熟地黄对ADHD 模型大鼠前额叶皮质线粒体的保护作用
2020-09-15袁海霞倪新强韩新民陈天翼喻闽凤宋宇尘
袁海霞 倪新强 韩新民 郑 敏 陈天翼 喻闽凤 宋宇尘
(1.南京中医药大学中医儿科学研究所, 江苏南京210023; 2.深圳市中医院, 广东深圳518038)
注意缺陷多动障碍 (attention deficit hyperactivity disorder,ADHD) 是儿童时期常见的神经发育障碍性疾病,我国的发病率约为6.29%[1],其核心症状为注意缺陷、多动和冲动,常与焦虑、品行障碍、对立违抗障碍、学习困难等共存,并与自杀和犯罪活动关系紧密,严重影响患儿健康成长[2⁃3]。ADHD 是一种病因复杂的异质性疾病,其发病机制尚未被完全揭示。越来越多的研究表明线粒体功能紊乱与ADHD 发病有密切联系。研究发现与能量代谢相关的线粒体DNA 10398A/G的多态性引起的线粒体功能障碍可能是ADHD 的重要遗传病因[4]。有病例报道,ADHD 儿童的线粒体DNA 存在点突变,另有一名在青春期患ADHD的中年女性存在线粒体DNA 序列的重新排列[5]。有学者利用线粒体DNA 缺失细胞(ρ0 细胞) 分别与ADHD 患儿和正常儿童的血小板进行细胞融合构建胞质杂交细胞,结果发现,与正常儿童相比,ADHD 胞质杂交细胞呈现出明显的线粒体功能缺陷,表现为线粒体膜电位低下、氧化应激升高等[6]。
另外,有影像学研究表明,ADHD 患儿大脑皮质(尤其是前额叶皮质) 存在明显的成熟延迟,表现为皮质达到50%峰厚度的平均年龄比正常儿童晚3 岁[7]。前额叶皮质是调控自发性活动、注意力、学习记忆和认知能力的重要脑区,神经元发育障碍在其成熟延迟的机制中扮演不可或缺的角色,而线粒体功能障碍至关重要地参与了神经元发育障碍的病理过程[8]。综上所述,深入探索线粒体功能紊乱在ADHD 病因病机中扮演的角色以及寻求靶向保护线粒体功能的治疗手段具有重要意义。鉴此,本实验以幼龄自发性高血压大鼠(spontaneously hypertensive rats,SHR) 为儿童ADHD 模型,针对熟地黄调控其前额叶皮质线粒体功能的作用及相应机制展开研究。
1 材料
1.1 动物 SPF 级SHR 大鼠30 只,Wistar⁃Kyoto(WKY) 大鼠10 只,均为雄性,3 周龄,购自北京维通利华实验动物技术有限公司,实验动物生产许可证号SCXK (京) 2016⁃0011。动物饲养于12 h光照/12 h 黑暗逆转交替(人工光照,9: 00 关灯,21: 00 开灯)、室温20~22 ℃、相对湿度45%~55%的动物房中,自由进食及饮水。
1.2 药物 熟地黄Rehmannia glutinosa(Gaetn.)Libosch.ex Fisch.et Mey.购于北京同仁堂药店(批号151001,产地河南),由南京中医药大学中药资源学教研室主任巢建国教授鉴定为正品。称取熟地黄适量,第1 次加入10 倍量蒸馏水,浸泡0.5 h,武火煎沸后改用文火煎45 min;第2~3 次分别加8 倍量水煎30 min;每次均用8 层纱布过滤,合并3 次药液,真空冷冻干燥,得熟地黄水提物。盐酸哌甲酯缓释片 (Methylphenidate,MPH,国药准字J20151103) 为阳性对照药,购自西安杨森制药有限公司。全部药物均用0.5% CMC⁃Na 配置成混悬液,超声溶解,4 ℃保存。
1.3 仪器 UC7 超薄切片机(德国Leica 公司);HT7700 透射式电子显微镜(日本Hitachi 公司);FACS Calibur 流式细胞仪(美国Becton⁃Dickinson公司);BS⁃420 全自动生化仪(深圳迈瑞生物医疗电子股份有限公司)。
1.4 试剂 2%多聚甲醛-2.5%戊二醛混合固定液(货号 JRS0278)、4% 戊二醛固定液 (货号JRS0293),4%多聚甲醛固定液(货号JRS0271)、丙酮(货号E646)、锇酸(货号r022953) 均购自南京金益柏生物科技有限公司;DMEM 培养基(货号10569010) 购自美国Gibco 公司;胎牛血清(货号FCS500) 购自上海依科赛生物制品有限公司;线粒体膜电位检测试剂盒(货号KGA604)、活性氧检测试剂盒(货号KGT010⁃1) 购自江苏凯基生物技术股份有限公司;超氧化物歧化酶测试盒(货号HY⁃60001)、过氧化氢酶试剂盒(货号HY⁃60015) 购自北京华英生物技术研究所;TUNEL 细胞凋亡检测试剂盒(货号C1098) 购自碧云天生物技术有限公司。
2 方法
2.1 分组和给药 大鼠适应性喂养5 d,采用随机区组设计,将SHR 大鼠分为3 组,每组10 只,分别为模型组、MPH 组 (2 mg/kg)、熟地黄组(2.4 g/kg),WKY 大鼠为正常组。熟地黄给药剂量为生药剂量,前期数据挖掘显示熟地黄10 g 为临床治疗儿童ADHD 使用频次最多的单日剂量,参照《中药药理研究方法学》[9]方法进行大鼠与人临床用药剂量之间的换算,并结合参考文献研究[10]进行优化,最终得出熟地黄给药剂量为2.4 g/kg。正常组和模型组均以0.5% CMC⁃Na 溶液灌胃,给药体积为2 mL/kg。每日称定大鼠体质量及摄食量后,于9: 30~10: 30、15: 30~16: 30分2 次灌胃,连续干预4 周。
2.2 透射电镜观察前额叶皮质线粒体超微结构 各组大鼠先做麻醉处理,以2%多聚甲醛-2.5%戊二醛混合液进行慢速灌流,迅速取出全脑,切取前额叶皮质(不超过1 mm×1 mm×1 mm),置于4%戊二醛溶液4 ℃固定2~4 h;0.1 mol/L 磷酸缓冲液(PBS) 漂洗,1%锇酸-0.1 mol/L PBS 室温下进行后固定2 h;后经漂洗,酒精⁃丙酮梯度脱水,浸透,包埋聚合、切片(60~80 nm)、铀铅双染色,在透射电子显微镜下观察,采集图像分析。
2.3 JC⁃1 染色法检测MMP 各组大鼠麻醉后分离前额叶皮质,胰酶消化1 h,DMEM 培养基终止消化,调整细胞浓度约为1×106/mL,按照试剂盒说明书配制孵育缓冲液和JC⁃1 工作液,取500 μL JC⁃1 工作悬浮细胞,细胞培养箱37 ℃孵育20 min;2 000 r/min 室温离心5 min,收集细胞,孵育缓冲液清洗2 次,重悬,流式细胞仪检测,统计绿色荧光百分比。
2.4 DCFH⁃DA 荧光探针检测细胞内ROS 水平 标本采集及调整细胞浓度同“2.3” 项下,按照试剂盒说明书用无血清培养液稀释DCFH⁃DA(1 ∶1 000),用稀释后的DCFH⁃DA 悬浮细胞,37 ℃细胞培养箱内孵育20 min,每5 min 颠倒混匀1 次,使探针和细胞充分接触;无血清细胞培养液洗涤细胞3 次;流式细胞仪检测细胞内ROS 水平。
2.5 比色法检测SOD、CAT 水平 各组大鼠麻醉后分离出前额叶皮质,取10 mg 组织分别存放于4 ℃预冷的缓冲液中,采用低温研磨仪制备组织匀浆,10 000g离心5 min,取上清液,按照试剂盒说明书进行操作,检测前额叶皮质中SOD、CAT水平。
2.6 TUNEL 染色检测神经元凋亡 各组大鼠麻醉后用4% 多聚甲醛固定液进行灌注,经固定、脱水、透明、包埋切片、脱蜡、水化后按照TUNEL细胞凋亡检测试剂盒说明书进行染色处理,中性树胶封片后在光学显微镜下观察细胞凋亡情况。
2.7 统计学分析 采用SPSS 19.0 进行统计学分析,计量数据以() 表示。多组间比较方差齐则采用单因素方差分析,方差不齐则采用秩和检验。P≤0.05 为差异有统计学意义。
3 结果
3.1 熟地黄对SHR 大鼠进食量的影响 如表1 所示,药物干预第2 周时,与模型组比较,熟地黄组的进食量下降(P<0.01);第4 周时,MPH 组进食量与模型组比有下降趋势(P<0.05),而熟地黄组与MPH 组比较,差异无统计学意义(P>0.05)。
表1 各组大鼠每周平均进食量(g,, n=10)Tab.1 Average weekly food intake of rats in each group (g,, n=10)

表1 各组大鼠每周平均进食量(g,, n=10)Tab.1 Average weekly food intake of rats in each group (g,, n=10)
注:与同时间模型组比较,*P<0.05,**P<0.01。
3.2 熟地黄对SHR 大鼠体质量的影响 如表2 所示,与同时间模型组相比,MPH 组和熟地黄组大鼠的体质量差异无统计学意义(P>0.05)。
表2 各组大鼠每周平均体质量(g,, n=10)Tab.2 Average weekly body weight of rats in each group (g,, n=10)
表2 各组大鼠每周平均体质量(g,, n=10)Tab.2 Average weekly body weight of rats in each group (g,, n=10)
3.3 熟地黄对SHR 大鼠前额叶皮质线粒体超微结构的影响 如图1 所示,相较于正常组,模型组前额叶皮质线粒体出现肿胀扩张,线粒体嵴断裂消失严重,有的线粒体出现固缩,内部结构不清晰,其神经元核膜也呈现出边界模糊,有断裂。与模型组比较,MPH 组线粒体仍有肿胀扩张,线粒体嵴消失较多,仍可见多处的神经元核膜断裂;熟地黄组线粒体内部结构相对清晰,可见线粒体嵴,未见明显肿胀的线粒体,其神经元核膜虽有断裂,但内外膜有相对清晰的边界。
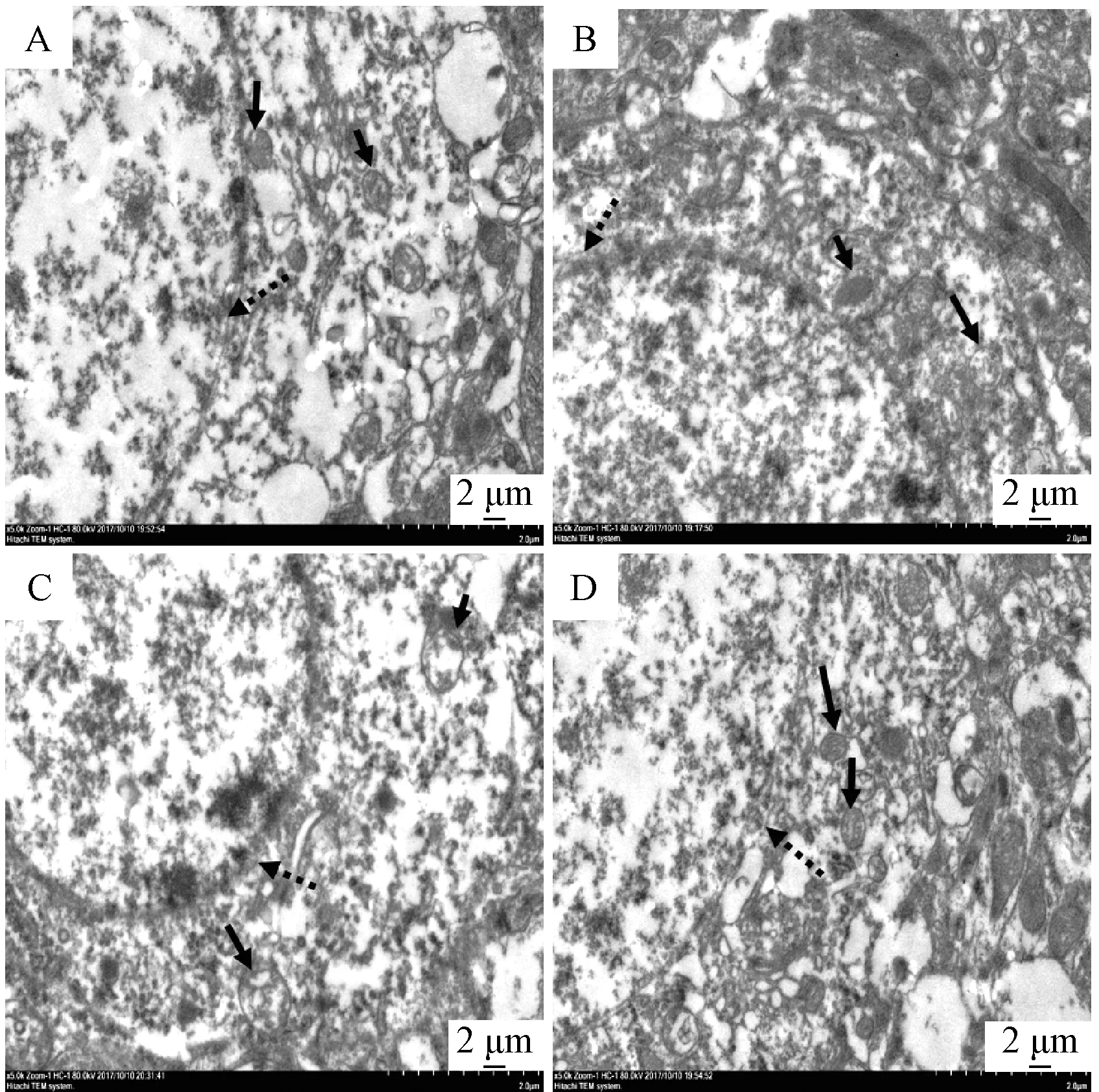
图1 各组大鼠前额叶皮质线粒体超微结构Fig.1 Ultrastructure of prefrontal cortex mito⁃chondria of rats in each group
3.4 熟地黄对SHR 大鼠前额叶皮质MMP 的影响 MMP较高时,JC⁃1 形成聚合物产生红色荧光;MMP 较低时,JC⁃1 为单体产生绿色荧光,因此绿色荧光水平与MMP 呈负相关。如图2 所示,与正常组比较,模型组大鼠前额叶皮质MMP 降低(P<0.01);与模型组比较,MPH 组MMP 下降(P<0.01);熟地黄组MMP 则升高(P<0.01)。
3.5 熟地黄对SHR 大鼠前额叶皮质细胞内ROS水平的影响 如图3 所示,模型组大鼠前额叶皮质细胞内ROS 水平高于正常组(P<0.01);与模型组比较,MPH 组细胞内ROS 水平升高(P<0.01),而熟地黄组细胞内ROS 水平则降低(P<0.01)。
3.6 熟地黄对SHR 大鼠前额叶皮质SOD、CAT 水平的影响 如图4 所示,模型组大鼠前额叶皮质SOD、CAT 水平低于正常组(P<0.01);与模型组比较,MPH 组SOD 水平下降(P<0.05),熟地黄组SOD、CAT 水平升高(P<0.05,P<0.01)。

图2 各组大鼠前额叶皮质MMP (n=3)Fig.2 Prefrontal cortex MMP of rats in each group(n=3)

图3 各组大鼠前额叶皮质细胞内ROS 水平(n=3)Fig.3 Prefrontal cortex ROS level of rats in each group(n=3)
3.7 熟地黄对SHR 大鼠前额叶皮质神经元凋亡的影响 如图5 所示,模型组大鼠前额叶皮质神经元凋亡水平高于正常组(P<0.01);与模型组比较,MPH 组神经元凋亡增加(P<0.05),而熟地黄组神经元凋亡降低(P<0.01)。
4 讨论
经典的益肾填髓中药熟地黄是目前公开发表治疗ADHD 的有效复方中使用频次最多的药物[11];另外,绝大多数以补肾填精为主要治法的复方均将熟地黄列为君药[12⁃14],可见熟地黄是治疗ADHD的核心中药。本课题组已基于“肾脑相关” 以及“皮质成熟延迟” 理论提出了“熟地黄可能通过改善神经元发育障碍控制ADHD 核心症状” 的假设[15]。前期实验研究表明,熟地黄可以显著减少儿童ADHD 动物模型幼龄SHR 大鼠的多动、冲动行为,并能提高其空间学习记忆能力[16]。本研究是对假设和前期实验的延展性探索,试图从线粒体功能的角度阐释熟地黄改善ADHD 症状的潜在机理。
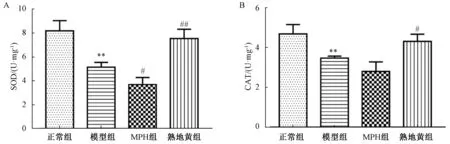
图4 各组大鼠前额叶皮质细胞内SOD、CAT 水平(n=3)Fig.4 SOD and CAT levels in prefrontal cortex of rats in each group (n=3)

图5 各组大鼠前额叶皮质神经元凋亡(n=3)Fig.5 Neuronal apoptosis in prefrontal cortex of rats in each group (n=3)
有研究表明,以MPH 为代表的精神兴奋剂虽能有效缓解ADHD 的核心症状,但长期服用此类药物可产生食欲减退、生长抑制等不良反应,会造成较低的体质量指数[17⁃18]。本研究发现,前3 周的MPH 干预对SHR 大鼠的进食量无明显影响;在第4 周时MPH 组进食量出现显著减少。第2 周时,熟地黄会降低SHR 大鼠的进食量,第3~4 周则对其进食量无明显改变。由此可见,熟地黄短期干预会影响SHR 大鼠进食量,这可能是由于中药特殊气味、口感或黏度影响其食欲造成,当长期服用适应后,进食量则不受影响。相比熟地黄,MPH 干预对SHR 大鼠食欲的不良影响在第4 周时才逐渐显现。二者对SHR 大鼠体质量未见明显影响。ADHD 是慢性精神发育障碍性疾病,其漫长的治疗期使得长期药物干预引发的不良反应更值得关注,从这一角度出发,熟地黄似乎比MPH 表现出一定优势。但因本实验未能评估药物对其体质量指数的影响;另外,未对停药后各组大鼠的进食量和体质量进行持续观测,因此,尚不足以得出相关确切的结论。
线粒体是细胞能量产生的主要场所,完整的结构是其发挥功能的物质基础。研究表明,与WKY大鼠相比,SHR 大鼠前额叶皮质存在线粒体功能缺陷,其ATP 合成率降低了30%,这为ADHD 可能是一种“能量缺陷” 疾病提供了依据[19]。有学者采用透射电镜比较了WKY 和SHR 大鼠前额叶皮质线粒体的超微结构,发现SHR 大鼠线粒体嵴和膜融合模糊不清,提示其线粒体结构存在异常[20]。与此结果相近,本研究发现SHR 大鼠前额叶皮质线粒体数量相对减少,有的线粒体肿胀扩张,嵴消失严重,呈现空泡化,有的线粒体出现固缩以致内部结构不清晰;除此,其神经元核膜亦有多处断裂,边界模糊不清。由此可以推测,SHR 大鼠前额叶皮质存在的线粒体数量减少和结构异常导致的功能缺陷极可能是其ATP 合成率降低从而引起能量缺乏的重要原因,亦或引发了神经元核膜结构的异常。这一结果从动物模型层面提示线粒体功能缺陷介导的神经元发育障碍可能参与了ADHD 的发生发展。经过4 周的干预,MPH 未能缓解SHR 大鼠前额叶皮质神经元核膜和线粒体的上述异常;熟地黄则减轻了其线粒体的肿胀,减少了嵴的消失,使之内部结构相对清晰。这从超微结构层次提示熟地黄对线粒体及神经元具有一定保护作用。
MMP 是反应线粒体功能的重要指标。MMP 降低会引起线粒体细胞色素C 的释放,进而触发凋亡级联反应,因此是细胞凋亡的早期特征。本研究发现,与WKY 大鼠比较,SHR 大鼠前额叶皮质MMP 显著降低,结合透射电镜结果,提示SHR 大鼠线粒体存在由结构异常导致的功能损伤。4 周的干预后,MPH 降低而熟地黄则提高了MMP 水平,提示熟地黄具有保护线粒体结构和功能的双重作用。线粒体能量代谢过程中产生的ROS 参与重要的生理活动,比如细胞生长和分化。然而,过度的生成以及失去抗氧化酶系统如SOD 和CAT 平衡的ROS 增多会直接导致线粒体DNA 损伤[21]。研究表明,SHR 大鼠前额叶皮质呈现出另一种抗氧化酶—谷胱甘肽过氧化物酶的活性降低,以及缺乏有效抗氧化平衡的ROS 的增加[22]。本研究结果与之相似,发现与WKY 大鼠相比,SHR 大鼠前额叶皮质细胞内ROS 明显增加,而SOD 和CAT 的活性则显著降低,说明SHR 大鼠脑部存在与其行为特征相关的氧化还原稳态失衡。有学者研究发现,MPH治疗可能诱发SHR 多个脑区的脂质过氧化、蛋白质羰基化增加,以及抗氧化酶的活性降低[23]。本研究中,MPH 的干预不仅增加SHR 大鼠前额叶皮质ROS 的水平,同时降低SOD 活性,说明MPH暴露会加重SHR 大鼠线粒体的氧化损伤,这与前人的研究结论相似[24]。与MPH 相反,熟地黄的治疗则可通过减少SHR 大鼠前额叶皮质细胞内ROS水平,提高SOD 和CAT 的活性维持氧化还原稳态,缓解线粒体氧化损伤,这可能是其发挥线粒体保护作用,进而改善SHR 大鼠ADHD 样行为特征的机制之一。
如前所述,线粒体不仅是能量代谢的场所,并且参与了内源性细胞凋亡。而能量代谢障碍与神经元凋亡已被证实与ADHD 的发生发展有密切联系[25⁃26],本研究已初步表明熟地黄可抑制SHR 大鼠前额叶皮质神经元凋亡。那么,熟地黄是否能够调控能量代谢以及抑制神经元凋亡的相关机制仍值得纵深研究。
