棉田间套复合体系提升生态系统服务功能研究进展
2020-10-10李小飞韩迎春王国平王占彪冯璐杨北方范正义雷亚平熊世武邢芳芳李亚兵
李小飞,韩迎春,王国平,王占彪,冯璐,杨北方,范正义,雷亚平,熊世武,邢芳芳,李亚兵
(中国农业科学院棉花研究所/ 棉花生物学国家重点实验室,河南安阳455000)
棉花是我国重要的经济作物,在整个国民经济中占有非常重要的地位,除西藏、青海、黑龙江三省外均有种植。 据联合国粮农组织数据库资料显示, 我国棉花常年种植面积约400 万公顷,平均籽棉产量为3 800 kg·hm-2[1]。 目前中国棉花的单产和总产居于世界第一位,新疆棉花单产及总产在中国远远高于全国平均水平,2016 年新疆棉区单产每公顷高达1 991 kg, 总产达到359 万t,占全国棉花总产量的67%[2]。 但是近十年来由于对粮食作物的重视、蔬菜种植面积的扩大,我国棉花种植面积呈现出逐年下降的趋势。 例如,2007 年我国植棉面积为593 万hm2,但截止2016年,我国植棉面积下降到338 万hm2[2]。 棉花与粮食作物及其他作物之间的争地矛盾已成为制约棉花生产的重要因素之一。 依靠两熟和多熟种植可以实现在有限的耕地上同时满足粮、棉、油、菜、饲料等多种农产品的需求[3]。棉田间套复合种植体系不仅能有效地缓解粮棉、经棉和果棉争地矛盾,同步实现棉花与其他作物增产增收[4],而且还可以作为协调作物产量与生物多样性保护二者之间矛盾的有效手段[5]。 因此,在农业绿色可持续发展研究中越来越受到人们的重视和应用[6]。
1 棉田间套复合体系发展现状与趋势预测
早在20 世纪60―70 年代, 我国棉田就由一熟制改革调整为麦棉、油棉和豆棉两熟制。20世纪80 年代中后期, 在巩固两熟制的基础上,基本实现了粮菜棉、粮饲棉等三熟制。 21 世纪初, 全国棉田两熟和多熟种植面积占总棉田面积的2/3,复种指数达到156%。 近年来,为了实现棉花轻简化和机械化生产, 我国棉田间套复合种植所占的比例稳中有降,复种指数降低[7]。随着农业结构的调整, 棉田间套复合体系也表现出了多元性和多样性, 已形成稳定的种植制度。总体来看,棉田套作模式已经由过去的麦棉套作改为棉花与多种作物套作, 归纳起来有四大作物搭配类型,即棉菜型、棉果型、棉药型、棉油粮(饲)型[7]。
虽然目前我国还没有间作套种面积的统计数据,但从一些报道也可以说明间作套种在我国现代农业中仍然占有一定地位[8]。据不完全统计,我国常年间(套)作种植面积约2 800 万hm2[9]。基于农户尺度上的调查数据显示,较2009 年相比,2014 年我国间(套)作和农林复合系统的种植面积并没有呈现大幅度降低的趋势[10]。 棉田一年一熟制为主的种植制度已初现弊端, 棉花/ 绿肥套作可以很好地解决冬季地表裸漏问题,降低养分淋溶,提高肥力,具有巨大的应用前景[11]。 中国西南部旱地面积比率高,复种程度高,是典型的旱作多熟农业区,间(套)作年种植面积达358万hm2,对该地区粮食增产起到了重要作用[12]。此外, 国家政策也大力支持和推广轮作和间作套作,支持因地制宜开展生态型复合种植,科学合理利用耕地资源,促进种地养地结合。 因此,绿色生态可持续的大背景下,间作套种在现代农业中发挥着不可替代的作用。
2 棉田间套复合体系提升生态系统服务功能
生态系统服务功能是指生态系统形成和所维持的人类赖以生存和发展的环境条件与效用,是人类赖以生存的基础。 可归纳为提供产品功能、调节功能、文化功能和支持功能[13]。 棉田间套复合种植能够提升生态系统服务功能,如增加生物多样性、 系统生产力和经济效益,提高光热利用效率,改善作物品质和控制病虫草害等(表1)。
2.1 增加生物多样性
农田生态系统作为人类干扰最为严重的生态系统类型。 在过去的几十年里,世界范围内的生物多样性加速降低,农业活动是造成生物多样性降低的1 个重要驱动因素,通过栽培措施保持农田生态系统生物多样性面临着巨大的挑战[13]。在棉田间套复合种植体系中,不同种类作物的生物学特性以及各个生育期内对环境的影响也不尽相同,无论是从外部表现、内部冠层结构,还是根系在土壤空间的分布上,彻底改变了单作棉田生物结构单一的问题,因此增加了作物时间和空间上的多样性。 同时,地上部多样性与地下部多样性存在紧密联系,多样性种植体系势必会对地下部多样性产生影响,包括根系化学成分、分泌物组成和微生物群落结构等[14]。 在砂培条件下研究3∶1 式麦棉套作棉花根系分泌物对土壤微生物的影响,结果表明棉花根系分泌物可以改变微生物群落结构,细菌和真菌的数量分别比对照高49.8%和52.9%, 增加地下部微生物的多样性[15]。微生物群落组成的变化与土壤氮素矿化速率和氮有效性密切相关[16]。

表1 棉田间套复合种植强化农田生态系统服务功能Table 1 Cotton-based intercropping systems enhance agroecosystem services and functioning
2.2 增加系统生产力和经济效益
近来的研究表明,与单一作物的连续种植相比,棉田间套复合体系具有生产力优势[17-18]。 土地当量比(Land equivalent ratio, LER)常被用来评价间(套)作体系是否具有间作优势[19],定义为获得与间作相同产量时所需要的单作的土地面积。当LER >1,表明间作具有生产力优势[20]。在黄河流域地区,不同行比配置模式的麦/ 棉套作LER为1.28~1.39, 地上部氮素累积量比单作高出40%~70%,表现出了明显的间作优势[17,21]。 Feng等[22]对比不同棉花种植模式下的投入量和产出量,结果表明,麦棉两熟种植能够增加系统周年经济效益10.9%~74.4%。 蒜套春棉由于具有较高的经济效益, 因此在鲁西南棉区被普遍采用[23-24]。 在长江流域地区,与单作棉花相比,麦/棉套作、麦后移栽棉和麦后直播棉平均小麦和棉花周年经济产量 (小麦籽粒产量与籽棉产量)增产120.2%~162.6%[25]。 党小燕等[26]研究新疆地区不同作物与棉花间作对土地当量比的影响,表明棉花与鹰嘴豆、花生、洋葱、萝卜间作后LER 分别为1.02、1.30、1.12、1.68,均大于1。此外,国际上也有相关的报道。印度棉花/洋葱间作时棉花的等价产量(Cotton equivalent yield)为1 974 kg·hm-2,显著高于单作棉花的1 779 kg·hm-2[27]。 Jayakumar 和Surendran[28]也得出相似的研究结果:棉花/ 洋葱和棉花/ 黑豆间作时棉花等价产量分别比单作棉花高出21.2%和13.3%。 在非洲,1∶1 和1∶2行比模式豇豆/ 棉花间作体系的LER 分别为1.4 和1.3[29]。 因此,合理的棉花间(套)作LER均大于1,表明棉花间(套)作种植能够提升农田生态系统生产功能。然而,不合理的间(套)作也会造成共生作物对资源的竞争加剧, 从而降低作物单位面积的物质产出[30]。合理的时间、空间和物种搭配是棉田间套复合体系增加生产力的前提。
2.3 提高光热利用效率
光合有效辐射PAR(Photosynthetically active radiation) 是指太阳辐射中能被作物吸收并用来进行光合作用的那部分能量。 作物群体光能利用率与群体冠层结构密切相关,直接影响其光合特性,进而影响生物学产量。 不同类型作物辐射利用效率存在显著差异,不同类型作物搭配组合可构建合理的冠层结构,改善冠层辐射传输,提高群体光能利用率[31]。Du 等[32]研究不同麦棉种植模式对冠层光能截获和利用效率的影响,结果表明麦棉两熟种植能够改善棉花的冠层结构,提高冠层内光的穿透性,群体的光合能力增加,光热利用效率比单作棉增加,其中泗杂3 号和中棉所50两个品种单作种植,棉花的光热利用效率分别为1.20 和1.14 g·MJ-1,而套作棉、麦后移栽棉和麦后直播棉的变幅为1.19~1.93 g·MJ-1,高于相应的单作棉花。 较单作小麦和单作棉花,麦棉两熟种植具有高的光截获能力一方面是小麦作物充分利用了冬季和春季的光合有效辐射,而在单作棉花中这部分光合有效辐射极易被浪费掉,另一方面是两熟种植中小麦作物的交错设计更有利于光的截获[33]。 然而,也有研究表明,间(套)作增加了系统的累积光截获量,但光利用效率很少会受到种植方式的影响[33]。
2.4 改善棉花品质
棉花纤维品质主要由基因型决定,同时也受栽培措施和环境条件的影响[4]。 衡量和评价棉花纤维品质的指标主要包括上半部平均长度、断裂比强度、马克隆值、整齐度和伸长率等。 在麦棉套作体系中,Zhang 等[17]研究发现,3∶1、3∶2 和4∶2 行比配置模式的麦棉套作下棉花的马克隆值显著高于单作棉花, 但单作棉花纤维长度、强度和整齐度指数与间作处理没有显著差异。 辽河流域棉花/ 花生间作可使棉花吐絮时间提前且集中,间作棉花纤维上半部平均长度增加0.73 mm,可纺性指数高6.67,断裂比强度高0.72 cN·tex-1[34]。此外,在新疆对不同作物与棉花间作种植下棉花纤维品质的研究也表明, 与单作棉花相比,棉花与洋葱间作能显著地提高棉花纤维上半部平均长度;棉花与线辣椒、鹰嘴豆、花生间作,纤维马克隆值显著地高于单作,但间作对棉花纤维的断裂比强度、整齐度指数、伸长率、反射率、纺纱均匀性指数没有显著影响[35]。
2.5 控制病虫害
不同遗传背景、农艺性状和经济性状的作物种植在一起,由于种间相互作用的增强,可改善作物营养状况,增强作物的抗性,或降低植物病原菌孢子的扩散,最终降低病害大面积发生的风险[36]。 例如棉花/田箐(Sesbania aculeate L.)间作处理的荧光假单胞菌侵染率最高,但棉花根腐病病情指数最低。 与单作相比,间作棉花营养生长、开花和成熟时期根腐病病情指数分别降低0.61、1.06 和1.17 百分点[37]。 Zhang 等[38]研究棉田边缘苜蓿防护林带控制棉蚜的效果,结果表明棉田周边种植苜蓿(Medicago sativa L.)对棉蚜有很好的控制效果,在棉蚜进入棉田的始期,苜蓿带中已经繁育了大量的天敌,总量达到棉田的6.9 倍。棉蚜进入棉田后的数量激增阶段,苜蓿带中棉蚜的主要天敌类群(瓢虫类)总数量达到棉田的13.6倍。 棉花与苜蓿接壤种植时,棉花蚜虫种群增长速率明显低于未与苜蓿接壤种植的棉花,而棉花蚜虫的天敌如蜘蛛、草蛉种群的增长速率则表现出相反的趋势,表明选用苜蓿作为棉花的间作或邻作作物,对调整捕食性节肢动物类天敌并利用其进行害虫生物防治具有重要意义[39]。 Men 等[40]通过2 年的田间试验也研究表明,麦棉两熟套作种植时,小麦种植带可以作为棉蚜天敌的“中转站”。 小麦收获后,这些棉蚜天敌迁移进入相邻的棉花作物,有效抑制了棉田蚜虫的大爆发。
2.6 抑制杂草生长
控制杂草也是棉田间套复合种植的另1 个重要生态效应。 在养分资源和光热利用上,如果物种搭配组合中存在对杂草具有较强竞争能力的作物时,多作物的复合种植就可以降低杂草发生率[41]。 Iqbal 等[42]通过2 年的田间试验研究表明,棉花与高粱、大豆和芝麻间作后,杂草香附子(Cyperus rotundus L.) 的密度比单作棉田降低70%~96%,生物量降低71%~97%,增加棉田总收益20%~60%。 大豆-棉花轮作种植也是高效控制香附子的有效途径之一[43]。
3 棉田间套复合体系提升生态系统服务功能的潜在机制
大量研究一致表明,棉田间(套)作复合种植体系能够增加农田生态系统物质产出、提高资源利用效率、控制病虫草害发生等。 这些强化的农田生态系统服务功能的发生必然与种间物种相互作用密切相关,包括(1)种间根系时空分布的异质性;(2) 种间促进作用;(3)化感作用(图1)。
3.1 生态位互补
生态位反映了物种在其所处的环境中的资源状态。 棉田复合种植借助高秆和矮秆、禾本科和豆科、窄叶和宽叶、喜荫和耐荫等作物的配置,充分发挥不同作物之间的形态互补、 生育期互补、生态互补和边缘优势,从而提高单位面积产量和光合效率[44]。麦棉套作共生期,小麦处于生殖生长时期,而棉花处于苗期,两作物一高一矮搭配,分别占据不同的冠层生态位,造成单位面积间作累积截获的光能显著高于单作[33]。
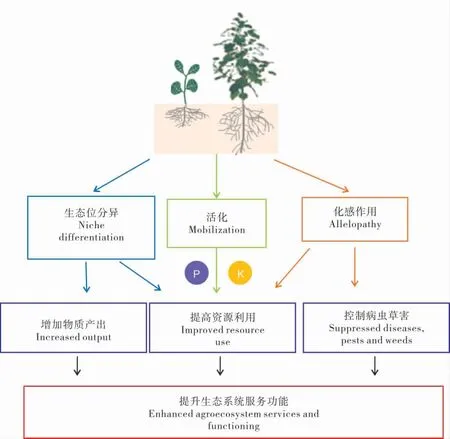
图1 棉田间套复合体系提升生态系统服务功能的潜在机制Fig. 1 Potential mechanisms for enhanced ecosystem services and functioning by cotton-based intercropping systems
棉田间套复合群体不仅存在地上部冠层光热资源的互补, 而且地下部存在根系系统的互补。 采用剖面法分层取样,并结合图像扫描分析,研究枣/ 棉间作模式对棉花根系分布的影响,表明单作棉花根系主要分布在水平0~20 cm、垂直0~20 cm 范围内;间作棉花根系主要分布在水平0~40 cm、垂直0~40 cm 范围内。 间作棉花单株根长密度总量较单作高1.8%,总根质量较单作高15.5%,间作促进棉花根系分布更广、更深,比单作棉花根系具有更大空间的生态位[45],最终增加了根系吸收面积。 当养分资源受限时,间作作物仍然能够获得高产所需的养分和水分资源。 王立国等[46]研究表明,盛花期后,套作棉浅层根系干物质质量在盛花期前低于单作棉,深层根系干物质质量显著高于单作棉, 根系总量也超过单作棉。麦棉共生期间自然套作群体中棉花根表面积、根长度和根干物质质量大于单作和隔根处理,小麦收获后同一生育期纱网隔根处理的棉花根表面积、根长度和根干物质质量大于单作,后期自然套作根系的衰退较其他处理慢[47]。 在生育期内土壤速效磷的时空分布特征表明麦棉两熟种植能够改变上层土壤速效磷的时空分布,这与作物根系分布的差异有关[48]。
3.2 种间促进作用
种间促进作用是指1 种作物促进另1 种作物生长或生存的现象[49]。在多样性种植体系中,最明显的是1 种作物通过根际过程改善资源有效性对邻体植物生长的改善作用[5]。 棉田间套复合体系提高土壤养分有效性的现象在很多研究中有报道[50-51]。 分别用棉花根系分泌物和中性去离子水(对照)添加到土壤中培养15 d 后,添加棉花根系分泌物培养的土壤速效磷和速效钾含量分别比对照增加19.2%和34.0%, 土壤酶活性比对照提高12.3%。 大田试验进一步验证了室内模拟试验的结果。 如套作棉花行间土壤速效磷、速效钾和微生物的总量均高于单作[15]。因此,增加的这部分养分资源不仅可以满足自身生长需求,而且还可以促进临体植物(棉花)生长。Rochester 等[52]分别研究豆科、非豆科与棉花间套种植对土壤氮素损失以及氮素肥力的影响,发现与非豆科/ 棉花间作相比,豆科/ 棉花间套种植可以降低土壤氮素损失45%,增加土壤氮素有效性,提高棉花产量,同时降低化学氮肥施用量高达127 kg·hm-2。在核桃/棉花农林复合系统中,树龄较小的核桃/棉花间作的微生物生物量碳为375 mg·kg-1,高于单作棉花的163 mg·kg-1和单作核桃树的88 mg·kg-1[53]。
3.3 化感作用
化感作用是植物通过释放化学物质抑制或促进临体植物生长、发育与繁殖的现象。 棉花根系分泌物能够促进土壤中磷和钾向有效态转化,速效磷和速效钾含量分别比对照提高19.2%和34.0%[15]。 通过向盆栽棉花添加小麦根区化感物质(酚酸类)[54],研究化感物质对棉苗生长的影响,结果表明:添加低浓度的化感物质对棉苗的生长影响较小,而高浓度的化感物质显著抑制了棉苗的生长,主要表现在棉苗干物质积累降低,根系内源保护酶活性显著下降, 丙二醛含量上升,电解质渗出率升高。 因此,小麦根系分泌物产生的化感作用影响了棉苗的生长,从而导致了麦套棉弱苗晚发[55]。但孟亚利等[51]研究发现,麦棉套作地下部不仅存在小麦对棉花营养竞争的不利一面,同时还存在小麦根系促进棉花根际土壤酶活性和土壤养分含量提高的有利一面,且此效应将持续在全生育期影响套作棉花。
利用作物间化感效应构建复合体系,可以有效地防治棉花病虫害的发生和危害,减少农药施用量[44]。 有研究表明大麦、小麦、向日葵等作物本身对杂草有化感作用, 可用来与其他作物间作,控制杂草生长,减少除草剂的用量[56]。棉花与高粱轮作,高粱根系的分泌液可以使棉花黄萎病病菌(大丽轮枝菌) 的种群数量减少或不能发育成菌丝,从而有效抑制棉花黄萎病的发生[57]。
4 问题和展望
相对于粮食作物的间(套)作(如小麦/ 玉米和蚕豆/ 玉米)[9],基于棉花等经济作物的间(套)作研究仍显相薄弱,且主要集中于单一作物的地上部生长和产量[22,25]、地下部根系分布[46,58-59]、或者相关农艺措施(如密度、品种、施肥和施用缩节胺等)的影响[23,60-62]。联系作物地上部生长、光热资源利用和地下部根系时空分布,从生态位分异和种间相互作用角度研究棉田间套复合种植强化生态系统服务功能相对较少。
由于豆科作物特有的生物固氮功能,禾本科/豆科间(套)作被广泛应用在低投入的农田生态系统[63-64]。Rochester 等[52]研究表明,在棉花种植体系中引进豆科作物可以提高土壤有效氮含量,降低土壤紧实度, 有利于下茬作物根系生长发育,进而增加棉花产量。 因此,在棉田间套复合种植体系中引进豆科作物被认为是1 种绿色可持续的种植模式[65]。但是,涉及豆科的棉花间(套)作体系种间氮素互补利用特征鲜见报道。 今后应该加强这方面的研究,从种间相互作用对豆科作物的固氮量、固氮比例以及作物种间氮素转移的调节作用等方面考虑。
大量研究表明,间套复合种植增加作物产量的同时,从土壤中带走的养分也多于单作[66],因此, 长期连续棉田间套复合种植是否会降低土壤肥力成为亟待研究的问题。 此外,多样性种植也常伴随着较高的地下部根系多样性, 包括根系生物量和化学组成[14]。 基于此,长期的间作套种是否会通过地下部根系和微生物多样性对土壤肥力产生正向反馈作用也是值得研究的科学问题。
农业的根本出路在于机械化,间(套)作种植模式也必须要实现机械化才有生命力[67]。 由于劳动力成本的上升和农村劳动力短缺现象的逐渐加剧,这一问题显得越来越重要。 要实现间(套)作生产的机械化,一方面要求农艺上有规范的高产高效的种间配置模式和行间距配置,另一方面具有相应的播种、 施肥和施药以及收获的机械。已有研究以“小麦- 玉米- 大豆”旱地新三熟带状复合种植模式为例,在西南丘陵小地块上基本实现了该带状复合种植模式下的耕地、播种、管理和收获等农业生产环节的机械化作业, 效率高,质量好[68]。为了进一步实现间(套)作的机械化,迫切需要在更多间(套)作模式和更广泛区域上进行试验和示范,使得间(套)作能够实现大面积的机械化。
