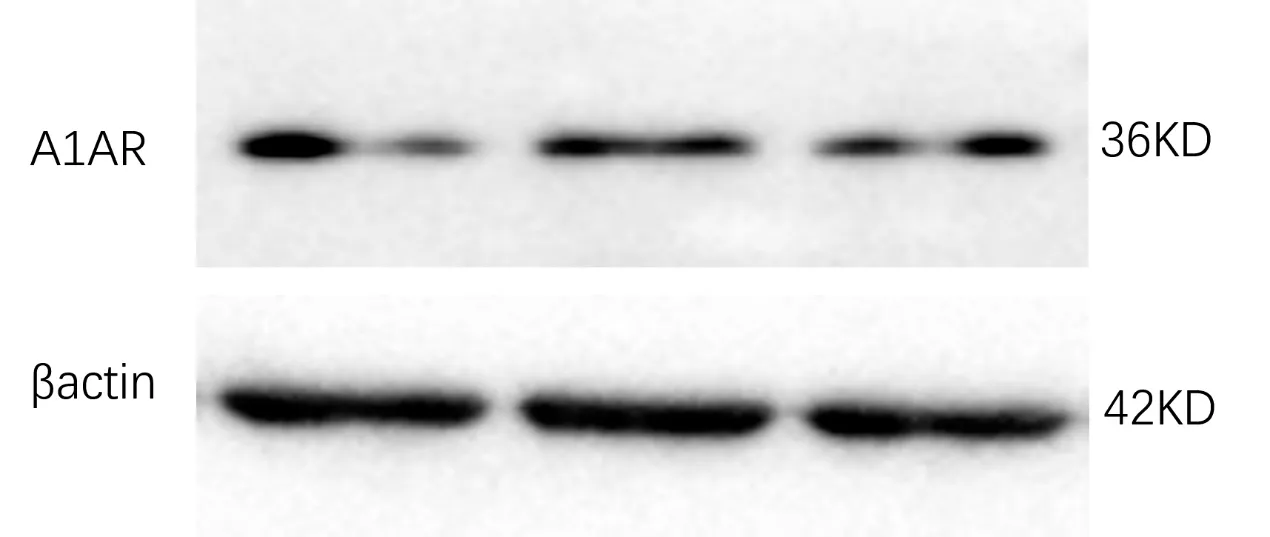
图4 Western blot 方法验证系膜细胞存在A1AR 表达

图6 AngⅡ刺激前后系膜细胞面积变化
2.3 小鼠系膜细胞A1AR 表达情况
细胞免疫荧光染色显示,A1AR 在小鼠肾小球系膜细胞中存在表达,并呈现细小颗粒状分布于胞浆中(图3,见封二)。同时,使用western blot技术也观察到原代小鼠肾小球系膜细胞能够表达A1AR,在36KD 水平可见深色条带(图4)。
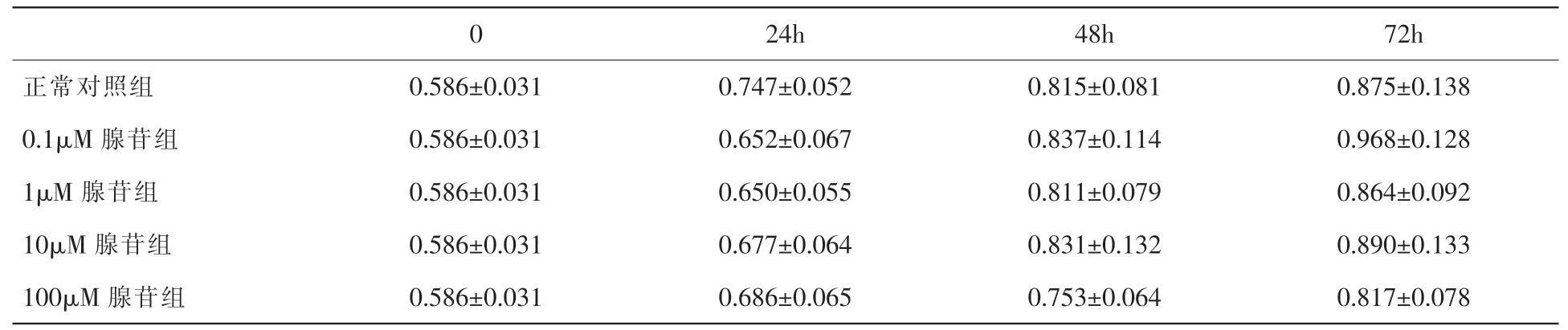
表1 不同浓度腺苷刺激系膜细胞在各时间点的OD 值
2.4 腺苷及A1AR 对系膜细胞收缩能力的影响
如图5(见封二)、图6 所示,100μmol/L 腺苷刺激48h 后,小鼠系膜细胞在AngⅡ刺激下收缩明显更为剧烈,30min 后细胞面积显著缩小,面积变化值达到8947±3658pixels,显著大于正常对照组4268±2131pixels(P<0.05),面积变化的幅度达到正常对照组2 倍以上。1μmol/L A1AR 激动剂CHA 刺激48h 后,系膜细胞显现出类似于腺苷的对AngⅡ的显著收缩反应,面积变化值为9813±1640pixels,同样显著大于正常对照组(P<0.05),与腺苷刺激组接近。使用1μmol/L A1AR 抑制剂DPCPX 刺激48h 后,系膜细胞对AngⅡ的收缩反应减弱,仅为2613±2546 pixels,显著弱于腺苷及CHA 刺激组(P<0.05),也低于正常对照组,但差异不具有显著性。

图5 AngⅡ刺激前后系膜细胞面积变化。A、B、C、D:AngⅡ刺激前;E、F、G、H:AngⅡ刺激30min后;A、E:正常对照组(NC);B、F:腺苷刺激组;C、G:A1AR激动剂组(CHA);D、H:A1AR抑制剂组(DPCPX)。腺苷和CHA组AngⅡ刺激前后面积变化显著大于NC组;DPCPX组AngⅡ刺激前后面积变化不显著,明显弱于腺苷和CHA组。
3 讨论
3.1 腺苷及A1AR 在肾脏血流动力学调节中的作用
近年来,腺苷及其受体在血管舒缩调节中的关键作用备受关注。腺苷是由糖苷键连接腺嘌呤和核糖而成的嘌呤核苷,在生理情况下以低浓度广泛分布于身体各种组织的细胞间液中。低氧、缺血、炎症、应激等刺激条件下,腺苷大量生成,目前大多研究认为在以上病理生理过程中腺苷可起到一定的保护作用[3,4]。腺苷通过腺苷受体发挥作用,目前已知的腺苷受体 (adenosine receptor,AR)有3 大类,包括A1AR、A2AR、A3AR,其中A2AR 又分为2 个亚类:A2aAR、A2bAR、均为G 蛋白耦联受体。在大多数血管中,腺苷促进血管舒张主要由内皮细胞表达的A2aAR 和A2bAR 通过激活GSα和蛋白激酶A 通路介导[5]。而A1AR 仅表达在有限的部位,肾组织内主要表达在入球小动脉、球旁器颗粒细胞及近曲小管上皮细胞等[5~7]。入球小动脉给予A2AR 激动剂CGS21680 造成管壁舒张,给予A1AR 激动剂CHA 引起管壁收缩,提示与大多数血管相同,在入球小动脉,A2AR 主要舒张管壁,A1AR 则产生收缩管壁的作用[8~10]。A1AR 敲除的小鼠TGF 完全缺失[11,12],人为上调A1AR 表达可使TGF 加强[13],提示A1AR 是维持TGF 正常进行的关键蛋白。然而,随着技术的进步,人们发现选择性敲除入球小动脉表达的A1AR,再通过显微分离灌注观察单个肾小球 (可除外肾素作用),发现其TGF 并非完全缺失,仅为部分降低[1],提示入球小动脉并非唯一表达A1AR 并直接介导TGF调节的结构。
系膜细胞是连接并支持肾小球毛细血管袢的主要结构,具有舒缩功能,能够调节毛细血管袢滤过面积,有参与TGF 的结构和功能基础。我们查阅到A1AR 在系膜细胞的表达此前偶有报道,但主要是集中在大鼠标本中[6,14],对于小鼠的研究比较少,且其具体功能尚不完全清楚。因此我们希望通过本研究,证实A1AR 在小鼠系膜细胞的确有表达,并观察其表达部位,进而探究腺苷和A1AR激动剂、抑制剂对系膜细胞舒缩功能的影响,旨在为系膜细胞参与TGF 调节提供更多理论依据。
3.2 A1AR 在系膜细胞中的表达以及腺苷和A1AR 参与系膜细胞收缩的作用
本研究尝试使用文献报道较多的分样筛法分离小鼠原代系膜细胞[2,15],将肾皮质成分研磨后通过不同孔径的筛网,最终获得纯化并经过胶原酶消化的肾小球。此法获得的肾小球固有细胞主要有3 种:系膜细胞、内皮细胞和上皮细胞。原代上皮细胞和内皮细胞的培养一般需要从幼年动物肾脏提取,本实验选用6~8 周龄小鼠为青壮年,不利于上述2 种细胞存活。此外,内皮细胞的培养基通常需要依赖成纤维细胞生长因子,上皮细胞也需要特殊培养液,因此通过反复传代可达到系膜细胞的逐步纯化,最终通过标记物免疫荧光染色鉴定为系膜细胞。
我们使用免疫荧光染色和western blot 方法证实,A1AR 在小鼠系膜细胞的确也有表达,免疫荧光染色展示出A1AR 呈细颗粒状分布于系膜细胞胞浆中。本研究中腺苷对系膜细胞的增殖并没有明显的影响。在此前提下,为更好观察腺苷对系膜细胞收缩功能的影响,我们选择生长曲线实验当中的最高浓度,也是文献当中使用较多的浓度——100μM 的腺苷浓度刺激小鼠系膜细胞,观察收缩情况的变化。
1μM AngⅡ刺激下系膜细胞能够在30min 内发生显著的收缩,已经被多项研究采纳用于其收缩功能的评估。在AngⅡ的刺激下,正常系膜细胞能够表现出在数分钟内收缩的现象,不同状态的细胞收缩程度可能不同,因此被用作一种评估其收缩能力的常用指标[16,17]。本文中使用活体显微镜成像系统在Ang Ⅱ刺激后连续动态观察30min,评估腺苷、A1AR 激动剂及A1AR 抑制剂对小鼠系膜细胞收缩功能的影响,尚属首次尝试。我们观察到与正常对照组相比,系膜细胞在加入腺苷和A1AR 激动剂的培养基中培养48h 后,受到AngⅡ刺激后的30min 内收缩幅度明显增加,而在A1AR 抑制剂培养48h 后,AngⅡ引起的收缩幅度有减弱,并通过动态影像获得了更为确切的证据。同时,对AngⅡ刺激前和30min 时的细胞面积变化进行分析,得到更为量化的研究结果,证实了腺苷及A1AR 参与系膜细胞收缩的调节。
综上,本研究证实了腺苷及A1AR 是调节系膜细胞收缩的重要机制,提示系膜细胞的舒缩很可能也发挥了协同参与TGF 的作用。
