基于SSR分子标记的78份核桃种质资源遗传多样性分析
2020-07-23陈利娜李好先唐丽颖曹尚银
敬 丹,骆 翔,陈利娜,李好先,唐丽颖,曹尚银
(中国农业科学院 郑州果树研究所,河南 郑州 450009)
核桃(JuglansregiaL.)属于胡桃科、核桃属的木本树种,是世界著名四大干果之一,也是世界上重要的油料树种,与油桐、油茶、乌桕并称我国四大木本油料树种[1]。核桃具有较高的经济价值,其中叶、青皮、核仁、木材以及核桃林本身的经济价值都很高[2]。核桃还具有重要的营养药用价值,核桃仁是上好的滋补品,含有多种营养成分,对人体健康及饮食健康起到了重要作用。我国是核桃属植物的起源和分布中心之一,栽培历史悠久,种质资源丰富,分布广泛[3]。加强对种质资源的收集和保护,既是对优良基因的一种保护,又是种质资源创新的前提。一般来说农家品种对自生境有较强的适应性,含有更多优良基因。然而核桃优良的农家品种资源分布较散,往往分布在山地、丘陵地区,收集存在着一定的障碍和困难,国内也尚未有专门单位对其进行收集。此外,将收集来的农家品种进行鉴定和分类保存不仅需要专门的种质资源圃,也需要耗费大量的人力、物力。再者由于传统的形态学鉴定方法存在一定的短板之处,随着核桃品种的多样化,一些亲缘关系较为接近的品种仅靠形态学鉴定方法难以鉴别品种间的差异,导致研究者对农家品种的重视程度不高[4]。因此,加强核桃优良农家品种的收集,建立一种有效的苗木品种资源鉴定方法,对于核桃种质资源的创新及核桃产业的发展尤为重要和迫切。
分子标记技术是在形态标记、细胞标记和生化标记后出现的一种新兴的技术手段,它以DNA多态性为基础,直接反映DNA水平上的遗传变异[4]。目前常用的分子标记有限制性片段长度多态性(RFLP)、随机扩增多态性DNA(RAPD)、扩增片段多态性(AFLP)、单核苷酸多态性(SNP)、简单重复序列(SSR)等[1]。在果树的育种工作中,分子标记技术可用于研究果树种质资源及亲缘关系的远近、分析其遗传多样性,这对于果树种质资源的评价和保存具有重要意义,同时还可利用分子标记技术辅助育种,加快育种效率。目前,分子标记技术已被广泛应用于植物遗传研究和育种实践中,如桃、苹果、梨、核桃、茶树等[4-5]。Freeman等利用SSR技术对不同地域茶树品种的亲缘关系及居群的分化程度进行了分析[6]。张鹏等对42份云南玉米自交系进行了遗传多样性分析,为云南地区玉米育种提供了参考依据[7]。在核桃方面的应用主要集中在核桃种群遗传结构分析、遗传标记开发等方面[8-9]。目前核桃中已有数百对特异性SSR引物,它们已被证明在核桃的多种遗传分析中具有实用价值[10]。但是大部分是针对核桃的优良品种之间的遗传多样性及亲缘关系的研究,而对于核桃农家品种资源的相关研究尚未见报道。本研究旨在收集分布在全国各地的核桃农家品种资源,利用SSR标记对所收集到的核桃农家品种及部分栽培种进行分子标记遗传多样性分析,以期为核桃农家品种资源的保存和利用以及核桃种质资源的创新奠定一定的工作基础。
1 材料与方法
1.1 试验材料
试验材料为收集来的78份核桃资源的幼嫩叶片,采后用锡箔纸包裹,在液氮中速冻,然后带回置于-80 ℃超低温冰箱保存,所有材料均采自中国农业科学院郑州果树研究所品种资源圃(表1)。
1.2 试验方法
1.2.1 DNA 提取 采用CTAB法提取78份核桃叶片的基因组DNA。利用1%的琼脂糖凝胶以及Bio-Photometer核酸检测仪(Eppendorf)检测DNA的质量、浓度与纯度,并将DNA样品的浓度稀释至50 ng/μL,置于-20 ℃冰箱保存备用。
1.2.2 SSR分析 以78份核桃的基因组DNA为模板,根据已发表的NCBI公共数据库中核桃的SSR引物,在各材料中进行扩增,所有引物均由尚亚生物技术有限公司合成。所用反应体系为20 μL的PCR反应体系,其中10×PCR buffer 2 μL,2.5 mmol/L的MgCl21.6 μL,2.5 mmol/L 的4×dNTP 1.2 μL,4 μmol/L的上、下游引物各0.8 μL,5 U/μL的Taq DNA聚合酶0.1 μL,30 ng/μL的DNA 2 μL,用ddH2O补至体积为20 μL。PCR反应程序为:94 ℃ 4 min;94 ℃ 40 s,45~60 ℃ 40 s,72 ℃ 1 min,35个循环;72 ℃ 10 min;4 ℃保存。PCR 扩增产物用6%非变性聚丙烯酰胺凝胶在DYY-Ⅱ型垂直板电泳仪及 DYC-30 型电泳槽中检测。吸取PCR产物2 μL进行加样,上样完毕后在20 mA电流条件下电泳40 min。电泳结束后进行银染。
1.2.3 谱带的记录及数据统计与分析 根据各分子标记在相同电泳迁移率 (相同分子量片段) 的有无对银染后的凝胶图像进行条带统计,在相同位置上,有DNA扩增条带记为“1”,无带记为“0”。利用PowerMarker V3.25软件计算等位基因数(NA)、多态信息含量(PIC)及遗传距离(GD)等信息。利用NTsys 2.1软件采用SAHN功能进行UPGMA(类平均法)聚类分析。利用Structure 2.2软件进行78份核桃资源群体结构分析,其中“Length of Burnin Period”参数设为10000,“Number of MCMC Reps after Burnin”设为50000,K设置为1~10,每个K值重复20次。最佳K值根据L(K)和ΔK随K值的变化确定[11],其中L(K)及ΔK的计算参照曾可为等[12]的方法进行。
2 结果与分析
2.1 78份核桃农家品种资源SSR标记多态性分析
在78份核桃农家品种资源中,一共从25对SSR引物中筛选出8对具有一定多态性的SSR引物,如表2所示。

表2 SSR标记引物信息
对这8对引物进行分析发现,等位基因(NA)数量变幅为1~16,共扩增出了55个多态性位点,平均每对引物扩增出的等位基因数为6.875。其中引物M02的扩增产物最多,扩增出16条带;最少的为引物W08,仅有1条带。此外,主等位基因频率(MAF)、基因多样性(Gene Diversity)、多态信息含量(PIC)之间也存在一定的差异。每个SSR位点的PIC在0.075~0.228,平均为0.151。MAF范围为0.823~0.959,平均为0.893。基因多样性范围为0.078~0.271,平均为0.173(表3)。
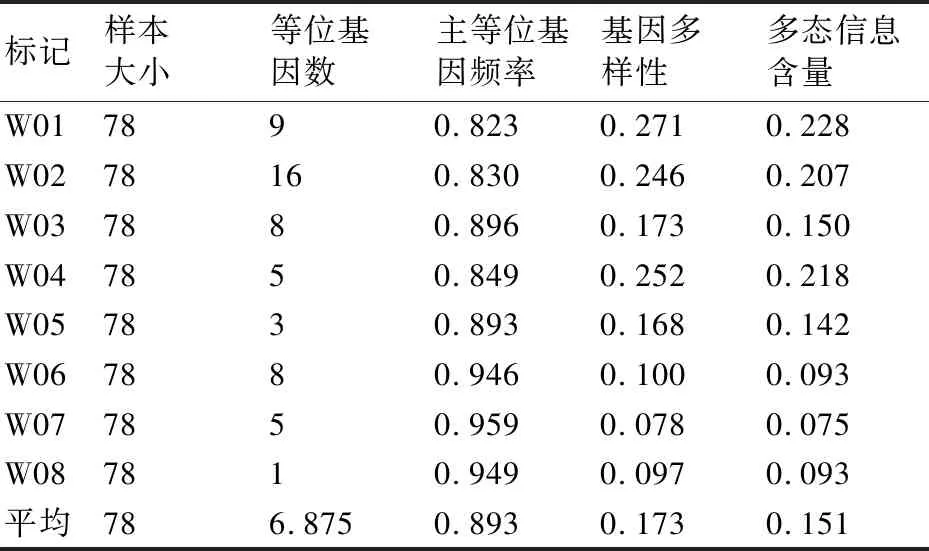
表3 8对SSR引物扩增结果及多态性信息
2.2 78份核桃品种间遗传多样性分析
基于这8对引物,采用UPGMA方法对78份资源进行聚类分析。78份核桃品种资源之间的遗传关系较为接近,遗传相似系数在0.605和1.000之间发生变化,其中陇南L(H1)和高岭4号(H18)、宋南3东4(H17)和中短05(H3)、济西11号(H21)和寿长核桃1号(H37)、响水湖怀柔5号(H22)和汾阳绵核桃10号(H28),安家滩4号(H25)、抗晚霜核桃(H26)和王河核桃2号(H27)、宋南2东5(H5)和中短22(H8)之间的遗传相似系数较大,遗传距离较小,说明这些品种间的差异较小,遗传关系较为接近。由图1可知,当Neis遗传距离为0.704时,可将78份核桃资源分为5部分,分别为Ⅰ、Ⅱ、Ⅲ、Ⅳ和Ⅴ。其中Ⅰ和Ⅱ品种较多,Ⅰ包括65个品种,占供试材料的83.33%。Ⅱ包括9个品种,占供试材料的11.54%。Ⅲ和Ⅳ分别仅包含GL4(H72)、西藏核桃(H69)一个品种,分别来自北京和西藏。Ⅴ包含两个品种,即宋南5东6(H76)和宋南2东9(H74),两者均来源于河南巩义。此外,该群体内的平均遗传距离为0.23,遗传距离0.2~0.3所占比例最大,为26.86%;遗传距离0.5以内的占96.38%(图2)。
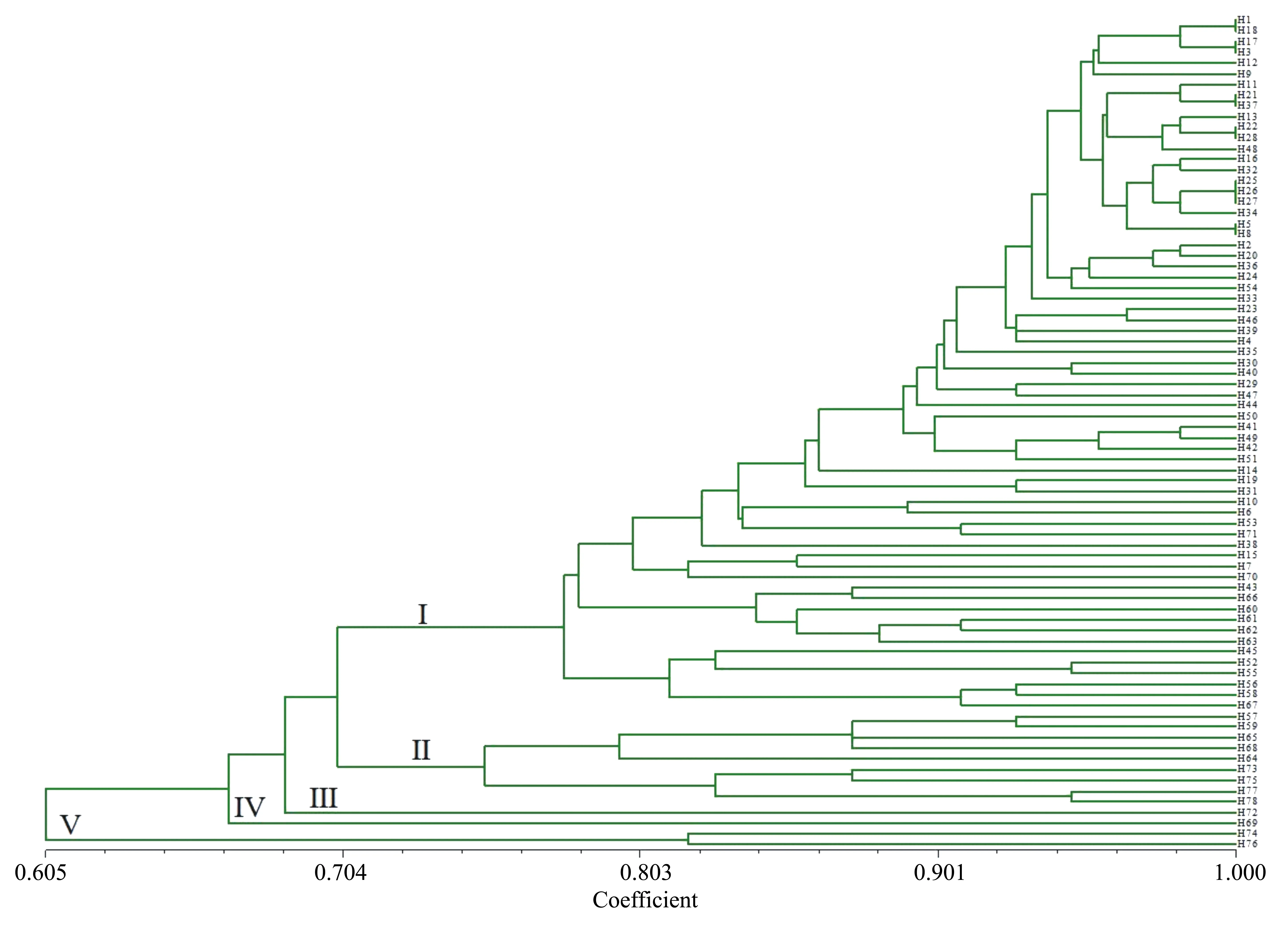
图1 78份核桃资源的UPGMA聚类结果
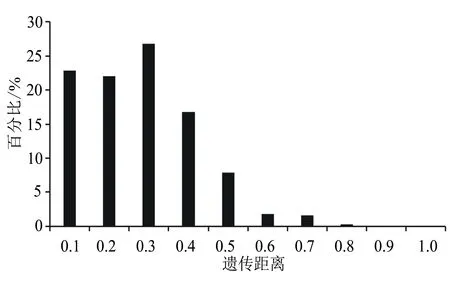
图2 78份核桃资源的遗传距离分布
2.3 群体结构分析
利用Structure 2.2软件对78份核桃资源进行群体结构分析,结果发现随着K值的增加,对数极大似然值L(K)值呈现递增的趋势,无明显的转折点(图3A);但ΔK值随K值增加呈先增加后下降的趋势。根据最大K值确定最佳亚群数[13],当K=2时,ΔK值出现最大峰值(图3B),由此说明该群体最适可划分为2个亚群(图4),分别为Q1(红色)和Q2(绿色)。其中Q1包含25份核桃资源,主要来自河南(10份),其余来源于北京、甘肃、新疆及河北等地;Q2包含53份核桃资源。78份资源未明显按照地理来源进行群体划分,该结果与UPGMA聚类结果相似,说明群体的划分与地理来源不完全相关。此外有31份种质既含有红色部分,又包含绿色部分,这些资源可能是两个祖先亚群杂交而来。
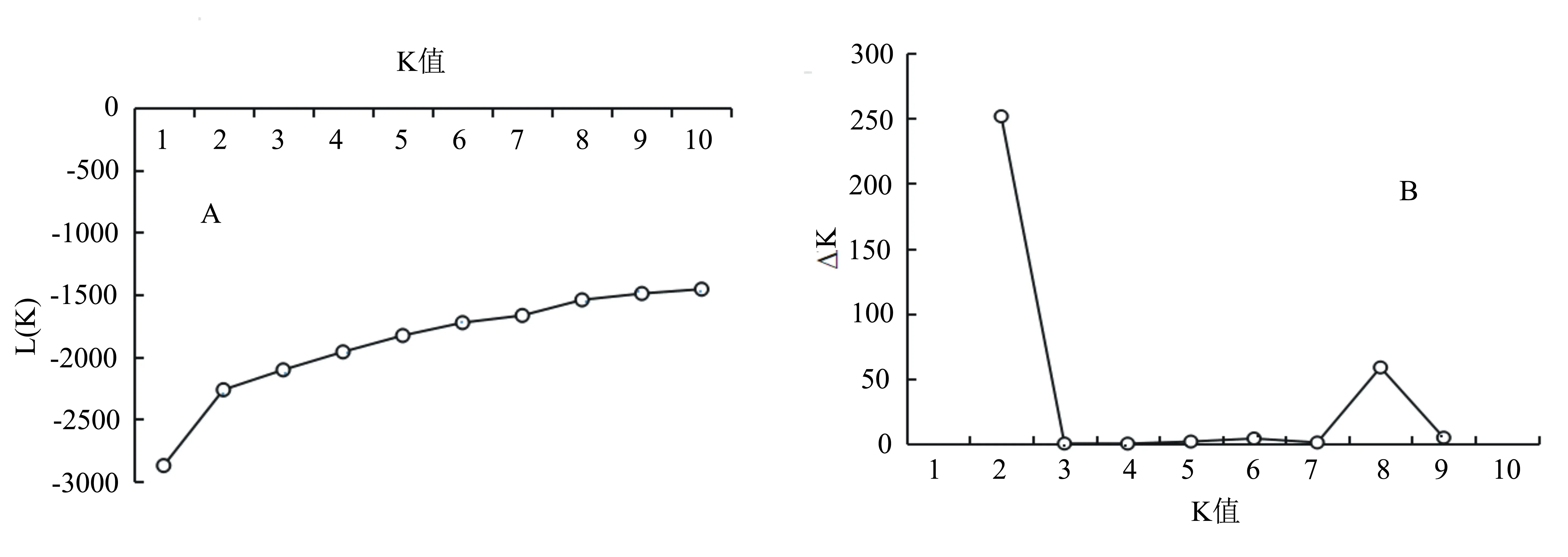
A为78份核桃资源的L(K)与K值关系图;B为ΔK与K值关系图。图3 78份核桃资源的L(K)、ΔK与K值关系图

图4 当K=2时78份核桃资源的群体结构
3 结论与讨论
不同物种适应新环境的基础是具备一定的遗传多样性,遗传多样性水平越高,该物种或种群对新环境的适应能力就越强[14-15]。目前遗传多样性的研究已从传统的形态学标记研究向分子标记研究转变,利用分子标记技术对多种核桃资源进行品种间的鉴定与分类不断取得进展。中国是核桃起源地之一,栽培历史悠久,种质资源丰富,核桃因其较好的食用价值、经济价值被推荐为世界各地人类健康饮食的重要资源,受到了广泛的欢迎,但中国核桃产业良种化程度不高,品种、类型混杂,难以区分。利用分子标记技术对一定群体内的核桃种质资源进行遗传特性分析,对于种质资源的区分、评价和保存及优良种质的选育具有重要的意义[16]。Wang H等[10]利用SSR标记揭示了西藏地区核桃及泡核桃的遗传多样性和杂种优势,发现西藏核桃群体具有平均水平的遗传多样性和大量的稀有等位基因。周于波等利用SSR技术,对四川29份核桃良种资源进行了遗传多样性分析和聚类分析,增强了品种间的区分能力[1]。与前人研究相比,本研究所选78份试材大部分为农家品种资源,一般来说,地方性品种对自生境适应性较强,对分散于全国各地的核桃农家品种资源进行遗传多样性分析,了解农家品种现有资源的多样性及其结构,可为遗传资源的保护和优良种质的筛选提供重要信息。
本研究利用25对SSR引物对78份核桃资源进行遗传多样性研究,最终筛选出8对引物,可加强对78份资源的鉴别。从78份供试材料共扩增出55个多态性条带,平均每对引物扩增出6.875个多态性位点。杨本芸等[17]利用10对引物对21份核桃品种进行扩增,每对引物平均扩增出6.7个多态性位点,多态性位点的相同与否可能与引物本身的多态性有关。78个不同品种间的遗传相似系数变化范围在0.605~1.000之间,而遗传相似系数可作为判断品种间亲缘关系远近的标准[1],本研究品种间遗传相似系数较大,遗传距离较小,平均遗传距离为0.23,由此说明该研究所采用的78份资源之间亲缘关系较为接近,遗传背景较为相似。这与陈良华等[18]对四川核桃遗传多样性的研究中野生核桃群体亲缘关系相近的结果相似。基于遗传相似系数,对78份材料进行UPGMA聚类分析,在遗传系数为0.704时,将群体分为5部分,分析发现栽培种与农家品种以及不同地理来源的核桃品种界限不明显,存在相互渗透的现象,也说明遗传聚类组的划分与地理来源不完全相关。姚明哲等[19]对江北地区茶树资源进行研究发现也存在该现象,四川的茶树品种被聚类到重庆茶树群体中。此外利用Structure 2.2软件对核桃群体进行遗传结构分析,发现最佳亚群体数为2,群体结构并不复杂,但存在相互渗透现象,可能存在种质的遗传变异或者基因交流[20]。
种质资源的研究是育种工作的一个重要基础,正确评价核桃种质资源的遗传多样性对其种质资源的创新具有重要意义。利用分子标记技术,同时结合形态学标记等方法对不同核桃品种的遗传多样性进行系统的研究与评价,可为培育核桃优良新品种、提高核桃现有种质资源的品质及抗性提供重要的科学理论依据。本研究78份核桃资源间的遗传关系相似,大部分品种间亲缘关系较为接近,因此应加强品种资源的有效利用,加强材料共享与交流,利用分子标记技术辅助育种,提高育种效率。
