茶多糖调控肥胖作用研究进展
2020-10-16欧阳建周方卢丹敏李秀平黄建安刘仲华
欧阳建,周方,卢丹敏,李秀平,黄建安,3,刘仲华,3*
茶多糖调控肥胖作用研究进展
欧阳建1,2,周方1,2,卢丹敏1,2,李秀平1,2,黄建安1,2,3,刘仲华1,2,3*
1. 国家植物功能成分利用工程技术研究中心,湖南 长沙 410128;2. 湖南农业大学茶学教育部重点实验室,湖南 长沙 410128;3. 教育部植物功能成分利用协同创新中心,湖南 长沙 410128
随着人们生活水平的提高,肥胖的发病率不断攀升,已经成为严重的健康问题。茶多糖作为一种与蛋白质结合的酸性杂多糖,能够通过调节食物摄取和能量吸收、调节脂肪生成、增强抗氧化防御酶活性和减轻炎症、调节肠道菌群紊乱和维护肠道屏障完整性等不同途径有效调控肥胖。综述了近年来茶多糖调控肥胖功能及其作用机理的研究进展。
茶多糖;肥胖;调控;作用机理
世界范围内,超重和肥胖人数的上升趋势越来越明显,肥胖已成为21世纪最严重的公共卫生问题之一[1-2]。过去40年,全球肥胖率增加了8倍[3]。目前我国的肥胖率也在不断攀升,7~18岁城市男女生超重及肥胖检出率已经分别达到28.2%和16.4%,这给我国整体健康水平带来巨大隐患[4]。
肥胖是指可损害健康的异常或过量的脂肪累积。当能量摄入和消耗长期不平衡时会刺激能量以甘油三酯(Triglyceride,TG)的形式在脂肪细胞中储存起来,导致脂肪组织重量、人体体重增加,最终产生肥胖[5]。超重和肥胖是肝病、心血管疾病和糖尿病等慢性疾病的主要危险因素[6-7]。世界卫生组织对肥胖的定义为身体质量指数(BMI)≥30 kg·m-2。一般认为,肥胖的发生是由多种因素引起,包括饮食、生活方式、遗传特点和社会因素4个方面,与能量吸收代谢、脂肪合成、体内氧化应激和肠道菌群等密切相关[8-10]。目前市场上的常用减肥药物有胰脂肪酶抑制剂奥利司他和食欲抑制剂西布曲明,其副作用较大[11]。肥胖一旦形成就很难控制,因此早期预防是控制肥胖的关键。
茶作为世界上最受欢迎的天然植物饮料之一,因富含茶多酚、茶氨酸、茶多糖等具有独特保健功能的生理活性物质而被全球越来越多的消费者所接受。与茶多酚相比,茶多糖(Tea polysaccharides,TPS)的健康功能研究相对较少。近年来,国内外的研究表明,茶多糖具有抗氧化、抗炎、免疫调节和保肝等多种生物活性[12-13],特别是茶多糖在控制肥胖等代谢综合征方面表现出良好的效果[14-16]。本文主要综述了茶多糖调控肥胖功能及其作用机制,并对茶多糖调控肥胖研究进行展望。
1 茶多糖组成及吸收代谢
茶多糖是一种与蛋白质结合的酸性杂多糖,主要由中性糖、糖醛酸和蛋白质组成。茶多糖在茶叶中的含量与茶叶的老嫩程度及茶树品种有关,一般粗老叶高于嫩叶,大叶种高于小叶种。由于原料和工艺不同,不同茶类的多糖组成也存在一定差异。Chen等[17]研究发现,乌龙茶多糖中含有26.66%中性糖、40.65%的糖醛酸和19.59%蛋白质,分子量为1.28×106Da,其中中性糖由-鼠李糖、-半乳糖、-阿拉伯糖和-葡萄糖组成,摩尔比为1.37∶1.00∶1.89∶1.30;而绿茶多糖中含有63.5%中性糖、25.14%的糖醛酸和8.63%的蛋白质,中性糖组分有核糖、鼠李糖和葡萄糖等。同样,采用大孔阴离子交换树脂柱层析法能够从茶叶中获得中性多糖(Neutral tea polysaccharides,NTPS)和酸性多糖(Acid tea polysaccharide,ATPS),中性茶多糖中总糖含量占82.7%,且以半乳糖为主,而酸性茶多糖中总糖含量占85.5%,以鼠李糖、阿拉伯糖、半乳糖和半乳糖醛酸为主。茶多糖的结构复杂。其结构可分为一级和高级结构,一级结构包括糖苷键类型、单糖组成、支链长度与位置等,而以氢键结合形式的主链形成的聚合体构象,包括单糖残基形成的有序空间内的规则构象构成其高级结构[18]。
茶多糖相对分子质量、糖苷键位置、单糖组成和单糖序列等都对其结构组成具有重要影响,与其生物活性关系密切。邵淑宏[19]研究表明,乌龙茶发酵程度的增大能提高多糖中的糖醛酸和蛋白含量,增强对-葡萄糖苷酶的抑制作用和抗氧化活性。Chen等[20]从绿茶中分离纯化3种茶多糖偶联物进行研究,发现相对分子量较低和蛋白含量较高的茶多糖偶联物对提高其生物活性具有重要作用。同时,茶多糖中鼠李糖含量、甘露糖含量和半乳糖醛酸含量与抗氧化活性之间呈极显著正相关,且糖醛酸含量更高的茶多糖能更好的抑制前脂肪细胞的增殖与分化,表现出更好的降脂减肥活性[18, 21]。刘海燕等[22]等研究发现,小分子质量、单糖种类和半乳糖比例的增加有助于TPS发挥其生物活性,抑制脂肪吸收,预防肥胖。
毒理学评价研究中将L-O2细胞暴露于25~400 μg·mL-1的茯砖茶多糖中24 h后,其细胞存活率仍保持在80%~120%,说明茯砖茶多糖未表现出明显的细胞毒性[23]。动物试验中,小鼠饲喂5.0 g·kg-1茶多糖后,试验过程无死亡,给药后小鼠胸腺、心脏、肾脏和肝脏均无毒性表现,且试验组与对照组的肝、脾、胸腺指标均无明显差异,说明茶多糖安全无毒[24]。
同时,采用口服或腹腔注射等不同的TPS摄入方式,其吸收代谢也有一定差异。研究表明,模拟唾液、胃和小肠消化前后,茯砖茶多糖的分子量、单糖含量和还原糖含量均无变化,其可以通过消化系统且不被破坏,到达大肠后可被其肠道菌群分解利用[24]。韦铮等[25]研究表明,茶多糖的抗氧化活性在经过模拟的胃肠消化条件时与周围环境的作用时间和pH无关,只与茶多糖的浓度有关。然而,也有研究表明绿茶多糖在唾液消化过程中不能被降解,在模拟胃肠消化过程中可以被分解成更小的片段,作为肠道菌群的主要能量和碳源,并刺激其生长产生有益健康的化合物[26-28]。张高帆等[29]研究显示,给小鼠腹腔注射200 mg·kg-1剂量的茶多糖组分TPF70后,其各肠段都能消化吸收,其中空肠和回肠吸收最快,在1 h出现吸收高峰,其后下降;胃、十二指肠和大肠在1 h出现高峰,并有部分残留于粪便中排出体外。同时,其在内脏中的浓度比消化道高,以肝脏中的浓度最高,并在3 h达到高峰。
2 茶多糖对肥胖的调控作用
2.1 细胞试验
肥胖是一种营养代谢紊乱,临床表现为脂肪细胞增生、肥大。脂肪细胞增生是由前脂肪细胞的增殖和分化引起的,其紊乱可导致脂肪组织内脂质过度沉积,直接影响机体构成和肥胖及相关疾病的发展。3T3-L1是一种被广泛用来研究对脂肪生成影响的细胞系,李娟等[21]研究发现,添加100 μg·mL-1茶多糖能显著抑制甘油三酯的合成与3T3-L1前脂肪细胞的分化。其中,绿茶多糖、乌龙茶多糖和红茶多糖使前脂肪细胞的分化率分别显著下降至62.00%、97.24%和82.95%。脂肪酶与肥胖之间具有密切关系,利用脂肪酶抑制剂可有效抑制肠道脂肪酶的活性,影响体内的脂肪代谢,减少体内脂肪积累和合成,从而达到控制和治疗肥胖的目的[30-31]。李祥龙等[32]研究表明,黑茶茶多糖和茶褐素对脂肪酶也具有显著的抑制作用,其中普洱茶多糖的抑制效果明显,其半抑制浓度仅为47.57 mg·mL-1。茯砖茶多糖与茶褐素为可逆竞争性抑制关系,而六堡茶、普洱茶与康砖茶多糖为可逆竞争性与非竞争性混合型。此外,-葡萄糖苷酶抑制剂能降低小肠上消化酶的活性,延缓机体吸收葡萄糖的速率,研究发现青砖茶多糖抑制-葡萄糖苷酶活性的效果与阳性对照品阿卡波糖接近,且两者的半抑制浓度无显著性差异,而当大红袍TPS-3浓度为1 mg·mL-1时,其对-葡萄糖苷酶的抑制率达到43.4%[19,33]。
2.2 动物试验
高脂饲料所致的动物肥胖模型与人类主要因能量输入过多而产生肥胖最为接近,目前已广泛应用于肥胖及其相关疾病的发生、发展和作用机制的研究[34-35]。Ren等[36]研究发现,400 mg·kg-1和800 mg·kg-1含硒绿茶多糖可显著改善高脂饮食(High-fat diet,HFD)喂养小鼠的胰岛素敏感性,减少肝脏脂肪变性和氧化应激损伤,并使氧化应激和肝脂质恢复至接近正常值。刘海燕等[22]研究发现,对小鼠进行高脂饮食处理的同时进行白茶多糖(300 mg·kg-1)灌胃8周,与模型组相比,其体重降低26.63%,脂肪湿重降低46.31%,皮下脂肪细胞直径降低27.96%,对肝损伤和炎症具有良好的预防作用,且其预防肥胖的效果较枸杞多糖和桑叶多糖更佳。李清亮[37]研究表明,1 mg·mL-1黄大茶多糖可以调节HFD诱导小鼠的肠道菌群紊乱,维护肠道屏障功能,从而调节肥胖。此外,茶多糖还可以有效降低HFD肥胖大鼠的体重增长,500 mg·kg-1的六堡茶多糖灌胃给高脂血症肥胖大鼠干预4周后,与模型组相比其体重受到明显抑制,同时其肝脏脂滴明显减少、变小,肝脏脂肪变性减轻[38]。郭郁等[39]研究表明800 mg·kg-1的TPS灌胃肥胖大鼠6周后,与模型组相比TPS组对体重增长和脂肪质量的抑制率分别为12.63%和24.18%,血清中脂联素(Adiponectin,ADP)和TG分别升高49.95%和降低41.56%。400 mg·kg-1的红茶多糖持续给高脂饮食大鼠灌胃45 d后其体重和附睾脂肪细胞数量与正常饮食组接近,与模型组相比体重增长降低16.78%,血清中ADP上升了50.53%,肝脏中TG和丙二醛(Malondialdehyde,MDA)分别下降33.34%和21.39%,粪便中短链脂肪酸含量增加了69.08%,从而可控制高脂饮食引起的肥胖[14]。另外,500 mg·kg-1的普洱茶多糖饲喂高脂饮食小鼠28 d对小鼠的正常发育和饮水量没有影响,但能有效抑制血脂升高,与高脂模型组相比,其血清中TG、低密度脂蛋白胆固醇(Low density lipoprotein-cholesterol,LDL-C)和总胆固醇(Total cholesterol,TC)分别下降40.60%、24.38%和25.36%[40]。
3 茶多糖调控肥胖的作用机制
3.1 茶多糖减少食物摄取和抑制脂肪吸收
身体能量的动态平衡为身体的能量摄入等于消耗。当在某一特定时间内能量摄入超过消耗增加体重,或者,能量消耗超过摄入减少体重时,会出现一个新的稳态平衡设定值,在能量平衡的动态阶段控制体重要比在能量平衡的稳态状态下控制体重更容易[41]。能量稳态是由来自下丘脑神经回路、肠道、脂肪组织和其他周围器官的信号来平衡,这些信号有助于调节我们的食欲、进食行为和饱足感[42]。动态的能量平衡对于预防肥胖十分重要,进食行为是获取能量的关键。研究表明,400 mg·kg-1的茶多糖可以有效降低高脂饮食大鼠的日常食物摄入量和食物的利用率,减少能量的摄入和脂肪的积累,减轻体重和Lee’s指数,从而控制高脂饮食引起的肥胖[14,43]。
不良饮食是肥胖发生的重要原因,高脂高糖食物中含有大量能量,长期食用易使身体动态能量平衡失衡,减少对食物中能量的吸收是预防肥胖的重要方式[41,44]。食物摄入后的脂质在小肠内乳化、水解和吸收,胆固醇和脂肪酸可被小肠顶端表面的脂质转运体作用进入肠上皮细胞,被吸收的脂质被包装成乳糜微粒分泌到淋巴系统并循环[45]。体外研究表明,茶多糖可以干扰脂质的消化吸收。采用Caco-2细胞模型研究了绿茶提取物和天然绿茶TPS对水稻淀粉消化后葡萄糖转运的影响发现,TPS能抑制淀粉水解成葡萄糖,显著降低葡萄糖的扩散速率和比例,限制葡萄糖在肠道细胞中的运输,有效降低餐后血糖水平[46]。同时,TPS处理可以降低高脂饮食大鼠对脂质和脂肪酸的消化和吸收,显著改善脂质水平和脂质氧化,抑制肝脏中总胆固醇和总脂质的积累,刺激胆固醇转化为胆汁酸,通过增加粪便排泄量达到降低胆固醇的作用,从而有效预防和改善肥胖[15,38,43]。
3.2 茶多糖调节脂肪生成
脂肪组织在调节人体能量平衡和营养稳态方面发挥着重要作用[11]。人体内有两种脂肪组织,即棕色脂肪组织(Brown adipose tissue,BAT)和白色脂肪组织(White adipose tissue,WAT),它们在结构和功能上有所不同[47]。BAT主要分布在小型哺乳动物和新生儿中,专门通过产生热量来消耗能量;而WAT主要分布于成年人中,以甘油三酯的形式储存多余的能量[47-48]。超重或肥胖的明显特征是脂肪细胞数量和大小因增生和肥大而过度增加[49]。针对脂肪细胞的生命周期和代谢功能,抑制脂肪细胞增殖和分化,以及调节能量代谢是预防和治疗肥胖的有效途径[45]。李娟等[21]研究发现,100 μg·mL-1的TPS可以显著抑制3T3-L1前脂肪细胞的增殖,增加G0/G1分化期细胞数量抑制前体细胞向成熟脂肪细胞的分化,激活磷酸腺苷活化蛋白激酶(Adenosine monophosphate-activated proteinkinase,AMPK)信号通路调节脂肪因子的表达,最终减少TG和成熟脂肪细胞的积累。
动物体内脂质的主要来源有两种,一种是胃肠道的直接吸收,另一种是肝脏和脂肪组织中合成。此外,脂肪的分解代谢和氧化主要在肝脏和骨骼肌中进行。一系列研究表明,TPS可以通过调控脂质代谢基因的表达和脂肪生成来调节脂肪总量、脂肪区域和体重的变化[50-52]。高剂量的TPS干预(800 mg·kg-1·d-1)可以上调高脂饮食饲养小鼠的肉碱棕榈酰基(Carnitine palmitoyl transterase-1,CPT-1)的表达,下调过氧化物酶体增殖物激活受体(Peroxisome proliferator-activated receptor gamma,PPAR)、固醇调节元件结合蛋白1c(Sterol regulatory element-binding protein-1c,SREBP-1c)、脂肪酸合酶(Fatty acid synthase,FAS)和肝X受体(Liver X receptor,LXR)的表达,显著降低脂肪指数和脂肪细胞面积,增强脂肪酸的氧化和分解以及抑制脂肪生成和积累,从而调节肥胖[43,53]。Wu等[14]对基因表达谱阵列筛选出参与肝脏和附睾组织脂质代谢的13个基因进行研究发现,TPS可通过影响脂质代谢的差异基因表达调节包括胆汁酸分泌、转化生长因子信号传导、胰岛素信号传导、糖脂代谢、脂肪酸降解和AMPK信号通路等代谢,抑制脂肪形成、加速脂肪消化和促进脂肪分解。
3.3 茶多糖的抗氧化和抗炎作用
越来越多的研究显示,氧化应激和慢性低度炎症是与肥胖等代谢性疾病的起因、发病机制和发展密切相关的两个重要机制[36,53-54]。事实上,内脏脂肪组织堆积会引发促氧化和促炎症状态,而氧化应激又可通过刺激白色脂肪组织的沉积和改变食物摄入,增加前脂肪细胞的增殖、脂肪细胞的分化和成熟脂肪细胞的大小,从而引发肥胖[55-56]。
肥胖作为一种慢性疾病,表现为身体脂肪的不断增加。脂肪组织不仅是甘油三酯的储存器官,还能产生具有生物活性的脂肪因子,包括具有炎症功能的白细胞介素6(Interleukin,IL-6)、肿瘤坏死因子(Tumor necrosis factoralpha,TNF-)和调节食物摄入量的瘦素(Leptin,LEP)。分泌的脂肪因子反过来刺激产生活性氧,对不同器官造成直接或间接损害[55]。随着氧化还原状态的改变和代谢风险的增加,氧化应激是肥胖的一个后果,也是肥胖进一步发展的诱因。随着脂肪组织的增加,谷胱甘肽过氧化物酶(Glutathione peroxidase,GSH-PX)和过氧化氢酶(Catalase,CAT)等抗氧化酶的活性明显下降,不断产生的活性氧和抗氧化能力的下降最终会导致机体产生与肥胖相关的各种代谢综合征[55]。一系列试验表明,TPS具有良好的抗氧化和抗炎作用,能有效调控肥胖[23,36,53]。超氧化物自由基被认为是一种主要的活性氧,细胞试验表明,茯砖茶多糖浓度超过500 μg·mL-1时对超氧化物自由基的清除能力超过85%,从而有效减少DNA、蛋白质和脂质的氧化损伤[23]。动物试验中,Mao等[38]发现500 mg·kg-1的高剂量六堡茶多糖能显著恢复由HFD诱导的高脂血症大鼠血清和肝脏中GSH-PX和CAT活力下降,显著提升肝脏中的谷胱甘肽酶(Glutathione,GSH)水平,而治疗组血浆中的MDA水平呈显著地剂量依赖性降低。Ren等[36]研究表明,紫阳绿茶多糖能显著恢复果糖引起的小鼠肝脏总超氧化物歧化酶(Total superoxide dismutase,T-SOD)和GSH-PX活性的下降,减轻胰岛素抵抗、肝脏氧化应激和肝脏脂肪变性,有效预防非酒精性脂肪肝的发生。此外,TPS和茶多酚的协同作用能进一步提高肝脏的抗氧化能力,且比TPS或茶多酚单独处理具有更强的抗氧化作用[57]。然而,有研究表明茶多酚是粗茶多糖化合物中的主要抗氧化成分,TPS几乎没有抗氧化活性[58]。同时,绿茶TPS可以抑制高脂饮食大鼠对脂肪酸的吸收和脂肪细胞因子如TNF-、IL-6的基因表达,降低肝糖异生和糖原分解等活动从而减少甘油三酯的积累[43,55]。
3.4 茶多糖调节肠道菌群
人体肠道中存在大量的微生物,种类繁多,大致可以分为3类:中性菌、有害菌和有益菌。随着分子生物学的发展,肠道菌群与肥胖的关系近年来受到越来越多的关注。一系列研究显示,肠道及其微生物与宿主的生理、营养和免疫等功能密切相关[59-61]。人类肠道生态系统的稳定性和多样性主要与生存环境和饮食习惯有关,且其微生物种群的多样性有20%以上依赖于与饮食习惯相关的环境因素[62-63]。肠道微生态的紊乱和菌群结构的失衡会导致宿主循环系统内毒素的积累,从而诱发机体的慢性炎症,导致肥胖。
诸多研究表明,茶多糖可以调节肠道菌群紊乱,抑制与肥胖相关的有害菌生长,促进有益菌的增殖,维护肠道屏障功能完整性,从而预防和治疗肥胖[26-27,37,53]。肠道是吸收营养和水分的主要器官,是与环境因素接触最大的区域,含有大量的专门免疫细胞,可以协调防御反应,防止或抵消宿主及其免疫系统暴露于不同来源的干扰源[64]。李海珊等[12]研究发现,50 mg·kg-1和100 mg·kg-1的TPS喂养小鼠21 d后显著增强其肠道巨噬细胞吞噬能力,提高结肠内含水量和短链脂肪酸的生成,从而增强机体的免疫调节功能以维护肠道健康。部分研究者进一步探讨了HFD诱导的肥胖等代谢综合征和肠道菌群的关系。李清亮[37]研究表明,黄大茶TPS能调节高脂饮食小鼠的肠道菌群组成,降低厚壁菌门()与拟杆菌门()的比例,增加与多糖代谢相关及抑制肥胖发生的菌群如丁酸弧菌属()、普世菌属()和双歧杆菌属()的含量,降低与肠道菌群失调和肥胖发生的相关菌属如韦荣球菌科()和脱硫弧菌科()的相对丰度,增强肠道上皮细胞的通透性,提高肠道屏障作用。肠道微生物群也可以直接或通过信号分子(包括短链脂肪酸和胆汁酸)影响许多重要代谢过程[65-68]。研究者利用模拟仿生系统发现茶多糖通过调节肠道菌群发酵促进短链脂肪酸产生,其可被机体的肠道吸收并作为肠道上皮细胞和肝脏的主要能源,影响肠道屏障功能和糖异生等代谢,抑制胆固醇的积累和脂肪的生成[37,65]。另外,胆汁酸与肠道菌群之间存在双向关系,一方面胆汁酸在肝脏中由胆固醇产生,在肠道中由肠道菌群代谢;另一方面结肠胆汁酸可以调节微生物的组成,抑制某些微生物的生长[61,69]。
4 研究展望
综上所述,茶多糖可有效调控肥胖,其作用机制主要包括茶多糖减少食物摄取和能量吸收、调节脂质代谢基因的表达和抑制脂肪生成、增强抗氧化防御酶活性和减轻炎症、调节肠道菌群紊乱和维护肠道屏障完整性等多种途径(图1)。尽管TPS显示了良好的抵抗肥胖特性,但与菌类多糖和某些植物多糖相比还有很大差距。目前,全球已有不少菌类多糖的分子量、结构式、空间构像、构效关系、作用靶点都非常清晰,并已经开发出天然药物面市。因此,茶多糖及其在控制体重、抵御肥胖功能方面仍有一系列的问题需要深入探究。
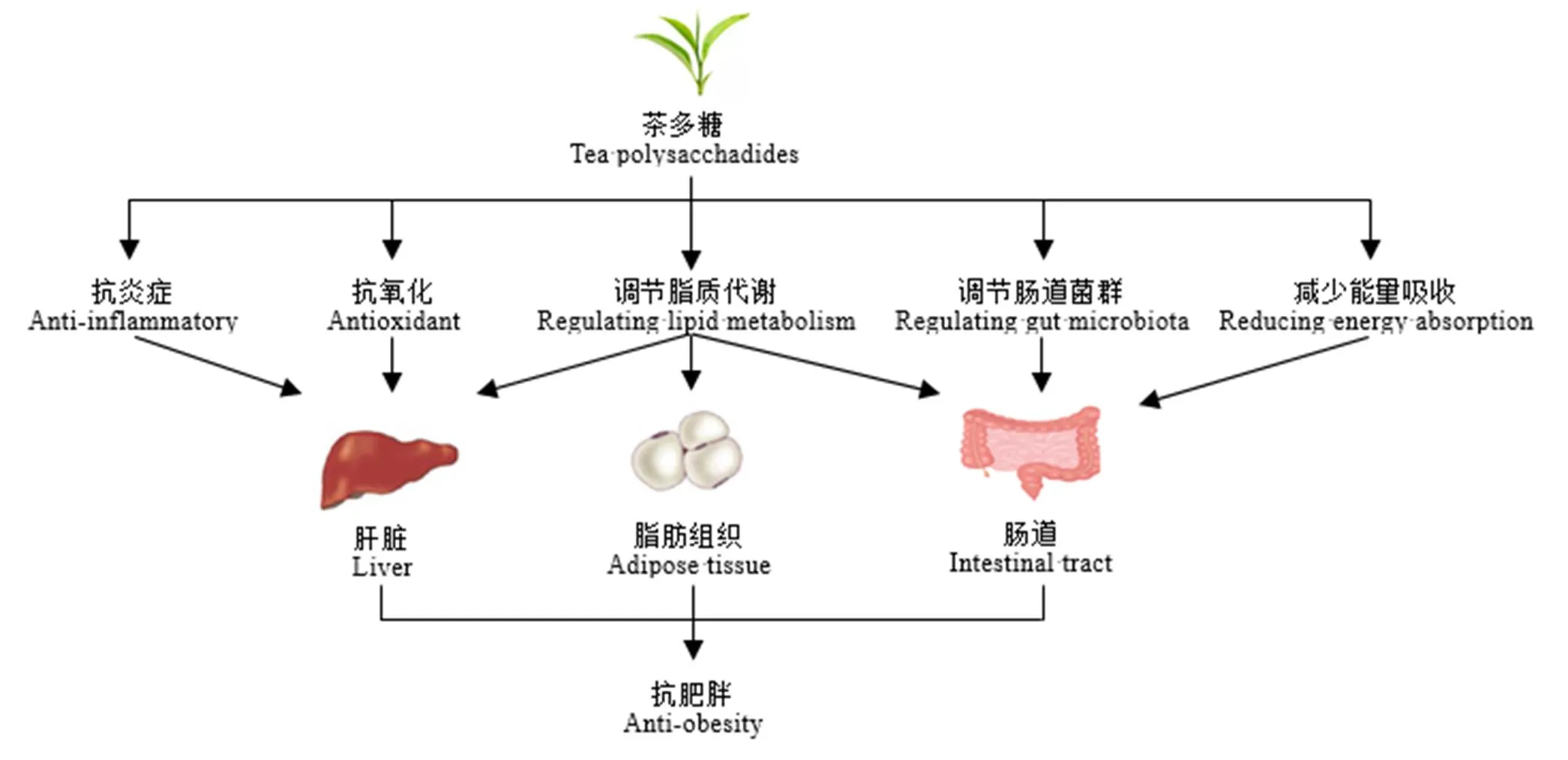
图1 茶多糖调控肥胖机制分析
首先,TPS的化学结构错综复杂,多种生物活性与其化学结构之间的关系仍需进一步研究。分子量大小曾被认为是影响TPS抗氧化活性的重要参数,低分子量TPS组分比高分子量TPS组分具有更高的抗氧化活性[70-72]。然而,分子量与TPS抗氧化活性的关系在许多其他研究中仍没有得到证实[73-74]。近几十年来,研究者对TPS的化学结构和生物活性进行了广泛的探索,但由于结构的复杂性和多样性,其结构特征、溶液行为、空间构象与生物活性之间的关系尚不清楚[75]。
其次,TPS的分离纯化技术与市场对TPS大规模高质量的需求不相适应。目前,已开发的溶剂提取、酶法提取、超声波提取、微波提取及沉淀法、柱色谱法、纳米膜法等茶多糖提取分离方法,还难以满足国内外天然药物和功能食品生产对纯度和规模的要求。因此,需要建立纯度高、分子量与空间结构稳定、构效关系明晰的茶多糖分离纯化技术体系,以满足规模化和标准化生产。
再次,TPS调控肥胖是其自身还是体内代谢物发挥作用仍不明确。目前的体内外试验表明,TPS可以调节高脂食物诱导大鼠的肠道菌群结构,但基于斯皮尔曼相关分析很难证明这些关联的因果关系。因此,有必要应用无菌动物和粪便移植进一步研究调节肠道菌群失调和代谢物的关系,从而作为预防和治疗肥胖等代谢性疾病的新靶点[9]。
最后,TPS调控肥胖的有效剂量及其临床效果亟待研究。目前TPS调节肥胖的研究主要集中在细胞和动物领域,且以黑茶TPS为主,而在人体临床及流行病学方面的研究几乎空白。动物和人体在有效剂量和吸收代谢方面差异较大,体外试验、动物试验与人体临床的效果还存在差距。
因此,TPS调控机体肥胖的有效剂量、作用机制及体内代谢过程均有待系统深入研究,此外,还需开展人体流行病学研究。
[1] Breda J, Jewell J, Keller A. The importance of the World Health Organization sugar guidelines for dental health and obesity prevention [J]. Caries Research, 2019, 53: 149-152.
[2] Silvia B S, Ana O R, Mirjam M H, et al. Clustering of multiple energy balance-related behaviors in school children and its association with overweight and obesity: WHO European Childhood Obesity Surveillance Initiative (COSI 2015-2017) [J]. Nutrients, 2019, 11(3): 511. doi: 10.3390/nu11030511.
[3] Ezzati M, Bentham J, Di C M, et al. Worldwide trends in body-mass index, underweight, overweight, and obesity from 1975 to 2016: a pooled analysis of 2416 population-based measurement studies in 128.9 million children, adolescents, and adults [J]. Lancet, 2017, 390(10113): 2627-2642.
[4] 马冠生. 中国儿童肥胖报告[R]. 北京: 人民卫生出版社, 2017. Ma G S. Report on childhood obesity in china [R]. Beijing: People's Medical Publishing House, 2017.
[5] Guida S, Venema K. Gut microbiota and obesity: involvement of the adipose tissue [J]. Journal of Functional Foods, 2015, 14: 407-423.
[6] Guh D P, Zhang W, Bansback N, et al. The incidence of co-morbidities related to obesity and overweight: a systematic review and meta-analysis [J]. BMC Public Health, 2009, 9: 88. doi: 10.1186/1471-2458-9-88.
[7] Piche M E, Poirier P, Lemieux I, et al. Overview of epidemiology and contribution of obesity and body fat distribution to cardiovascular disease: an update [J]. Progress Cardiovasc Diseases, 2018, 61(2): 103-113.
[8] Lee E Y, Yook K H. Epidemic obesity in children and adolescents: risk factors and prevention [J]. Front Med, 2018, 12(6): 658-666.
[9] Chen G, Chen R, Chen D, et al. Tea polysaccharides as potential therapeutic options for metabolic diseases [J]. Journal of Agricultural and Food Chemistry, 2018, 67(19): 5350-5360.
[10] Guirro M, Herrero P, Costa A, et al. Comparison of metaproteomics workflows for deciphering the functions of gut microbiota in an animal model of obesity [J]. Journal of Proteomics, 2019, 209: 103489. doi: 10.1016/j.jprot.2019.103489.
[11] Song D, Cheng L, Zhang X, et al. The modulatory effect and the mechanism of flavonoids on obesity [J]. Journal of Food Biochemistry, 2019, 43(8): e12954. doi: 10.1111/jfbc.12954.
[12] 李海珊, 刘丽乔, 聂少平. 茶多糖对小鼠肠道健康及免疫调节功能的影响[J]. 食品科学, 2017, 38(7): 187-192.Li H S, Liu L Q, Nie S P. Effects of green tea polysaccharides on intestinal health and immune regulation in mice [J]. Food Science, 2017, 38(7): 187-192.
[13] Yang C S, Zhang J, Zhang L, et al. Mechanisms of body weight reduction and metabolic syndrome alleviation by tea [J]. Molecular Nutrition & Food Research, 2016, 60(1): 160-174.
[14] Wu T, Guo Y, Liu R, et al. Black tea polyphenols and polysaccharides improve body composition, increase fecal fatty acid, and regulate fat metabolism in high-fat diet-induced obese rats [J]. Food Function, 2016, 7(5): 2469-2478.
[15] Nakamura M, Miura S, Takagaki A, et al. Hypolipidemic effects of crude green tea polysaccharide on rats, and structural features of tea polysaccharides isolated from the crude polysaccharide [J]. International Journal Food Science Nutrients, 2017, 68(3): 321-330.
[16] Li S, Chen H, Wang J, et al. Involvement of the PI3K/Akt signal pathway in the hypoglycemic effects of tea polysaccharides on diabetic mice [J]. International Journal Biological Macromolecules, 2015, 81: 967-974.
[17] Chen H, Wang Z, Qu Z, et al. Physicochemical characterization and antioxidant activity of a polysaccharide isolated from oolong tea [J]. European Food Research and Technology, 2009, 229(4): 629-635.
[18] 艾于杰. 抗氧化活性茶多糖构效关系研究[D]. 武汉: 华中农业大学, 2019. Ai Y J. Study on the structure-activity relationship of antioxidant polysaccharides [D]. Wuhan: Huazhong Agricultural University, 2019.
[19] 邵淑宏. 乌龙茶多糖理化性质及抗氣化、降血糖活性研究[D]. 杭州: 浙江大学, 2015. Shao S H. Study on physical and chemical properties of dragon tea polysaccharide and its anti-gasification and hypoglycemic activity [D]. Hangzhou: Zhejiang University, 2015.
[20] Chen H, Zhang M, Qu Z, et al. Antioxidant activities of different fractions of polysaccharide conjugates from green tea () [J]. Food Chemistry, 2008, 106(2): 559-563.
[21] 李娟, 刘锐, 吴涛, 等.不同茶多糖对3T3-L1前脂肪细胞分化的抑制作用比较[J]. 食品科学, 2017, 38(21): 187-193. Li J, Liu R, Wu T, et al. Comparison of the inhibitory effects of different tea polysaccharides on the differentiation of 3T3-L1 preadipocytes [J]. Food Science, 2017, 38(21): 187-193.
[22] 刘海燕, 任发政, 李景明, 等. 几种植物多糖的结构特征与预防肥胖活性研究[J]. 中国食物与营养, 2019, 25(12): 44-51. Liu H Y, Ren F Z, Li J M, et al. Structural characteristics and anti-obesity efficacy of several plant polysaccharides [J]. Food and Nutrition in China, 2019, 25(12): 44-51.
[23] Chen G, Wang M, Xie M, et al. Evaluation of chemical property, cytotoxicity and antioxidant activityandof polysaccharides from Fuzhuan brick teas [J]. International Journal Biological Macromolecules, 2018, 116: 120-127.
[24] Chen H X, Zhang M, Qu Z S. Compositional analysis and preliminary toxicological evaluation of a tea polysaccharide conjugate [J]. Journal of Agricultural and Food Chemistry, 2007, 55: 2256-2260.
[25] 韦铮, 贺燕, 郝麒麟, 等. 茶多糖在模拟胃肠消化体系的抗氧化作用研究[J]. 食品与发酵工业, 2020, 46(10): 109-117.Wei Z, He Y, Hao Q L, et al. Study on the antioxidant effect of tea polysaccharides under the conditions of simulating gastrointestinal digestion[J]. Food and Fermentation Industries, 2020, 46(10): 109-117.
[26] Chen G J, Xie M H, Wan P, et al. Digestion under saliva, simulated gastric and small intestinal conditions and fermentation in vitro by human intestinal microbiota of polysaccharides from Fuzhuan brick tea [J]. Food Chemistry, 2018, 244: 331-339.
[27] Li W W, Wang C, Yuan G Q, et al. Physicochemical characterisation and α-amylase inhibitory activity of tea polysaccharides under simulated salivary, gastric and intestinal conditions [J]. International Journal of Food Science & Technology, 2018, 53(2): 423-429.
[28] Fernández J, Redondo-B S, Gutiérrez-del R I, et al. Colon microbiota fermentation of dietary prebiotics towards short-chain fatty acids and their roles as anti-inflammatory and antitumour agents: A review [J]. Journal of Functional Foods, 2016, 25: 511-522.
[29] 张高帆, 陈萍, 徐思绮, 等. 茶多糖对四氧嘧啶模型小鼠的降糖作用及其体内分布规律研究[J]. 营养学报, 2015, 37(4): 384-388. Zhang G F, Chen P, Xu S Q, et al. Hypoglycemic effect of tea polysaccharide on alloxan model mice and its in vivo research on distribution law [J]. Journal of Nutrition, 2015, 37(4): 384-388.
[30] Sánchez J, Priego T, Palou M, et al. Oral supplementation with physiological doses of leptin during lactation in rats improves insulin sensitivity and affects food preferences later in life [J]. Endocrinology, 2008, 149(2): 733-740.
[31] 杨志秋, 詹莉莉, 傅正伟. 脂肪酶抑制剂应用于抗肥胖的研究进展[J]. 现代生物医学进展, 2011, 11(21): 4178-4181. Yang Z Q, Zhan L L, Fu Z W. Recent advances of lipase inhibitor in the application of anti-obesity [J]. Progress in Modern Biomedicine, 2011, 11(21): 4178-4181.
[32] 李祥龙, 李晓梅, 杨煦, 等. 黑茶茶褐素与茶多糖对脂肪酶的抑制作用[J]. 食品与机械, 2018, 34(3): 27-31, 58. Li X L, Li X M, Yang X, et al. Study of inhibition of black tea theabrownin and tea polysaccharides on lipase [J]. Food and Machinery, 2018, 34(3): 27-31, 58.
[33] 舒婷, 肖畅, 何慧, 等. 青砖茶粗多糖抑制-葡萄糖苷酶活性的研究[J]. 食品科技, 2019, 44(3): 194-199. Shu T, Xiao C, He H, et al. Inhibitory effects of crude polysaccharide of green brick tea on-glucosidase activity [J]. Food Science and Technology, 2019, 44(3): 194-199.
[34] 和兴萍, 罗燕, 李雪, 等. 几种降脂减肥实验动物模型的建立与比较[J]. 中华中医药学刊, 2017, 35: 1747-1751. He X P, Luo Y, Li X, et al. Comparison of several kinds of lipid-Lowering diet experimental animal model [J]. Chinese Journal of Traditional Chinese Medicine, 2017, 35: 1747-1751.
[35] 陈粉粉, 郭爱伟, 周杰珑, 等. ICR小鼠肥胖模型的建立以及肥胖指标和脂肪组织形态学比较[J]. 安徽农业科学, 2012, 40(5): 2720-2073. Chen F F, Guo A W, Zhou J L, et al. Establishment of the obesity model of ICR mice and the comparison of the obesity index and morphology of adipose tissue [J]. Journal of Anhui Agri, 2012, 40(5): 2720-2073.
[36] Ren D, Hu Y, Luo Y, et al. Selenium-containing polysaccharides from Ziyang green tea ameliorate high-fructose diet induced insulin resistance and hepatic oxidative stress in mice [J]. Food Function, 2015, 6(10): 3342-3350.
[37] 李清亮. 黄大茶茶多糖对饲喂高脂日粮小鼠肠道菌群的调节作用[D]. 合肥: 安徽农业大学, 2018. Li Q L. Polysacchardies in Large Yellow tea modulate gut microbiome in HFD fed mice [D]. Hefei: Anhui Agricultural University, 2018.
[38] Mao Y, Wei B, Teng J, et al. Polysaccharides from Chinese Liupao dark tea and their protective effect against hyperlipidemia [J]. International Journal of Food Science & Technology, 2018, 53(3): 599-607.
[39] 郭郁, 吴涛, 刘锐, 等. 红茶提取物减肥作用研究[J]. 现代食品科技, 2017, 33(2): 16-21. Guo Y, Wu T, Liu R, et al. Study on the weight-loss effect of black tea extracts [J]. Modern Food Science and Technology, 2017, 33(2): 16-21.
[40] 吴文华. 洱茶调节血脂功能评价及其生化机理的研究[D] . 重庆: 西南农业大学, 2003. Wu W H. Functional appraisal of blood lipid adjusted by Puer tea and study on its physiochemical mechanism [D]. Chongqing: Southwest Agricultural University, 2003.
[41] Olsen M K, Johannessen H, Cassie N, et al. Steady-state energy balance in animal models of obesity and weight loss [J]. Scandinavian Journal Gastroenterology, 2017, 52(4): 442-449.
[42] Ahima R S, Antwi D A. Brain regulation of appetite and satiety [J]. Endocrinology and Metabolism Clinics of North America, 2008, 37(4): 811-823.
[43] Xu Y, Zhang M, Wu T, et al. The anti-obesity effect of green tea polysaccharides, polyphenols and caffeine in rats fed with a high-fat diet [J]. Food Function, 2015, 6(1): 297-304.
[44] Maniadakis N, Kapaki V, Damianidi L, et al. A systematic review of the effectiveness of taxes on nonalcoholic beverages and high-in-fat foods as a means to prevent obesity trends [J]. ClinicoEconomics and Outcomes Research, 2013, 5: 519-543.
[45] Huang J, Wang Y, Xie Z, et al. The anti-obesity effects of green tea in human intervention and basic molecular studies [J]. European Journal Clinical Nutrition, 2014, 68(10): 1075-1087.
[46] Chung J O, Yoo S H, Lee Y E, et al. Hypoglycemic potential of whole green tea: water-soluble green tea polysaccharides combined with green tea extract delays digestibility and intestinal glucose transport of rice starch [J]. Food & Function, 2019, 10(2): 746-753.
[47] Saely C H, Geiger K, Drexel H. Brown versus white adipose tissue: a mini-review [J]. Gerontology, 2012, 58(1): 15-23.
[48] Frühbeck G, Becerril S, Sáinz N, et al. BAT: A new target for human obesity? [J]. Trends in Pharmacological Sciences, 2009, 30(8): 387-396.
[49] Ferranti S, Mozaffarian D. The perfect storm: obesity, adipocyte dysfunction, and metabolic consequences [J]. Clinical Chemistry, 2008, 54(6): 945-955.
[50] Hammad S S, Eck P, Sihag J, et al. Common variants in lipid metabolism-related genes associate with fat mass changes in response to dietary monounsaturated fatty acids in adults with abdominal obesity [J]. The Journal of Nutrition, 2019, 149(10): 1749-1756.
[51] Bazhan N M, Baklanov A V, Piskunova J V, et al. Expression of genes involved in carbohydrate-lipid metabolism in muscle and fat tissues in the initial stage of adult-age obesity in fed and fasted mice [J]. Physiological Reports, 2017, 5(19): e13445. doi: 10.14814/phy2.13445.
[52] Catalán V, Rodríguez A, Ramírez B, et al. Association of increased Visfatin/PBEF/NAMPT circulating concentrations and gene expression levels in peripheral blood cells with lipid metabolism and fatty liver in human morbid obesity [J]. Nutrition Metabolism & Cardiovascular Diseases, 2017, 21(4): 245-253.
[53] Chen G J, Xie M H, Wan P, et al. Fuzhuan brick tea polysaccharides attenuate metabolic syndrome in high-fat diet induced mice in association with modulation in the gut microbiota [J]. Journal of Agricultural and Food Chemistry, 2018, 66(11): 2783-2795.
[54] Manna P, Jain S K. Obesity, oxidative stress, adipose tissue dysfunction, and the associated health risks: causes and therapeutic strategies [J]. Metabolic Syndrome And Related Disorders, 2015, 13(10): 423-444.
[55] Fernandez S A, Madrigal S E, Bautista M, et al. Inflammation, oxidative stress, and obesity [J]. International Journal of Molecular Sciences, 2011, 12(5): 3117-3132.
[56] Savini I, Catani M V, Evangelista D, et al. Obesity-associated oxidative stress: strategies finalized to improve redox state [J]. International Journal of Molecular Sciences, 2013, 14(5): 10497-10538.
[57] Wang J, Liu W, Chen Z, et al. Physicochemical characterization of the oolong tea polysaccharides with high molecular weight and their synergistic effects in combination with polyphenols on hepatocellular carcinoma [J]. Biomed Pharmacother, 2017, 90: 160-170.
[58] Wang Y, Zhao Y, Andrae M K, et al. Tea polysaccharides as food antioxidants: an old woman's tale? [J]. Food Chemistry, 2013, 138(2/3): 1923-1927.
[59] Zhang L, Gui S, Liang Z, et al.cecropin (Mdc) alleviates-induced colonic mucosal barrier impairment: associating with inflammatory and oxidative stress response, tight junction as well as intestinal flora [J]. Frontiers in Microbiology, 2019, 10: 522. doi: 10.3389/fmicb.2019.00522.
[60] Musso G, Gambino R, Cassader M. Interactions between gut microbiota and host metabolism predisposing to obesity and diabetes [J]. The Annual Review of Medicine, 2011, 62: 361-380.
[61] Huang F, Zheng X, Ma X, et al. Theabrownin from Pu-erh tea attenuates hypercholesterolemia via modulation of gut microbiota and bile acid metabolism [J]. Nature Communications, 2019, 10(1): 4971. doi: 10.1038/s41467-019 -12896-x.
[62] Rothschild D, Weissbrod O, Barkan E, et al. Environment dominates over host genetics in shaping human gutmicrobiota [J]. Nature, 2018, 555: 210-215.
[63] Rosenbaum M, Knight R, Leibel R L. The gut microbiota in human energy homeostasis and obesity [J]. Trends Endocrinol Metab, 2015, 26(9): 493-501.
[64] Bernardi S, Del Bo C, Marino M, et al. Polyphenols and intestinal permeability: rationale and future perspectives [J]. Journal of Agricultural and Food Chemistry, 2020, 68(7): 1816-1829.
[65] Dugas L R, Lie L, Plange-Rhule J, et al. Gut microbiota, short chain fatty acids, and obesity across the epidemiologic transition: the METS-Microbiome study protocol [J]. BMC Public Health, 2018, 18(1): 978. doi: 10.1186/s12889-018 -5879-6.
[66] Tremaroli V, Backhed F. Functional interactions between the gut microbiota and host metabolism [J]. Nature, 2012, 489(7415): 242-249.
[67] Petriz B A, Castro A P, Almeida J A, et al. Exercise induction of gut microbiota modifications in obese, non-obese and hypertensive rats [J]. BMC Genomics, 2014, 15: 511. doi: 10.1186/1471-2164-15-511.
[68] Queipo M I, Seoane L M, Murri M, et al. Gut microbiota composition in male rat models under different nutritional status and physical activity and its association with serum leptin and ghrelin levels [J]. Plos One, 2013, 8(5): e65465. doi: 10.1371/journal.pone.0065465.
[69] Wahlström A, Sayin S I, Marschall H U, et al. Intestinal crosstalk between bile acids and microbiota and its impact on host metabolism [J]. Cell Metabolism, 2016, 24(1): 41-50.
[70] Chen H, Qu, Z, Fu L L, et al. Physicochemical properties and antioxidant capacity of 3 polysaccharides from green tea, oolong tea, and black tea [J]. Journal of Food Science, 2009, 74(6): 469474. doi: 10.1111/j.1750-3841.2009.01231.x.
[71] Chen H X, Zhang M, Qu Z S, et al. Antioxidant activities of different fractions of polysaccharide conjugates from green tea () [J]. Food Chemistry, 2008, 106: 559-563.
[72] Du L L, Fu Q Y, Xiang L P, et al. Tea polysaccharides and their bioactivities [J]. Molecules, 2016, 21(11): 1449. doi: 10.3390/molecules21111449.
[73] Wang Y F, Liu Y Y, Huo J L, et al. Effect of different drying methods on chemical composition and bioactivity of tea polysaccharides [J]. International Journal Biological Macromolecules, 2013, 62: 714-719.
[74] Zhang X, Chen H X, Zhang N, et al. Extrusion treatment for improved physicochemical and antioxidant properties of high-molecular weight polysaccharides isolated from coarse tea [J]. Food Research International, 2013, 53: 726-731.
[75] Chen G, Yuan Q, Saeeduddin M, et al. Recent advances in tea polysaccharides: extraction, purification, physicochemical characterization and bioactivities [J]. Carbohydrate Polymers, 2016, 153: 663-678.
ResearchProgress of Tea Polysaccharides in Regulating Obesity
OUYANG Jian1,2, ZHOU Fang1,2, LU Danmin1,2, LI Xiuping1,2, HUANG Jian'an1,2,3, LIU Zhonghua1,2,3*
1. National Research Center of Engineering and Technology for Utilization of Botanical Functional Ingredients, Changsha 410128, China; 2. Key Lab of Education Ministry of Hunan Agricultural University for Tea Science, Changsha 410128, China; 3.Co-Innovation Center of Education Ministry for Utilization of Botanical Functional Ingredients, Changsha 410128, China
With the improvement of people's living standards, the incidence of obesity has been rising, which has become a serious health problem of the society. Tea polysaccharide, as an acid heteropolysaccharide combined with protein, can regulate food intake and energy absorption, regulate adipogenesis, enhance antioxidant defense enzyme activities and reduce inflammation, regulate intestinal flora disorders and maintain different pathways such as intestinal barrier integrity, thereby effectively regulates obesity. The regulation mechanism of tea polysaccharides on obesity was reviewed based on the researches in recent years.
tea polysaccharide, obesity, regulation, mechanism of action
S571.1;Q946.3
A
1000-369X(2020)05-565-11
2020-03-16
2020-05-17
国家重点研发计划(2017YFD0400803)、国家自然科学基金(31801574)、国家茶叶产业技术体系研究项目(CARS-19-C01)
欧阳建,男,硕士研究生,主要从事茶叶深加工与功能成分利用方面的研究。*通信作者:larkin-liu@163.com
