基于物候模型的气温对温带落叶阔叶林生态系统物候过程的影响
2020-10-09张银龙
龚 元, 张银龙
(1.南京林业大学生物与环境学院,江苏 南京 210037;2.南京林业大学江苏省南方现代林业协同创新中心,江苏 南京 210037;3.阿拉巴马大学生物科学系,塔斯卡卢萨 AL35487)
森林生态系统碳循环是全球陆地生态系统碳循环的重要组成部分,植物群落的光合作用使得森林生态系统可以在白天吸收一部分大气中的CO2[1-5],因此森林生态系统成为陆地生态系统中主要碳汇之一[6-10].由于受到植被生长环境、植物生理等因素的影响,森林生态系统会在不同时间尺度呈现不同的物候特征,且该生态系统物候过程会对森林生态系统碳循环产生直接影响[11-15].因此掌握不同森林生态系统的物候特征对更好地了解陆地生态系统碳循环过程、分析植被对全球气候变化的响应、合理森林布局都具有参考作用[16-17].
植物物候过程是陆地表面物候特征(land surface phenology, LSP)的重要组成部分,主要包括陆地植物群落生态、生理动态变化及周期性生物事件发生的时间节点[16].植物群落物候特征的研究内容主要包括生长季开始(start of growing season, SOS)、生长季节结束(end of growing season, EOS)和生长季长度(length of growing season,LOS/LGS)的估算等[16].早期陆地表面物候过程的研究主要是利用光学卫星观测数据(remote sensing, RS),即MODIS(https://modis.gsfc.nasa.gov/)植被指数产品(LAI/EVI/NDVI/GPP),统计生理模型拟合植被指数,模拟植物群落的生长曲线,并提取生态系统的关键物候时间节点[16].近年来,涡动协方差技术(eddy covariance, EC)的应用为直接观测陆地生态系统CO2排放动态提供了技术支持[16-21].涡动相关技术是用于观测一定生态尺度上较平坦的地球表面与大气之间的物质、热量以及动量交换的技术手段[22].为了更好地研究陆地生态系统的碳循环过程,将涡动相关技术应用于森林生态系统CO2通量的观测.通过涡动相关系统对一定生态尺度的下垫面与大气的碳交换进行长期且高时间分辨率的不间断观测,与遥感技术的长数据周期(≥3 d)[23-24]相比,EC具有较高的时间分辨率和数据精确度等.目前利用涡动相关系统的CO2通量观测数据对森林生态系统的物候特征进行量化成为植物物候学的重要研究内容[15-16].
基于涡动相关系统CO2通量观测值,使用统计模型提取植物物候过程的研究也较多.Richardson et al[14]使用国际通量网多站点森林CO2通量(GEP/NEP),基于物候模型分析了温带森林生态系统CO2通量对全球气候变化的响应,结果表明全球增温会延长温带森林生态系统生长季的长度.Niu et al[15]使用国际通量网CO2通量数据,基于物候模型分析了北半球生态系统CO2通量对年均气温变化的响应,结果表明森林通量站点的物候特征均对年均气温变化的敏感性较高.Gonsamo et al[16]使用国际通量网CO2通量数据,基于一个非线性逻辑函数分析了北美温带森林生态系统的物候特征,并且基于遥感数据(NDVI/NDII)开发了一个新的物候指数(PI),在PI与通量数据的交叉验证中,遥感数据与涡动通量数据均较好地体现了下垫面森林景观的物候过程.可利用非对称高斯函数、D-L拟合和S-G滤波法等[25]提取下垫面物候特征参数.在以上列举的统计模型中,具有代表性的是由Gu et al[17]构建的植物群落光合作用物候模型(CPC).该模型主要通过单日GPP 30 min观测值的最大值提取其动态变化的关键时间点,划分局域尺度生态系统的物候阶段.其物候阶段主要包括准备阶段、恢复阶段、稳定阶段、衰落阶段和终止阶段,该模型还包括计算生长季长度(LGS)、生长季中点、生长季开始和结束点(OGS/EGS)等物候特征参数[17].
基于涡动相关系统研究森林生态系统物候过程,对进一步了解陆地生态系统碳循环机制和应对全球气候变化具有参考意义.本研究利用国际通量网所提供的FLUXNET2015数据集[26](https://fluxnet.fluxdata.org/doi/FLUXNET2015/US-MMS)和美国摩根门罗州国有森林(US-MMS)通量观测站点2000—2014年共计15年的CO2通量数据,结合微气象物候模型,分析纬度局域温带森林生态系统的物候特征及其与气温的关系,为相关研究提供参考.
1 材料与方法
1.1 研究区概况
研究区为US-MMS,通量观测塔的地理坐标为(39°19′N, 86°25′W).该区域所属温度带为温带,为温带大陆性气候,多年平均降水量1 032 mm,多年平均气温10.85 ℃(图1),年平均入射光量子通量密度343.6 μmol·m-2·s-1,最冷月均温出现在1月份(约-1 ℃),最热月均温出现在8月份(约22 ℃)[27-28].该国家森林总面积约95.3 km2,搭载通量观测(CO2/H2O)和微气象观测设备的通量塔距离地面46 m(图2),下垫面冠层高度约26 m,下垫面海拔275 m[27-28].通过涡动相关系统(EC)观测CO2/H2O通量、风向和风速等数据.通过微气象梯度观测系统观测降水(P)、光合有效辐射(PAR)、土壤温度(TS)、净辐射(Rn)等微气象数据.在通量塔观测源区内的木本植物,主要包括枫树(Acersaccharum)、山毛榉(Faguslongipetiolata)等落叶阔叶林,树龄约60~80 a,平均冠层高度25~27 m,且该区域落叶阔叶林有明显的叶期和无叶期[27-28].该国家森林站点在FLUXNET的注册ID为US-MMS,从1998年开始对US-MMS进行生态观测,积累了较长时间的CO2通量数据和微气象数据,还包括地土壤温湿度、叶面积指数(LAI)等其它辅助生态数据.
1.2 涡动相关通量观测系统的组成
该US-MMS国有森林整体CO2/H2O通量采用涡动相关系统进行观测和记录[27-28].该系统由3个三维超声温度、风速测量仪器(Campbell Scientific Instruments, USA)和3个闭路式CO2/H2O红外气体分析仪器(Li-6262, Licor, USA)组成,其安装高度距离地面46、34、2 m(图2),通量塔地理坐标为(39°19′N, 86°25′W).通量和微气象数据的采集和存储设备为CSI 21X(Campbell Scientific Instruments, USA)10 Hz高频数据采集仪器,并且以30 min时间间隔计算存储CO2和H2O通量,以及微气象数据平均值[27-28].
本研究采用的通量与气象数据来自于国际通量网(https://fluxnet.fluxdata.org/),通过US-MMS站点(http://sites.fluxdata.org/US-MMS/)观测的生态系统GPP数据、空气温度和降水数据见表1.其中,GPP为生态系统总初级生产力,依据国际通量网数据处理标准完成通量数据质量控制和数据插补[27-28].
1.3 植物群落光合作用的物候模型
为了分析US-MMS国有森林2000—2014年植被物候特征的动态变化,以及探讨气温变化对生态系统物候过程的潜在影响,构建基于生态系统GPP观测数据的植物群落光合作用的物候模型[17].该模型基于年际尺度的每天半小时最大GPP来量化植物群落光合能力的季节变化[17].通过物候模型计算US-MMS国家森林日GPP累积值,模拟植物生长曲线(CPC),讨论该研究区植物群落的物候特征.植物群落光合作用的物候模型表示如下:
(1)
式中,A(t)函数表示该研究区的GPP数据,t表示日序.该模型中的参数均为经验拟合参数,且在计算拟合参数时可以依据经验和实际情况对所计算的参数进行调整[4,17].
(2)
式中,k(t)为该研究区的通量增长率函数;将年际尺度(1~365 d)的GPP数据带入式(1)中计算,可得到所需拟合参数,再将计算出的所有拟合参数带入k(t)函数即可计算出GPP增长率[4,17].另外,Gu et al[17]基于式(1)和式(2)定义了两条植物生长和衰落的直线,分别为生长季恢复线(recovery line, RL)和生长季衰落线(senescence line, SL),分别表示如下:
ARL(t)=kPRRt+A(tPRD)-kPRRtPRD
(3)
式中,ARL(t)为生长季恢复线.
ASL(t)=kPSRt+A(tPSD)-kPSRtPSD
(4)
式中,ASL(t)为生长季衰落线[17].
kPRR为GPP增长率最大值,即GPP最大恢复速率;tPRD为GPP最大恢复速率;kPSR为GPP增长率最小值,即GPP最大衰落速率;tPSD为GPP最大衰落速率,可由下式计算[17].
tPRD≈t01+b1lnc1
(5)
tPSD≈t02+b2lnc2
(6)
kPRR=k(tPRD)
(7)
kPSR=k(tPSD)
(8)
Gu et al[17]定义的植物群落最大光合作用能力(Ap),可以由下式计算:
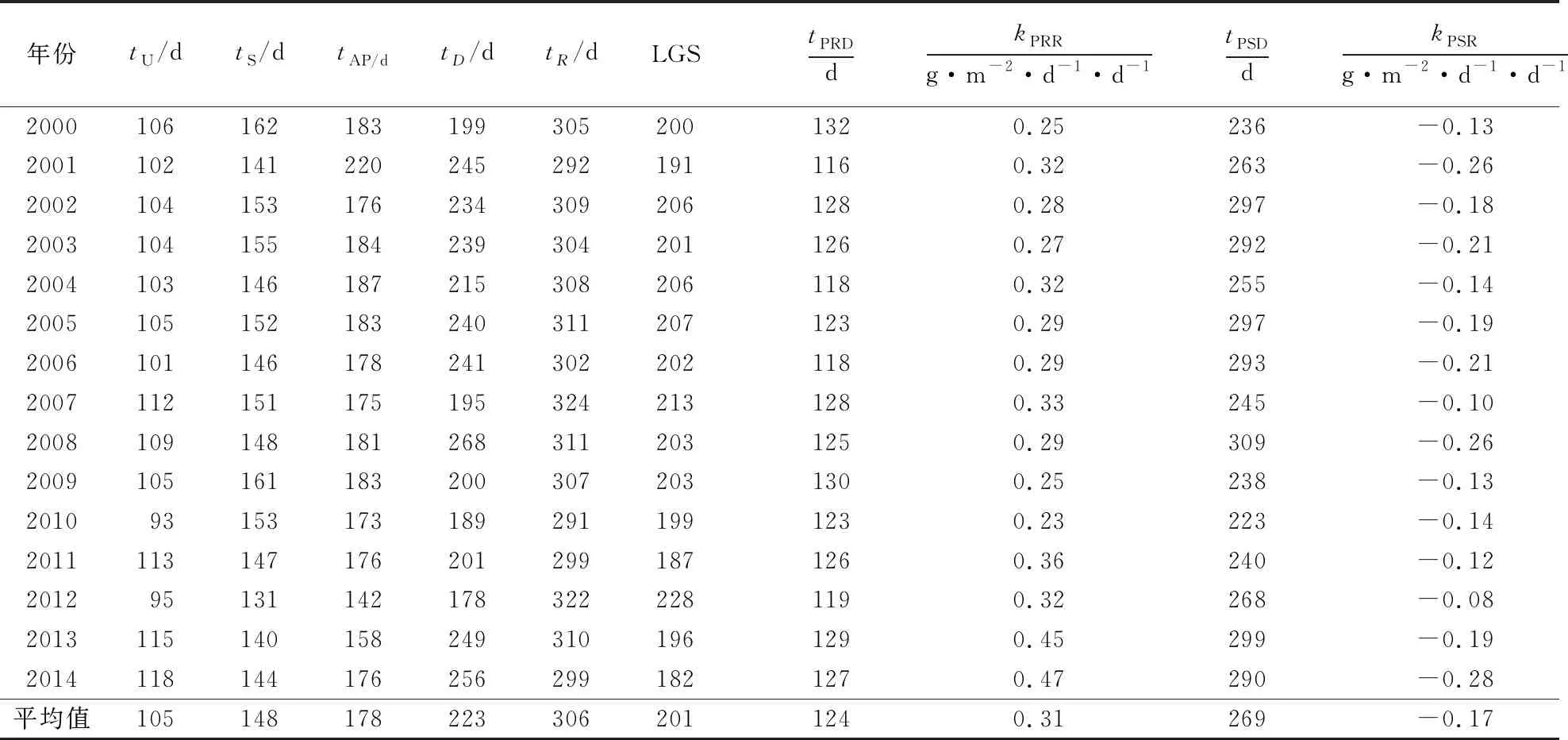
Ap=max{|A(t),tstart (9) 式中,tstart=1,tend=365.在获得RL和SL后,Gu et al[17]基于RL与x轴的交点定义生长季开始(upturn day,tU).当RL的y值达到Ap的t值时,定义为生长季稳定日(stabilization day,tS);当SL的y值达到Ap的t值时,定义为生长季下行日(downturn day,tD).SL与x轴的交点定义为生长季结束(recession day,tR).并且依据tU、tS、tD、tR的位置定义了准备阶段、恢复阶段、稳定阶段、衰落阶段和结束阶段等几个关键物候节点[17]. 为了分析该研究区温带国有森林生态系统(US-MMS)的物候特征,以及讨论气温变化对生态系统物候过程的潜在影响,利用US-MMS站点2000—2014年总计15年的GPP数据(表1),参考Gu et al[17]构建的植物群落光合作用物候模型,模拟该生态系统的植被生长曲线,提取物候时间点(表2).以2000年为例,利用物候模型计算该生态系统的植被生长季曲线A(t)、不同物候阶段和GPP增长率k(t),结果如图3所示. 表2 2000—2014年US-MMS生态系统的物候特征 表1 GPP与微气象数据描述 从图3可看出,黄色圆圈为GPP,红色曲线为植物生长曲线A(t),左侧黑色虚线为生长季恢复线RL,右侧黑色虚线为生长季衰落线SL,从左至右4条蓝色直线所代表的t值分别为tU、tS、tD、tR,蓝色虚线代表光合作用最强点tAP.据此判断US-MMS生态系统在2000年生长季的准备阶段为1~106 d,恢复阶段为106~162 d,光合作用最强点为183 d,稳定阶段为162~199 d,衰落阶段为199~305 d,结束阶段为305~365 d[17].全年整个生长季为106~305 d,生长季长度为200 d,光合作用能力峰值[17]为14.49 g·m-2·d-1.在图3右侧,红色曲线为GPP增长率k(t),左侧蓝色三角为GPP春季最大恢复点,右侧蓝色三角为GPP最大衰落点.据此可知US-MMS生态系统在2000年的tPRD、kPRR、tPSD、kPSR分别为132 d、0.25 g·m-2·d-1·d-1、236 d、-0.13 g·m-2·d-1·d-1.其中0.25 g·m-2·d-1·d-1代表RL的斜率,-0.13 g·m-2·d-1·d-1代表SL的斜率[17]. 从表2可知,US-MMS生态系统从2000—2014年生长季开始,约在第105 天,光合作用最强的日序为178 d,生态系统生长季准备阶段为1~105 d,生长季恢复期为105~148 d;光合作用稳定阶段约148~223 d,约从第223天生长季开始光合作用能力逐渐下降,直到第306天生态系统生长季结束,生长季衰落期为223~306 d,生长季长度约201 d.US-MMS生态系统单年生长季长度最大值为228 d,出现在2012年;单年生长季长度最小值为182 d,出现在2014年;其余年份生长季长度均为190~210 d.从表2可知,US-MMS生态系统春季光合作用最大恢复率的多年平均值为0.31 g·m-2·d-1·d-1,日序为124 d.秋季光合作用最大衰落率的多年平均值为-0.17 g·m-2·d-1·d-1,日序为269 d.其原因可能是全年124 d后,生态系统进入春季,气温开始回升,植物生长速率达到最大,吸收了更多的CO2,导致GPP增加;在全年269 d后生态系统由秋季进入冬季,气温的回落和植物的凋零导致光合作用减弱,GPP呈下降趋势[17].该研究的US-MMS生态系统GPP动态特征与Gu et al[17]研究的温带森林生态系统GPP增长率特征较一致. 2.2.1 生长季开始和结束对生态系统物候过程的影响 为分析US-MMS生态系统生长季开始和结束与生态系统生长季长度的关系,即春季和秋季对生态系统物候过程的影响,采用tU和tR与生态系统生长季长度进行线性回归分析,探讨生长季开始和结束的时间对生态系统生长季长度的影响,结果如图4所示. 从图4可看出,US-MMS生态系统在2000—2014年,生长季长度与生长季开始日、结束日的关系较明显,具体表现为生长季开始日与生长季长度呈负相关关系,即随着生长季开始日的提前或者推迟,生长季长度出现增加或者减少.生长季结束日与生长季长度呈正相关关系,即随着生长季结束日的提前或者推迟,生长季长度出现减少或者增加.其中生长季结束日对生长季长度的影响大于生长季开始日对生态系统生长季长度的影响.晚秋是该局域温带落叶阔叶林生态系统生长季的主要控制因子,即随着秋季的延长该生态系统生长季延长. 为进一步分析和讨论生态系统生长季开始日和结束日对该US-MMS生态系统物候过程的潜在影响,采用生态系统生长季开始日和结束日与生态系统生长季的统计学中点[17]进行线性回归分析,结果如图5所示.从图5可看出,US-MMS生态系统在2000—2014年,生态系统生长季中点与生长季开始日、生长季结束日的关系呈显著相关(P<0.01),且都呈正相关关系.生态系统生长季开始日和结束日对生态系统生长季中点的影响具体表现为随着生长季开始日和结束日的推迟或者提前,生长季中点也出现相应的推迟和提前,即生长季开始和结束日均会影响US-MMS生态系统夏季中期所出现的时间点.据此推测US-MMS生态系统生长季开始日的提前和结束日的推迟均会导致生态系统生长季长度延长,这与生态系统生长季开始日、结束日和生态系统生长季长度关系的分析结果较一致. 2.2.2 生长季恢复期和衰落期对生态系统物候过程的影响 为进一步分析春季和秋季长度对植物生长季的影响,采用Gu et al[17]定义的生长季恢复期和生长季衰落期与生长季长度进行对比分析,讨论春季和秋季长度对生长季长度的影响,结果如图6所示.从图6可看出,2000—2014年US-MMS生态系统生长季恢复期长度为25~60 d,平均值为43 d;生长季衰落期长度为43~144 d,平均值为83 d;其中生长季恢复期长度最大值出现在2010年,该年的生长季长度为199 d;生长季衰落期长度最大值出现在2012年,该年的生长季长度为228 d,2012年也是所有年份中生长季长度最长的年份.为进一步分析生长季恢复期长度、衰落期长度与生长季长度的关系,统计出恢复期长度和衰落期长度占整个生长季的比重,结果如图7所示. 从图7可看出,2000—2014年US-MMS生态系统生长季恢复期占生长季长度的百分比总体上低于生长季衰落期占生长季长度的百分比.其中生长季恢复期占生长季长度百分比为12%~27%,生长季衰落期占生长季长度百分比为21%~63%.因此2000—2014年US-MMS生态系统的物候过程主要受到秋季长度和起始时间的影响.为分析春季和秋季长度对生长季长度的影响程度,使用生长季恢复期长度和衰落期长度与生长季长度进行线性回归分析,结果如图8所示.依据线性回归分析结果,2000—2014年US-MMS生态系统物候过程主要受到生长季衰落期和生长季结束日的影响,生长季开始日和生长季恢复期对生态系统生长季长度的影响相对较小.随着US-MMS生态系统生长季衰落期的延长,生态系统生长季长度有延长的趋势,春季长度对生长季长度的影响较小. 气温是影响植物生长的重要环境控制因子,为讨论全球温室效应背景下2000—2014年气温变化对US-MMS生态系统生长季长度的影响,将年均气温与US-MMS生态系统生长季长度进行线性回归分析,结果如图9所示. 从图9可看出,US-MMS生态系统生长季长度与气温呈正相关关系,即随着年均气温的上升,生态系统生长季长度有增加的趋势.其原因可能是气温升高为植物生长提供了较好的光照和热量条件,促进了植物光合作用,增加了生态系统生长季长度[17].为进一步分析气温对生态系统物候过程的影响机制,对生态系统生长季的开始和结束与年均气温进行线性回归分析,讨论气温变化对春季和秋季的影响,结果如图10所示.从图10可看出,年均气温对US-MMS生态系统物候过程的影响,主要表现为影响生态系统生长季开始的日序,而对生态系统生长季结束的影响较小.年均空气温度对生态系统生长季开始的影响,主要表现为随着气温的上升生态系统生长季的开始日提前. 本研究结果表明春季增温会导致生态系统生长季提前,这与Richardson et al[14]和Zhu et al[33]的研究结果一致.Gu et al[17]认为在温带森林生态系统中,晚秋是生态系统生长季长度的重要影响因子.Noormets et al[29]研究结果表明美国田纳西混交林森林系统生长季长度约为205 d.Churkina et al[30]基于遥感数据研究的多站点温带森林生态系统的生长季长度约200 d.Wu et al[31]采用数据周期为8 d的遥感GPP数据基于非线性函数研究北美多站点森林生态系统物候特征,结果表明生态系统开始吸收CO2的时间点随着纬度的升高而推迟,生态系统结束光合作用的时间点随着纬度的上升而提前,且其研究的US-MMS站点的生长季长度约205 d.刘啸添等[32]基于遥感和GPP观测数据分析温带针阔混交林的物候特征,得到的结果也与本研究结果相似.Zhu et al[33]基于分段逻辑模型分析了1982—2006年北美植被物候变化的时空格局,结果表明植物休眠期的开始日期是导致生长季延长的主要原因,返青日对生态系统生长季长度的影响较小.Fisher et al[34]认为温带森林季节性温度变化会触发春季生长季的开始.Chmielewski et al[35]在对欧洲植被物候的研究中发现自1989年以来,随着平均气温上升下垫面植物生长季开始的日期有提前的趋势. 以上研究结果均与本研究结果一致,即气温变化对森林生态系统物候过程的控制方式,主要表现为增温会导致生长季开始日提前,气温增长对森林生态系统晚秋现象的影响较小.其原因可能是增温导致该生态系统生长季开始日提前,使得该生态系统中的植物群落对氮元素需求量增加,致使生态系统在夏季后期的植物光合作用容易受到氮元素总量的限制,致使夏末秋初的生态系统GPP开始下降,生态系统过早进入秋季,秋季成为该生态系统生长季长度的主要贡献因子[6]. 本研究结果表明:(1)US-MMS温带森林生态系统在2000—2014年生态系统生长季开始日约105 d,第223天光合作用能力开始下降,直到第306天生态系统生长季结束,生态系统生长季长度约201 d;(2)增温会导致US-MMS温带森林生态系统生长季长度的延长,生态系统春季GPP对增温敏感;(3)气温会影响US-MMS生态系统的物候过程,但气温的动态变化不是生态系统物候过程的唯一环境控制因子.气温影响生态系统物候的方式为通过控制生长季开始日来影响生态系统物候过程,即增温会导致生态系统生长季开始日的提前,但气温对生态系统生长季结束日的影响较小.2 结果与分析
2.1 生态系统物候特征

2.2 生态系统物候过程
2.3 气温对生态系统物候过程的影响
3 讨论与小结
3.1 讨论
3.2 小结
