中国人群血型ABO 亚型的分子基础研究
2020-11-06范亮峰蔡晓红王钰箐王学锋
雷 航 ,范亮峰,蔡晓红,王钰箐,刘 曦,金 沙,沈 伟,陆 琼,向 东,王学锋,邹 纬
(1.上海交通大学医学院附属瑞金医院输血科,上海 200025;2.上海市血液中心血型参比室,上海 200051)
ABO 血型系统是输血和器官移植血型鉴定中最重要的血型系统,由血型之父Karl Landsteiner 于1900 年发现[1]。ABO 血型不相容的输血会导致患者发生严重的急性溶血性输血反应,从而导致急性肾功能衰竭,甚至死亡[2]。A、B、AB 和O 型4 种血型在全球范围内的分布频率有很大差异,我国人群的B等位基因频率从南向北逐渐增高,而O 等位基因频率则从南向北逐渐降低,A 等位基因分布频率差异趋势不明显[3]。在我国人群中,A 等位基因主要为A1.02,B 等位基因主要为B.01,O 等位基因则有O.01.01 和O.01.02 这2 种常见的基因型[4]。ABO亚型是一种遗传性ABO 弱表现型,为红细胞上H抗原表达正常但A 抗原和B 抗原的数量减少或质量发生改变,血浆中可伴有不规则的抗A 或者抗B抗体。ABO 亚型是导致ABO 血型鉴定困难、甚至误判的重要原因,而ABO 亚型的漏检或误判可导致急性溶血反应[5-6]。ABO 亚型分子检测方法,可提高血型变异型鉴定的准确率,目前已被广泛用于解决疑难ABO 血型鉴定,尤其是针对拟输血的患者,具有重要的临床应用价值。
本研究对1 545 505 名献血者和116 209 例受血者中检出的ABO 亚型进行表型与基因型关联性研究,探讨中国人群ABO 亚型的分布特点及背景。
资料与方法
一、资料
收集2014 年7 月至2019 年6 月间在上海交通大学医学院附属瑞金医院(以下简称瑞金医院)门急诊进行血型鉴定的患者116 209 例及在上海市血液中心及各郊县献血的不重复献血者1 545 505 名。另选择120 名正常ABO 血型的随机献血者作为对照。本研究纳入的样本来源均不包括新生儿、血液系统疾病患者或近1 个月内有ABO 不同型输血史的患者。标本类型均为乙二胺四乙酸二钾抗凝的外周全血。
二、方法
1.血清学检测: 所有血标本均先经Galileo 全自动血型分析仪(Immucor 公司,美国)、Autovue 全自动血型仪(强生公司,美国)和IH-1000 全自动血型仪(Bio-Rad 公司,美国)进行ABO 和Rh 血型鉴定。对ABO 正反不符的血标本采用试管法进行ABO 亚型鉴定[7],使用的试剂包括抗A 和抗B 血型定型试剂(单克隆抗体)、抗A 和抗B 单克隆抗体、抗H 单克隆抗体、抗A1 凝集素、A2 细胞、抗体筛选红细胞试剂盒和人ABO 血型反定型用红细胞试剂盒(上海血液生物医药有限责任公司)。脐血O 细胞、人源多克隆抗A、人源多克隆抗B 试剂为实验室自制试剂(效价1∶128)。ABO 亚型均以血清学标准进行分类[1-3,8]。
2.基因组DNA 抽提与PCR 扩增:采用血液基因组DNA 抽提试剂盒[天根生化科技(北京)有限公司]抽提外周血标本中的基因组DNA。对544 例ABO 亚型的ABO 基因启动子、增强子、7 个外显子及其侧翼序列和内含子6 进行PCR 扩增。所用相关PCR 引物序列参考文献[9]进行设计。
3.PCR 产物纯化、测序及克隆:采用DNA 琼脂糖凝胶纯化试剂盒[天根生化科技(北京)有限公司]纯化并回收PCR 产物,用末端标记双脱氧法对ABO 基因的启动子、增强子、7 个外显子以及其侧翼序列和内含子6 PCR 产物纯化后进行测序。将PCR 产物克隆到pMD18-T 载体进行单倍型鉴定。
4.ABO 等位基因的命名方式
核苷酸和氨基酸采用国际输血协会的基因突变和等位基因术语进行命名。ABO 基因序列比对分析的参考序列为ABO*A1.01 和ABO*B.01 等位基因序列。若还未被国际输血协会命名,则按文献首次报道中的命名显示。
结 果
一、ABO 亚型的表型鉴定
经对上海市血液中心的献血者及瑞金医院门急诊患者进行ABO 亚型初筛和确认检测,确定的亚型表型和例数分布情况见表1。本研究在1 661 714 份血型鉴定标本中共检出617 例ABO亚型,其中1 545 505 份来自献血者,检出ABO 亚型590 例,检出率为3.82/万;116 209 份来自门急诊患者,检出ABO 亚型27 例,检出率为2.32/万。总体调查人群的ABO 亚型检出率为3.71/万,检出率最高的表型为B(A)(0.066‰),其次是B3(0.045‰)和Bx(0.039‰)。在检出的所有ABO 亚型中,B(A)表型是检出构成比(17.83%)最高的亚型,其次则是B3(12.16%)和Bx(10.53%)表型(见图1A)。

表1 不同来源的ABO 亚型表型检出数及检出率
二、ABO 亚型的基因检测
本研究对544 例ABO 亚型样本进行基因检测和分析,在381 例无血缘关系的个体中共检出42 种ABO 亚型等位基因(见表2)。在42 种检出的ABO亚型等位基因中,ABO*BA.04 是检出构成比最高的亚型等位基因,其检出构成比为25.20%,随后为ABO*BA.02(11.55%)和ABO*cis-AB.01(9.71%)(见图1B)。
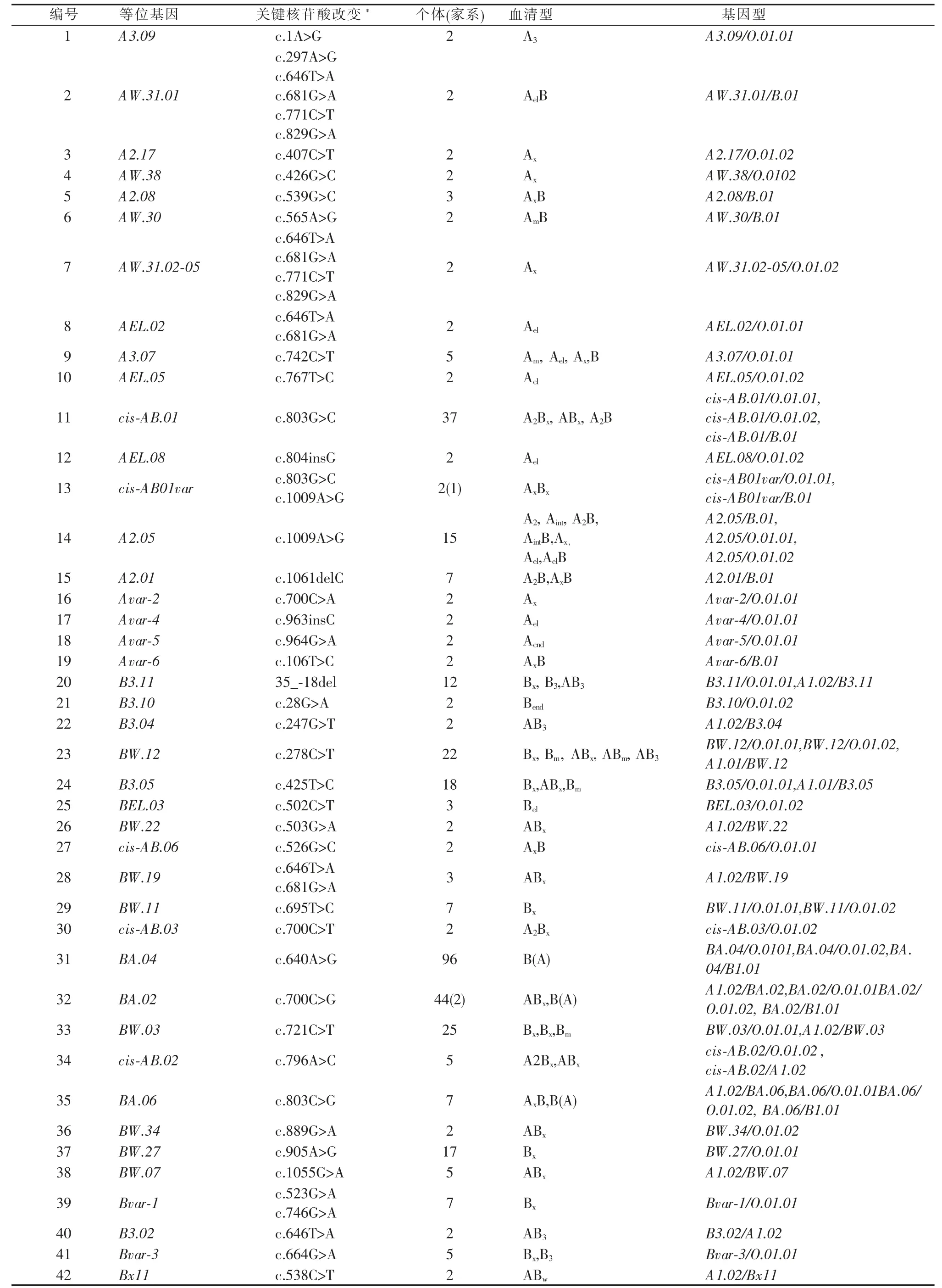
表2 本研究中检出的42 种ABO 亚型等位基因

图1 总调查人群中ABO 亚型的各表型和基因型的检出构成比
另163 例ABO 亚型在目前测序区域里未发现突变位点,占基因检测总数的30.0%。在这些个体中,有16 例在ABO 基因5′UTR 增强子测序时检测到3 种等位基因,证实存在嵌合现象;虽然由于常见A、B、O 等位基因间存在较多相同的多态性位点,导致在这16 例样本的单个外显子测序中无法直接判断是否同时存在A、B、O 3 种等位基因。根据血清学检测结果,这些嵌合体个体的红细胞与抗A 或抗B 试剂反应时,表现出不同程度的混合视野凝集,出现类似于以下亚型的双群红细胞特征如AendB、A3B、A3B3、Bend、ABend、B3、AB3。另外3 例表型特征符合Ael和ABel,但检测到的基因型分别为O/O 和A/O 型,推测也可能为微嵌合体。其余的血清学分类如下,65 例为“3”型,34 例为“x”型,19 例为“el”型,18 例为“m”型,8 例为“end”型。
讨 论
一、ABO 亚型表型检出率的分析
近年来,ABO 亚型的检出率从最早的1.5/万[10]到近期的2.8/万[11],略有增长的趋势,这可能与血型检测相关技术的改进有关。另外,由于有些A2亚型个体在血清学鉴定时不会出现正反定型不符的情况,因此这些A2亚型个体会被漏检,故本研究得出的检出率可以看作是表型频率的下限值。本研究在约166 万例外周全血样本中共检出ABO 亚型617 例,检出率为3.71/万。检出的617 例ABO 亚型共包含28 种ABO 亚型表型,其中B(A)表型的频率最高,其检出构成比为17.83%,其次为4 种与B3和Bx相关的亚型,提示这几种亚型在我国人群中较为常见。
二、ABO 亚型等位基因检出率的分析
目前已报道的ABO 亚型等位基因约有300 余种[12-13],本研究对381 例ABO 亚型样本进行基因检测,共检出42 种已知ABO 亚型等位基因,包括启动子区域突变1 种;可导致氨基酸序列改变的ABO亚型等位基因37 种;可导致外显子跳跃的剪切位点突变等位基因1 种,以及可导致截短蛋白的移码突变等位基因3 种。在已检出的42 种ABO 亚型等位基因中,ABO*BA.04 是检出构成比最高的亚型等位基因,其检出构成比为25.20%,其次为ABO*BA.02 (11.55%)和ABO*cis-AB.01(9.71%)(见图1B)。
三、ABO 亚型表型与基因型关联性的分析
对本研究中检出的等位基因进行分析发现,ABO 亚型等位基因所对应的表型可与首次检测报道时的表型不一致,即一种基因型可对应多种表型,其原因可能是一种亚型等位基因会受其共表达的A 或B 基因的竞争或协同作用影响,或使用的抗血清试剂存在差异所致。如最常见的BA 等位基因,当其与A 基因共表达时,个体可表现为ABx表型;而当其与B 基因共表达时,个体则表现为AxB表型。故BA 等位基因导致的AxB 或者ABx的表型,与单纯的A(或B)亚型和正常B(或A)基因共表达时的表型在血清学上很难区分。根据本研究数据,BA 等位基因频率明显高于A 亚型和B 亚型等位基因,因此当出现AxB 或ABx表型时,应首先考虑B(A)亚型的可能性。本研究中所检测到的42 个亚型等位基因中,与“x”表型相关的有29 个(其中17 种A 亚型中有9 种,18 种B 亚型中有14 种,以及全部6 种AB 杂交等位基因),提示“x”表型是我国人群中基因异质性最强的表型;而B(A)和cisAB亚型作为基因型种类较为集中且明确的亚型表型[11],仅检出9 种相关基因型。此外,本研究还检出6 种A3或B3亚型相关等位基因,从目前已报道的引起“3”型的突变类型可以发现,其突变类型有引起催化区氨基酸置换的错义突变,以及引起穿膜区和茎状区部分丢失的剪接突变[14-15],但“3”型所具有的独特混合视野表型的形成机制目前尚未完全阐明。
四、ABO 亚型形成机制的分析
目前对于ABO 亚型形成的机制研究发现,与ABO 亚型有关的ABO 基因突变类型有点突变、缺失、插入和基因重组等多种[11],如A1.02 等位基因是由于A 等位基因在156 位发生亮氨酸取代脯氨酸突变形成,生成的糖基转移酶并不影响酶活性[16]。许多亚型等位基因是由2 个ABO 等位基因的部分序列发生杂交而形成[17],而这些杂交等位基因的重组多发生于内含子6 中[18]。杂交基因的产物有无活性与杂交基因6 号外显子的来源有关,若来源于O1 或O1v,则基因产物无活性;若来源于A 或B 等位基因,则基因产物具有酶活性,其特异性由外显子7 决定。其他导致ABO 亚型的分子机制还有导致翻译提前终止的无义突变[3];导致无法形成正常功能酶的剪接位点异常[19];导致酶发生特异性改变[20]或催化活性减弱[21]的特定位点氨基酸置换;导致ABO 基因转录水平下降的启动子区域内缺失突变或单点突变[22-23];导致ABO 基因表达发生改变的1 号内含子调控元件突变[24-26]。
本研究在163 例ABO 亚型样本中未检测到突变,除外19 例可能为嵌合体后,剩余最常见的类型为“3”型,其次为“x”型,这可能与这2 种表型在我国人群中的比例较高、发生机制复杂及多样化有关。目前已发现的“3”型表型相关的基因突变多与基因调控或剪接位点突变有关。由于某些内含子深部突变也可能导致剪接异常[27-28],因此对这些编码区及其侧翼序列、已知调控区域未发现突变的个体,需要进行转录水平、转录后和翻译后水平的研究,以进一步探明ABO 抗原表达异常的分子机制。
总之,本研究在中国人群中进行了大规模ABO亚型表型与基因型关联性研究,从分子水平对381 例ABO 亚型标本涉及到的42 种ABO 亚型等位基因进行了系统分析。结果显示,B(A)表型是我国人群中最常见的ABO 亚型,B3和Bx相关的亚型次之;最常检出的亚型等位基因则是ABO*BA.04、ABO*BA.02 和ABO*cis-AB.01。针对ABO 亚型等位基因,需要进一步行基因突变与蛋白表达、结构与功能间关系的研究,以阐明突变导致ABO 抗原表达减弱的分子机制,为将来ABO 亚型的分子诊断提供可靠的检测靶点。
