5 年生马尾松生长性状3 地点家系变异及评价
2020-12-08解懿妮刘青华蔡燕灵孙林山徐清乾周志春
解懿妮,刘青华*,蔡燕灵,孙林山,徐清乾,周志春
(1. 中国林业科学研究院亚热带林业研究所,国家林业和草原局马尾松工程技术研究中心,浙江省林木育种技术研究重点实验室,浙江 杭州 311400;2. 广东省林业科学研究院,广东 广州 510520;3. 湖北太子山林场管理局,湖北 京山 431822;4.湖南省林业科学研究院,湖南 长沙 410004)
树木的抽梢可以反映出树高的生长情况,是速生树种重要的选择性状之一。一般认为,抽梢长度越长,树木的树高生长越快[1]。不同树种的抽梢次数不同,抽梢次数是一个相对稳定的性状,受环境的影响较小,而年抽梢长度受环境的影响较大[2-4]。黄永权等[3]发现,3 年生火炬松(Pinus tadeaL.)年抽梢次数1~7 次,平均抽梢次数为4 次,91.53%的火炬松抽梢3~5 次,且抽梢次数越多,树高生长越大;张振等[5]得出,后续抽梢生长量、抽梢长度及抽梢次数均对树高的生长产生正向影响。由此可见,树木的抽梢对树高生长有直接的影响。
马尾松(Pinus massonianaLamb.)属松科(Pin aceae)松属(Pinus)植物,其速生丰产性显著、材性优良、产脂力高及适应性强,是我国亚热带地区重要的速生丰产用材和高产脂造林树种[6-7]。马尾松遗传改良的系统研究始于20 世纪80 年代初,现已完成第2 代育种并开始了第3 代育种[8]。子代测定是评价亲本材料遗传品质优劣的一条有效途径。林能庆[9]对139 个19 年生闽西马尾松优树自由授粉子代的生长性状、木材基本密度和结实量进行调查分析,筛选出71 个优良家系和69 个优良单株;谭健晖等[10]对29 个22 年生马尾松初级种子园半同胞家系的纤维形态指标进行分析,最终筛选确定113 号家系为纸浆材优良家系;张振等[11]、刘青华等[12]均对遗传交配设计子代测定林进行了优良杂交组合和优良单株的选择,选出的优良家系或单株可作为下一代种子园亲本材料或通过审定直接在生产上应用。已有的马尾松遗传变异研究多以单个地点的测定林为试验对象,即使涉及多个地点其覆盖的范围也较窄[13-16],由此筛选出的优良家系或无性系只针对单个地点,不能大范围进行推广与应用。在多个地点进行马尾松家系区域试验测定,可以选出适应特定区域的专用家系和适应不同环境条件的通用家系,做到适地适家系,提高马尾松的生产力[17-20]。本文以建立在湖北省京山市太子山、湖南省桂阳县和广东省信宜市旺埇3 个地点的5 年生马尾松46 个优良自由授粉家系试验林为研究对象,揭示其生长和抽梢等性状的遗传变异及性状相关,并进行家系稳定性和速生性评价,以期为各区试点及气候相似区域选择速生、适应性强的马尾松家系提供科学依据。
1 材料与方法
1.1 材料来源与试验设计
试验材料为福建、广西、湖南、广东和浙江马尾松种子园内的最佳优良无性系种子,其中,福建7 个无性系种子(ZP001、ZP242、ZP318、ZP325、ZP386、ZP773 和ZP80),来自福建1 代种子园;广西6 个无性系种子(GLP55、GLN1、GLN2、GLP50、GLP53 和GLP54),来自广西1 代种子园;湖南10个无性系种子(HN1~HN10),来自湖南1 代种子园;广东10 个无性系种子(GD1~GD10),来自广东1 代种子园;浙江10 个无性系种子(ZJ02、ZJ12、 ZJ25、 ZJ32、 ZJ39、 ZJ48、 ZJ53、 ZJ85、ZJ5148 和ZJ5153),来自浙江2 代种子园。
2013 年分别在湖北京山、湖南桂阳和广东信宜3 个地点的苗圃中培育成马尾松1 年生容器苗,2014 年1—3 月分别在当地营建家系测定林。试验采用完全随机区组设计,5 次重复,40 株小区(4 行10 列),株行距2.0 m×2.5 m。每个试验点设3 个对照(以湖南、广东和浙江3 地1 代种子园混系种子作为对照)。湖北京山、湖南桂阳和广东信宜3 个地点参试的家系(含对照)分别为44、43 和45 个,3 个地点共有的家系41 个。
3 个家系区域试验点属于热带和亚热带季风湿润性气候区。湖北京山太子山(30°58′ N,112°48′ E)年平均气温16.4℃,年平均降水量1 094.8 mm,无霜期240 d 左右,四季分明,光照充足,热量丰富,雨量充沛,气温适中。湖南桂阳(25°55′ N,112°34′ E)年平均气温17.7℃,无霜期277 d,气候温暖,四季分明,热量充足,雨水集中,春温多变,夏秋多旱,严寒期短,暑热期长。广东信宜旺埇(22°48′ N,111°21′ E)年平均气温26℃,年平均降水量1 433 mm,节气明显,无霜期长。
1.2 测定林调查与数据处理
2018 年底对湖北京山、湖南桂阳和广东信宜区域试验点优良家系测定林调查每小区中间两列的20 株植株,测定指标为树高(H)、胸径(D)、冠幅(Cr)、轮盘总数(WBN)、当年抽梢次数(NCS)、当年抽梢长度(LCS)及第1 次抽梢长度(LIS),广东信宜试验点未调查当年抽梢次数和第1 次抽梢长度。
方差分析时轮盘总数和当年抽梢次数经平方根数据转换。采用Bartlett χ2检验进行方差齐性检验。以小区单株测定值为单元进行单点方差分析,单点方差分析模型为:
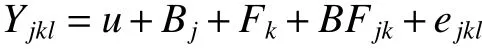
式中:Yjkl为单点试验第j 区组第k 家系第l 单株的观测值,u 为群体平均效应,Bj为第j 区组效应,Fk为第k 家系的效应,BFjk为第k 家系和第j 区组的互作效应,ejkl为机误。
以3 个地点共有的41 个家系小区均值为单元进行多点联合方差分析,多点联合方差分析模型为:
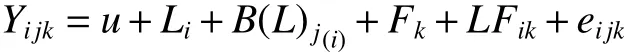
式中:Yijk为多点试验第i 地点内第j 区组第k 家系的均值,u 为群体平均效应,Li为第i 地点效应,B(L)j(i)为第i 地点第j 区组的效应,Fk为第k 家系的效应,LFik为第k 家系和第i 地点的互作效应,eijk为机误。
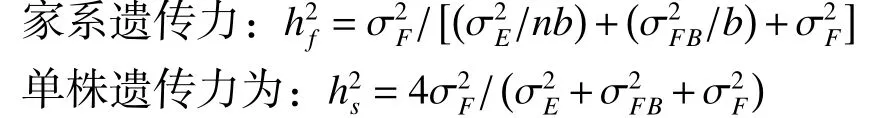
方差分析、多重比较、相关分析和主要遗传参数的估算均在SAS 8.1 软件中进行,分别调用GLM、DUNCAN、REG 和VARCOMP 模块进行分析[21]。家系树高和胸径性状的稳定性根据唐启义[22]编著的DPS 数据处理系统调用DPS 17.0 中的 “专业统计→品种区试及其他农学试验统计→有重复AMMI 模型” 进行分析。
AMMI(加性主效应乘积交互作用)稳定性分析模型为:

式中:Ykij为基因型k 在环境i 的第j 次重复的观测值,μ 是总体均值,αk是第k 个基因型与总体均值的离差(即基因型主效应),βi是第i 个环境与总平均的离差(即环境主效应),λr是第r 个交互效应主成分轴(IPCA)的特征值或奇异值,代表交互作用平方和中由该轴解释的部分,ψkr是第r 轴的基因型特征向量值,σir是第r 轴的环境特征向量值,n 为保留在模型中的主成分因子轴的总个数,ρki为提取n 个IPCA 轴后留下的残差(交互剩余项),εkij为试验误差。
家系与地点的相对稳定性参数(Dg)[23]为:

式中:n 为提取的IPCA 个数,ωn为权重系数,表示每个IPCA 所解释的平方和占全部IPCA解释的平方和的比例,ϒ2kn为第k 个基因型在第n 个IPCA 上的得分。Dg值越小,稳定性越高。
2 结果与分析
2.1 马尾松生长性状的家系遗传变异
单点方差分析结果表明(表1)表明:除湖南桂阳试验点的冠幅(Cr)性状外,3 个试验点的马尾松生长性状在家系间均差异极显著。湖北京山点胸径(D)、树高(H)和冠幅的均值分别是4.29 cm、3.28 m 和2.31 m,胸径最大的家系(GLN2)是最小家系(ZP242)的180.85%,树高最大的家系(GD7)是最小家系(ZP242)的153.70%,冠幅最大的家系(GLN2)是最小家系(ZJ39)的129.29%;湖南桂阳点胸径和树高的变幅分别为2.75~8.24 cm 和3.23~4.92 m,均值分别是5.47 cm和3.97 m。胸径最大家系(GLN2)是最小家系(GD8)的299.64%,树高最大家系(GD9)是最小家系(ZJ12)的152.32%;在地处热带地区的广东信宜点马尾松家系生长最快,5 年生平均胸径和树高分别为7.52 cm 和4.03 m,其最大家系的胸径和树高分别是最小家系的264.51%和204.62%。3 个试验点抽梢性状中,当年抽梢次数(NCS)、当年抽梢长度(LCS)和第1 次抽梢长度(LIS)的家系间差异极显著,家系变异系数为28.53%~38.91%,变异系数较大,表明在测定林中开展速生优质家系的再选择有很大的潜力。
2.2 马尾松生长性状的地点效应及家系与地点互作效应
方差齐性检验结果表明:3 个试验点各性状误差方差均不显著,因此对3 个试验点共有的41 个家系进行多点联合方差分析,结果(表2)显示:马尾松生长和抽梢性状的家系效应和地点效应均达极显著水平,说明地点对马尾松幼林生长和抽梢生长的影响较大,也同样与家系效应一样重要,意味着选择适生的栽植区域和立地条件有利于提高马尾松人工林生产力。5 年生马尾松家系在广东信宜点的树高(H)和胸径(D)高于湖南桂阳点1.51%和37.48%,高于湖北京山点22.87% 和75.29%。湖南桂阳点的当年抽梢次数(NCS)比湖北京山点高11.76%,广东信宜点的当年抽梢长度(LCS)分别比湖北京山点和湖南桂阳点高16.44% 和8.97%。除了受家系和区域试验点影响外,生长和抽梢性状还存在显著的家系×地点互作效应,即在一个地点生长表现好的家系在另一地点的表现不一定好(附表1),如HN4 家系树高和胸径在湖南桂阳点的表现均高于当地均值, 分别高出22.67%和12.98%,而在湖北京山点和广东信宜点均比均值低,树高在2 个试验点分别低于均值6.10% 和4.71%,胸径低于均值9.32% 和0.80%;家系GLN1、GD8、GLP55 和GD3 在湖北京山点和广东信宜点的生长表现均高于均值,在湖南桂阳点则低于均值。因此,为各区域配置马尾松良种时,应选出通用优良家系和各地的专用优良家系。
2.3 马尾松家系生长性状的遗传力估算
各区试点5 年生马尾松家系生长和抽梢性状的单株遗传力和家系遗传力估算结果(表3)显示:胸径、树高、轮盘总数、当年抽梢次数、当年抽梢长度和第1 次抽梢长度在3 个试验点具有较高的遗传力,家系遗传力为0.72~0.96,单株遗传力为0.45~0.88,说明这些性状受到较强的遗传控制。冠幅在湖北京山点和湖南桂阳点的单株遗传力和家系遗传力均较低,而在广东信宜点具有较高的家系遗传力(0.85)和单株遗传力(0.76)。该结果说明,马尾松的生长和抽梢性状皆受较强的遗传控制,为优良家系及优良家系内个体的选择和再选择提供了科学依据。

表1 各试验点马尾松家系生长性状的方差分析 Table 1 Variance analysis of growth traits of P. massoniana in each test site

表2 马尾松家系生长性状的多点联合方差分析 Table 2 Multi-site combined variance analysis of growth traits of P. massoniana
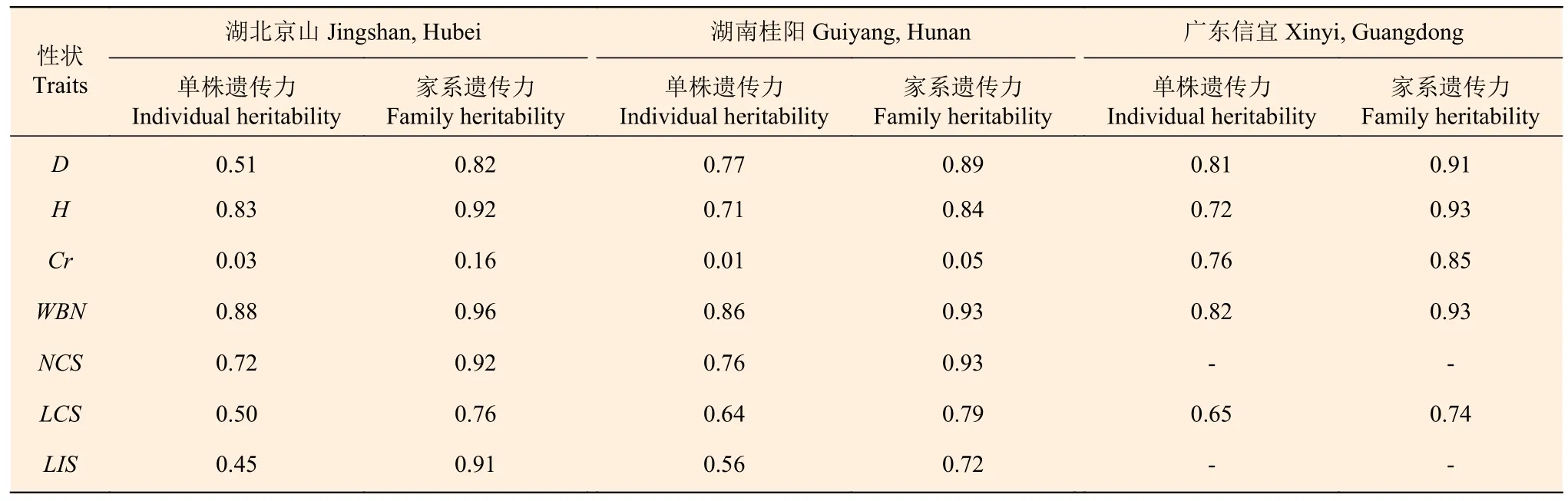
表3 马尾松家系生长性状的单株遗传力和家系遗传力估算值Table 3 Estimation of heritability of growth traits of P. massoniana
2.4 马尾松家系生长性状的相关性分析
对5 年生马尾松家系的生长和抽梢性状进行相关分析,结果(表4)表明:性状间遗传相关系数大多大于其表型相关系数。在湖北京山点和广东信宜点,家系胸径(D)、树高(H)和冠幅(Cr)间呈极显著的正相关,表型相关系数和遗传相关系数分别为0.405~0.916 和0.500~0.988,而湖南桂阳点的冠幅与胸径、树高间的相关性却不显著。当年抽梢长度(LCS)在3 个试验点均与树高、胸径间呈显著或极显著正遗传相关,说明当年抽梢生长量大的家系其树高、胸径也较大。湖北京山点和湖南桂阳点家系的胸径和树高与当年抽梢次数(NCS)呈正相关,而与第1 次抽梢长度(LIS)(固定生长)间均呈负相关,说明生长较快的家系抽梢次数更多,常有2 次及以上抽梢情况,也即多存在茎梢的自由生长,但固定生长量较小。在3 个试验点中也发现,家系轮盘总数(WBN)与树高、胸径、冠幅间均呈显著或极显著遗传正相关,因轮盘总数与抽梢次数密切相关,这也说明了抽梢次数对马尾松生长的显著正向效应。
2.5 马尾松家系生长性状多点稳定性分析
由于树高(H)和胸径(D)在地点效应、家系效应、地点与家系的交互效应均呈极显著差异,对树高和胸径进行线性回归模型及AMMI 模型分析,结果(表5)表明:树高联合回归仅能解释交互作用的1.82%,胸径联合回归仅能解释交互作用的25.56%,对交互作用的解释远远不够,信息遗漏程度极高,因此,普通线性回归模型不宜用于家系树高和胸径的稳定性分析;而AMMI 模型分析结果中看出,树高方面仅用第1 主成分分量就能解释交互作用的78.97%,胸径方面为79.31%,且均达到了极显著水平(P<0.01),表明用AMMI 模型解释家系与地点间的交互作用是可行的。进一步统计发现,树高和胸径AMMI 模型中2 项主成分分量就可解释交互作用的100%,说明AMMI 模型可以较为透彻地分析基因型与环境的交互作用,用来分析家系树高和胸径的稳定性可靠程度极高。
为进一步分析家系树高和胸径性状的稳定性,分别以家系平均树高、平均胸径为x 轴,以AMMI模型中第1 主成分分量IPCA1 值为y 轴作出树高和胸径AMMI-1 双标图(图1)。图中x 轴绝对值越大,家系的树高或胸径越大,y 轴绝对值值越大,家系的稳定性越差。树高和胸径AMMI-1 双标图显示,家系GD5 的IPCA1 绝对值均最小,稳定性最好,家系GD8 的IPCA1 绝对值最大,稳定性最差。

表4 马尾松家系生长性状的相关性分析 Table 4 Correlation analysis among growth traits of P. massoniana
为了更准确地比较家系的稳定性,需要计算出各家系的稳定性参数(Dg),Dg值越小,家系的稳定性越好。结果表明(附表2)表明:ZP325、ZP386、HN5、GD6、ZP318 家系的树高稳定性最好,GD8 家系的树高稳定性最差;ZJ5153、ZJ39、HN9、ZP318 和HN5 家系的胸径稳定性最好,GD8家系的胸径稳定性最差。
2.6 马尾松家系生长性状速生性评价
以树高(H)和胸径(D)同时高出各试验点均值5.00%,且高于各试验点相应1 代种子园对照的标准进行家系速生性评价,结果(表6)表明:湖北试验点速生家系有13 个,占比29.55%,分别是GD9、GD1、GLN2、GD7、GLP54、GLN1、GD8、GD2、GLP55、GD6、HN2、ZJ32 和HN9;湖南试验点的速生家系是GD9、HN4、GLN2 和GD7,共4 个;广东试验点中的速生家系共7 个,分别是GLN2、GD10、GD7、GLN1、GD8、GD2 和GLP55;其中,来自南部种源区广东的优良家系在各个试验点的速生家系中占较大比例。3 个试验点共有的马尾松速生家系为GLN2 和GD7,可作为湖北、湖南和广东3 地点速生、稳定型优良家系进行推广。
3 讨论
子代遗传测定是林木育种工作中一个重要的关键环节,对提高育种效率和合理制定育种策略等提供理论依据。本文通过对湖北京山、湖南桂阳和广东信宜3 个试验点建立的5 年生马尾松家系遗传测定林进行生长和抽梢性状研究,发现除湖南桂阳点的冠幅在家系间没有显著差异外,各个地点的生长和抽梢性状家系间均差异极显著,为优良家系的再选择提供了丰富的遗传变异基础,可进一步从中选出速生优质的家系[24-27]。多点联合方差分析结果表明,胸径、树高、冠幅、轮盘总数、当年抽梢次数、当年抽梢长度和第1 次抽梢长度存在显著的地点、家系以及家系×地点互作效应。广东信宜点的5 年生马尾松家系生长表现明显优于其余2 个试验点,这与广东信宜年均降水量大、平均气温高、无霜期长及土地肥沃有关,意味着环境差异对家系的生长有很大的影响。Alía 等[28]对西班牙5 个地点的19 年生43 个海岸松(P. pinaster Ait.)种源的生长表现研究发现,降雨量和温度等气候条件是影响树高和胸径的主要因素,土壤状况是次要因素;郑仁华等[19]对福建三明和仙游2 个试验点9 年生马尾松家系试验林的研究也得出,试验点立地条件影响马尾松的生长量,较好的立地条件可以提高马尾松的生长量。家系在不同试验点的生长显示,来自广东家系的幼林生长量较其它家系大,究其原因与产地效应有关。刘青华等[29]对24 年生马尾松种源生长的研究发现,马尾松种源胸径、树高和单株材积等性状与产地纬度有关,偏南部的种源生产力比偏北部的高,产地水热资源的差异对马尾松生长产生影响,水热资源较好的地区马尾松生长更好。
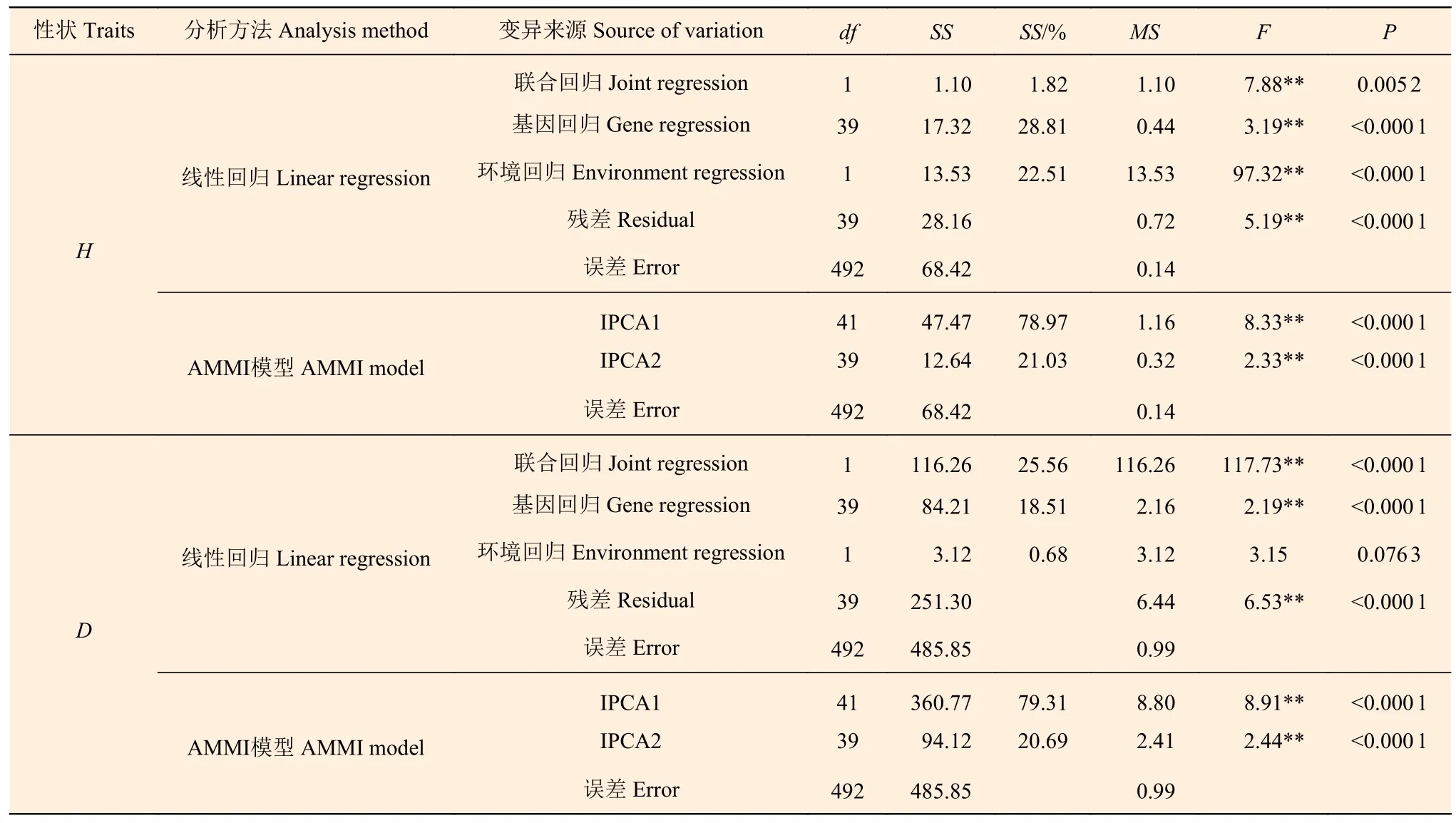
表5 树高、胸径线性回归模型和AMMI 模型分析 Table 5 Analysis of variance, linear model and AMMI model of Height, DBH

图1 AMMI 双标图Fig. 1 AMMI Biplot

表6 各试验点速生家系及稳定性参数 Table 6 Fast-growing families and stability parameters at each test site
遗传力大小对选择的效果有直接的影响[30]。3 个试验点5 年生马尾松树高、胸径的家系遗传力均大于0.80,受到强度的遗传控制,此结果与此前所报道的多数研究结果相比偏高。如刘青华等[31]对8 年生马尾松半同胞家系的研究得出,胸径和树高的家系遗传力分别为0.63 和0.47,白天道等[32]估算得出5 年生马尾松树高、胸径家系遗传力分别为0.523、0.598,6 年生马尾松分别为0.638、0.731。本研究中多数性状的家系遗传力均偏高,可能与研究的家系材料有关。本文研究的家系来自浙江、福建、湖南、广东和广西等地,其遗传背景来源广,家系间遗传差异大,进而导致遗传力估算值偏高。除胸径和树高外,3 个试验点轮盘总数和抽梢性状的家系遗传力均较高,也受到较强的遗传控制,而冠幅的单株遗传力和家系遗传力仅在广东信宜点较高,在湖北京山点和湖南桂阳点则较低,说明该性状遗传不稳定,还待进一步跟踪研究。胸径、树高、轮盘总数、当年抽梢次数、当年抽梢长度和第1 次抽梢长度的单株遗传力为0.45~0.88,受中度到强度的遗传控制,表明在优良家系中选择优良单株可达到更高的遗传增益。各试验点的性状相关分析显示,湖北京山点和广东信宜点家系胸径、树高与冠幅间呈极显著正相关,湖南桂阳点这3 个生长性状间也成正相关,但不显著。湖北京山和湖南桂阳的第1 次抽梢长度与当年抽梢次数和当年抽梢长度间的呈遗传负相关,这与张振等[5]得出初次抽梢生长量与后续抽梢生长量、抽梢次数、抽梢长度呈负相关的结论一致。树高、胸径与当年抽梢长度间呈极显著遗传正相关,轮盘总数与树高、胸径、冠幅间也呈显著或极显著的遗传正相关,意味着抽梢次数和抽梢长度能显著地影响树高的生长,抽梢次数越多,抽梢长度越长,进而使得树高更高,因此,选优时应选择自由生长量大的家系。湖南桂阳点性状间的相关关系与另外2 个试验点的相关相比不稳定,可能受环境影响较大。
AMMI 模型是目前国际上公认的分析品种稳定性的有效模型,其结合了主成分分析和方差分析,可以将基因型与环境间的互作效应分解开来,由此得出更准确的稳定性分析结果[33-34]。本研究根据5 年生家系胸径和树高AMMI 双标图和稳定性参数得出,ZP325、ZP386、HN5、GD6 和ZP318 家系的树高稳定性最好,ZJ5153、ZJ39、HN9、ZP318和HN5 家系的胸径稳定性最好,而GD8 家系的树高和胸径稳定性均最差。以高出平均胸径和树高5.00%,且高于各试验点相应的1 代种子园对照为速生性评价标准,在湖北京山、湖南桂阳和广东信宜3 个试验点分别得到13、4 和7 个速生家系,各试验点的速生家系可以作为专用家系在该地区进行良种配置,结合AMMI 模型稳定性分析,其中GLN2 和GD7 家系属于生长表现好,稳定性高的家系。根据刘青华等[29]24 年生马尾松种源试验结果,中部种源在中北亚热带地带的后期生长和生产力会超过南部种源,因此对于湖南桂阳点和湖北京山点的速生家系,还需进一步观察测定。
4 结论
5 年生马尾松半同胞子代测定林的生长性状和抽梢性状在家系间和不同地点间均存在丰富的遗传变异,具有很大的再选择空间。除冠幅外,各性状均受到较强的遗传控制。速生家系抽梢次数较多,抽梢长度较长,但固定生长量较短。湖北京山、湖南桂阳和广东信宜分别选出了13、4 和7 个速生家系;GLN2 和GD7 家系为速生兼稳定型优良家系,可在湖北、湖南、广东三省与本文试验地类似立地环境的地区推广。

附表 1 马尾松家系生长性状 Attached list 1: Growth traits of P. massoniana in each test site
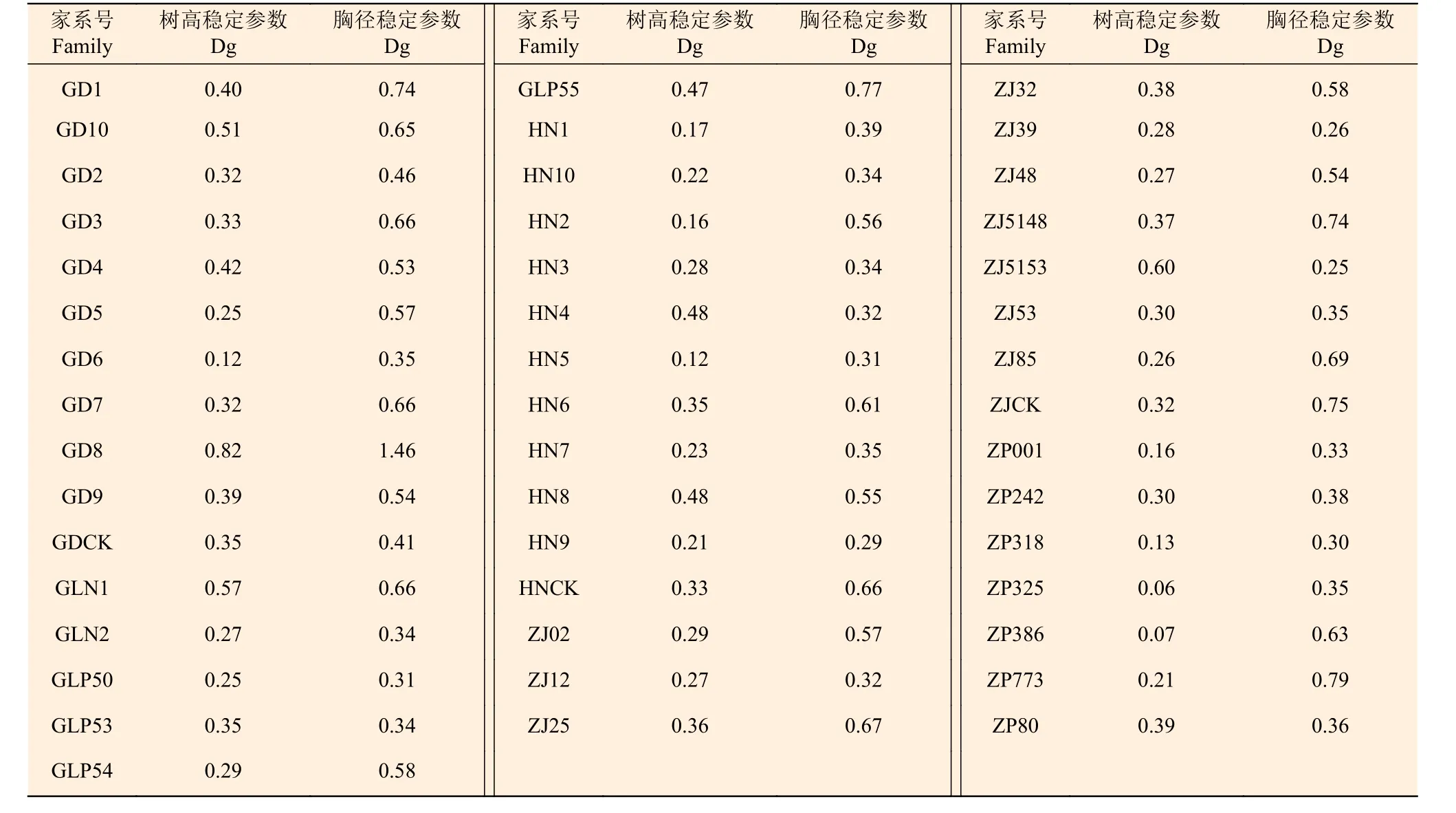
附表 2 马尾松家系稳定性参数 Attached list 2: Traits of P. massoniana stability parameters at each test site
