不良环境解磷微生物研究进展
2020-07-22王雪郦张芮瑞周少奇邱树毅
王雪郦,张芮瑞,周少奇,邱树毅
(1.贵州大学 贵州省发酵工程与生物制药重点实验室,贵州 贵阳 550025;2.贵州大学 酿酒与食品工程学院,贵州 贵阳 550025;3.贵州大学 生命科学学院,贵州 贵阳 550025;4.贵州科学院,贵州 贵阳 550001)
磷作为第二大重要的植物营养元素,在植物的细胞分裂、能量转移、信号传导、核酸合成以及光合作用等过程中发挥着重要作用[1-2]。据统计,我国缺磷土壤面积已占总耕地面积的2/3以上,土壤中有效磷平均含量仅有0.1%,严重影响植物生长和作物产量[3-4]。磷肥虽在一定程度上满足了作物生长对磷元素的需求,却易因养分流失出现肥效下降、土壤养分比例失调、土壤板结及退化等问题[5]。事实上,磷素的循环受多种因素影响,其中,解磷微生物(Phosphate solubilizing microorganism,PSM)的作用能加速土壤中难溶性磷酸盐的转化。因此,利用PSM制备土壤改良剂、生物肥料等,不仅能提高土壤有效磷的利用率、增加农作物的产量,还能有效减少磷素的流失[6]。目前,部分分离自土壤的PSM已投入实际应用,且效果显著。如洋葱伯克霍尔德菌(Burkholderiacepacia)作为生物肥料可以显著提高番茄、马铃薯、洋葱等作物的产量[7];巴西青霉菌(Penicilliumbilaii)对小麦等谷物类作物促生长作用明显,能有效减少化学磷肥的施加[8]。
李露莉等[9-12],以贵州磷化工企业加工过程中产生的中、低品位磷矿和尾矿渣为研究对象,借助PSM对难溶性磷的转化作用,提高中、低品位磷矿和尾矿渣的综合利用率。最初,从磷矿区土壤中筛选分离出1株黑曲霉(Aspergillusniger),经研究发现,该菌株对磷矿粉具有较好的解磷性能。此后陆续从磷矿区土壤中筛选分离得到普城沙雷氏菌(Serratiaplymuthica)、嗜麦芽窄食单胞菌(Stenotrophomonasmaltophilia)、泡囊短波单胞菌(Brevundimonasvesicularis)3株解磷细菌(Phosphate solubilizing bacteria,PSB)[13-14],将其制成复合菌液后应用于好氧堆肥发酵工艺,最终制得符合NY 525—2012的有机肥产品。然而,在好氧堆肥发酵阶段发现,复合菌液在高温堆肥过程中,因受高温环境的胁迫作用,其解磷能力并不理想。这一现象在KHAN等[15]的研究中也曾提到,地球上的环境条件变化多端,PSM的生长代谢常受到诸如温度、pH值、环境压力[1]等因子的胁迫作用。环境因子的变化一方面可能引起PSM发生正向反馈,加速难溶性磷的转化,另一方面PSM可能为维持自身稳定发生负向反馈。虽然目前已经从各类环境中分离筛选得到大量PSM,但绝大部分都只能适用于常规的生态环境,而对于不良环境下PSM的相关表征报道较少。所以,当前面临的真正难题是挖掘一类可以在极端生态系统下仍能解磷,并具有显著促植物生长活性的新型PSM,以解决不良土壤环境中的农业生产以及环境维护等问题。不良环境PSM能突破各种不良环境因素的限制,实现难溶性磷的有效转化。基于上述理论,国内外学者就不良环境PSM展开研究,通过自然环境的优胜劣汰原则或人造环境的选择进化作用,筛选分离出能在特定不良环境下生长的PSM。本研究重点阐述耐寒、耐高温、耐盐PSM的研究现状,分析PSM在热带地区、高寒地区的农业生产、在盐碱地区土壤修复中的研究情况,及多种特定环境PSM的研究现状,就不良环境PSM的未来研究方向进行展望。
1 典型不良环境PSM的种类
1.1 耐寒PSM
耐寒PSM,即在低温环境下仍能保持较高解磷性能的微生物。在世界上的许多山区,常年的低温环境严重限制了土壤肥力的发挥,研究耐寒PSM目的之一是选择合适的生物接种剂,用于高海拔山区的土壤,耐寒PSM的应用能为常年处于低温的土壤提供更多的有效磷,促进作物生长。由于我国以亚热带、温带气候为主,少有高寒地区,故国内有关耐寒PSM的研究鲜有报道,而国外耐寒PSM的研究主要集中在印度喜马拉雅地区和安第斯山脉等高寒山区。早在20世纪90年代中后期MISHRA等[16]通过1-甲基-3-硝基-1-亚硝基胍(NTG)对3株荧光假单胞菌(Pseudomonasfluorescens)GRS1、PRS9和ATCC13525进行诱变,得到其对应的耐寒突变体。DAS等[17]证实,上述耐寒突变体在10 ℃时解磷活力维持在较高水平,且解磷能力均高于各自对应的野生型。TRIVEDI等[18]对从印度喜马拉雅地区玉米根际土壤中分离出的波状假单胞菌(Pseudomonascorrugata)进行亚硝基胍诱变,在115个突变体中得到2株具有较高解磷能力的耐寒突变体,并证明其在4~28 ℃下拥有良好的解磷能力,最适生长温度为21 ℃。上述研究表明,通过传统诱变技术处理PSM,所得突变菌株可显著改善植物在较低温度下对土壤磷酸盐的利用,但是从商业菌种的生产角度出发,既要考虑菌株的解磷能力,还要考虑菌株的遗传稳定性,所以大部分学者仍采用保守的自然进化方法从原始环境中筛选耐寒PSM。
迄今从高山及亚高山地区的天然土壤中分离出来的耐寒PSB主要包括荧光假单胞菌、恶臭假单胞菌(Pseudomonasputida)、卢氏假单胞菌(Pseudomonaslurida)等假单胞菌属和枯草芽孢杆菌(Bacillussubtilis)、巨大芽孢杆菌(Bacillusmegaterium)等芽孢杆菌属[15,19]。此外还存在个别其他菌属的耐寒PSM,如SELVAKUMAR等[20]对从喜马拉雅地区分离得到的分散泛菌(Pantoeadispersa)的研究表明,该菌能显著提高小麦幼苗的发芽率、植物生物量和营养摄取量。
耐寒解磷真菌(Phosphate solubilizing fungus,PSF)方面,据早期相关报道,学者们先后从印度喜马拉雅地区和北极等地区的土壤中分离得到产磷酸酶的耐寒PSF,主要包括曲霉菌(Aspergillus)、青霉菌(Penicillium)、镰刀真菌(Fusarium)及脉孢菌(Neurospora)等。 2008年,PANDEY等[21]从印度喜马拉雅地区土壤中分离得到的246株真菌中,发现8种青霉菌属耐寒PSF。其中,桔青霉(Penicilliumcitrinum)等对温度、pH值及盐含量表现出较为广泛的耐受性,该团队后续从印度喜马拉雅地区不同酸性土壤中分离出10种耐寒解磷的曲霉菌种,分别研究其在9、14、21、28 ℃条件下的溶磷情况,结论表明,黑曲霉的解磷性能最优[22]。随后,黑曲霉在难溶性磷转化方面的贡献度也得到SINGH等[23]的进一步证实。而RINU等[24]发现,黑曲霉、灰绿曲霉(Aspergillusglaucus)及聚多曲霉(Aspergillussydowii)除了能溶解磷酸钙外,还能溶解磷酸铝及磷酸铁,其中,以黑曲霉的溶磷效率最佳。于是,在一段时间里大部分耐寒PSF的研究重点是溶磷效果最佳的黑曲霉,而对于在低温环境下解磷活性较低但却能实现持续解磷的真菌关注较少。有研究证明,黑曲霉在14 ℃下对磷酸三钙(TCP)的溶解量最高可达1 394 μg/mL,而从第35天开始其解磷能力逐渐下降;相反,灰绿曲霉在同等条件下虽最高解磷量不及前者,但直至第42天其解磷量仍呈上升趋势,说明其持续解磷作用时间明显长于前者。黑曲霉是目前报道中所提及的最优耐寒PSM,但考虑到低温生态系统的特殊性,因此,诸如灰绿曲霉等此类能实现低温环境持续解磷的耐寒PSM同样值得关注。现分离筛选所得的耐寒PSF已取得了良好的应用成果,如我国农业农村部环境保护科研监测所从吉林省延吉市安图县大豆根瘤中,分离得到了耐寒解磷固氮微生物——中华根瘤菌ATHC,并利用其制备固态生物菌剂应用于大豆的温室盆栽及田间小区试验,结果表明,该菌剂能增加供试大豆根系结瘤数量,提高大豆地下部与地上部生物量,显著增加供试大豆产量[25]。
现阶段,对于耐寒PSF的研究不仅停留在特定环境中PSF的筛选及解磷性能方面,还有学者就耐寒PSF的作用机制进行探索。GAWAS-SAKHALKAR等[26]从北极土壤中分离得到56株真菌,其中9株为耐寒PSF。经研究发现,调控耐寒PSF实现低温解磷性能的是以细胞内酸性磷酸酶为主的磷酸酶,以桔青霉PG162产细胞内酸性磷酸酶最优。磷酸酶的分泌对PSM的解磷作用有直接影响,所以不仅是针对耐寒PSF,就整个耐寒PSM而言,就其体内发生的酶促反应类型以及互作机制进行研究,对寒冷环境下解磷机制的阐述有着重要意义。
1.2 耐高温PSM
在夏季土壤温度高达50 ℃的热带地区,普通常温PSM易因酶变性导致新陈代谢变缓,当超过一定高温时,甚至可直接使微生物组织结构损坏,所以迫切需要开发能适应高温条件的PSM。耐高温PSM是研究较早的不良环境PSM,GAIND等[27]于1991年从高达45 ℃的高温环境中筛选出枯草芽孢杆菌、环状芽孢杆菌(Bacilluscirculans)及黑曲霉3株具有良好解磷性能的菌株,并在以绿豆为研究对象的生物试验中发现,枯草芽孢杆菌促生效果最优。随后MAHESWAR等[28]就耐高温PSM蜡状芽孢杆菌(Bacilluscereus)和枯草芽孢杆菌的最适生长温度进行研究,结果发现,上述菌株在30~45 ℃具有较高的解磷活力。CHANG等[29]从堆肥原料中分离出3株耐高温PSM,其中,作为优势菌群的斯密氏芽孢杆菌(Bacillussmithii)具有最优的解磷性能,3株耐高温PSM的解磷能力由高到低依次是斯密氏芽孢杆菌、凝结芽孢杆菌(Bacilluscoagulans)、地衣芽孢杆菌(Bacilluslicheniformis)。
耐高温PSM在我国主要应用于高温堆肥,将筛选驯化后制成的耐高温菌剂接种于含磷固体废弃物的好氧堆肥中,借助高温PSM等多种微生物的降解作用促使堆肥快速腐熟,实现含磷固体废弃物的资源化利用。既在一定程度上缓解我国磷肥资源紧缺以及环境污染和生态破坏的压力,又达到生物资源回收利用和可持续发展的目的。张芮瑞等[30]从白酒酒糟堆肥样品中分离得到5株耐高温PSM,包含2株耐高温PSB和3株耐高温PSF,均能耐受35~60 ℃的高温,且50 ℃下解磷性能最佳。李鸣晓等[31]从堆肥环境中筛选并驯化得到5株耐高温PSM,经研究发现,其适宜生长温度为40~50 ℃。魏自民等[32]在此基础上对其解磷性能进行验证,研究发现,高温条件下肥料中有效磷含量有所增加,证明上述耐高温PSM对高温环境具有正向反馈作用。杨天学等[33]从添加磷矿粉的高温堆肥样品中分离筛选到2株具有较高解无机磷性能的耐高温PSM,分别为软化芽孢杆菌(Bacillusmacerans)和巨大芽孢杆菌。目前研究所分离得到的耐高温PSM多以芽孢杆菌属的细菌为主,这可能与该类菌酶系统能够耐受较高温度的能力有关。符玲[34]从农业废弃物堆肥中筛选到1株毕赤酵母(Pichiafarinosa),其可在45 ℃高温条件下实现对难溶性磷的转化。基于微生物协同作用机制的考虑,赵霞[35]选取解磷能力较强的3株耐高温PSM(P3、P4、P5),将其按不同比例混合制成复合菌剂,在相同条件下复合菌剂的最高解磷量为257.93 μg/mL,显著高于各单株菌的解磷能力。在实际应用中,优势微生物的最适生长温度变化差异较大,将耐受不同温度的耐高温PSM经复配后制成复合菌剂,既能够保证环境中有稳定的有效磷,又可以为高温堆肥以及新产品的研发起到促进作用。
1.3 耐盐PSM
全世界范围内,土壤含盐量均是限制作物生长最主要的非生物胁迫因素之一[36]。我国的土壤盐碱化问题也极为突出,国内各类盐碱化土壤总面积多达9 900万hm2[37],开发利用盐碱土壤对我国农业生态系统的可持续发展具有重要意义。传统治理手段包括物理排碱法或化学改碱法,效果均不理想,但若利用耐盐PSM的产酸解磷机制,不仅可降低盐碱土壤pH值,还能提高其有效磷含量。所以,为进一步提高盐碱土壤的改良效果,促进盐碱地植被生长,达到改善盐碱地生态环境的目的,迫切需要对高效耐盐PSM进行研究。
RES等[38]从土壤样品中分离筛选出1株枯草芽孢杆菌,该菌可耐受高达2.2%的NaCl,且在含盐土壤中促玉米生长作用明显。SONI等[39]从含盐土壤中分离出84株耐盐菌株,其中巨大芽孢杆菌A12ag在盐含量达到60 mmol/L时还能显著提高番茄种子的发芽率,田间试验证明,其对番茄促生作用显著。
刘长霞等[40]从海滨盐碱土壤中筛选出4株耐盐解无机磷真菌,发现PSF的生物量和解磷能力随NaCl含量的增加呈下降趋势。李学平等[41]也得出相似结论,塔滨曲霉菌(Aspergillustubingensis)解磷能力随发酵液盐含量的升高而降低,说明耐盐PSM在NaCl含量到某一阈值后出现了负向反馈。现有数据表明,大量耐盐PSM对NaCl含量的耐受能力处于2%~10%,少量耐盐PSM耐受能力高于10%,如范延辉等[42]从黄河三角洲盐碱土壤中分离得到的草酸青霉(Penicilliumoxalicum),最高可耐受12.5%的NaCl,ZHU等[43]从中国东海岸大桥盐湖沉积物中分离出的1株盐单胞菌科的Kushneriasp.,最高可耐受20%的NaCl。上述菌株可作为耐盐PSM或作为可用于盐碱地区生物肥料的潜在菌源。
耐盐PSM在低盐胁迫下发挥溶磷能力,向处于低盐抑制环境中的植物提供有效磷,有效促进盐碱化土壤的增肥及修复,提高盐碱障碍耕地作物产量。如黄渤海平原部分盐碱地区已将某些耐盐PSM制成微生物菌剂或微生物化肥投入盐碱地的改善治理中,并已取得良好效果。然而,通过文献对比分析发现,大量耐盐PSM的研究多采用NaCl作为限制因子,仅少量研究选用CaCl2作为限制因子。实际上,PSM对上述两者的耐受能力差异较大,如陆洪省等[44]研究发现,芽孢杆菌SKDJP-1对NaCl耐受能力为3%~5%,但对CaCl2的耐受能力仅为1%左右。此外,不同种类耐盐PSM的解磷能力及其对盐碱条件的耐受能力存在较大差异,所以盐碱土壤的其他条件(如有机质的含量、不同盐碱土的盐分构成等)对耐盐PSM的影响,以及耐盐PSM对盐碱地区植物的促生机制和耐盐PSM的代谢调控机制值得关注和研究。
2 不良环境PSM的研究热点
2.1 分离筛选
结合张芮瑞等[30]高温PSM的筛选工作,得出不良环境PSM的常规分离筛选策略,即先从特定的自然环境或人为环境中选取合适的待分离样品(常为土壤样品),常用的筛选方法是在缺磷的合成培养基中加入控制磷源,初步分离出不同的不良环境PSM。由于磷可大致分为无机磷和有机磷,所以分离解无机磷微生物时常选择TCP或磷矿粉(也有部分研究选择磷酸铝以及磷酸铁)为控制磷源,而分离解有机磷微生物时通常选择卵磷脂为控制磷源。通过选择培养基初筛出具有一定解磷、溶磷能力的菌株,再经多次复筛纯化后即可得到PSM菌株,最后按照常规的形态学、分子生物学方法对菌株进行种属的鉴定及命名。
近期笔者以酱香型白酒丢糟和磷尾矿粉为主要原料进行好氧堆肥,分别从45、50、55 ℃堆肥样品中筛选出耐高温PSM。但在耐高温PSM筛选过程中发现,PSM固态培养时溶磷圈的大小与自身解磷能力并无显著关系,该结论与郭艺鹏[45]的试验结论相似(图1)。D/d(溶磷圈直径/菌落直径)值最大的P12菌株总解磷量并非最高,相反D/d值最小的P7菌株解磷量最高,达到349.10 mg/L。甚至有研究表明,许多分离菌株在固体培养基上不产生任何可见的透明圈,但仍可溶解液体培养基中的难溶性磷源[15]。由此说明,在不良环境PSM筛选过程中,不能仅通过固态溶磷圈的大小或有无作为判断PSM解磷能力高低。
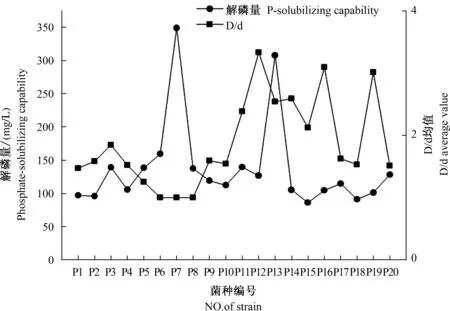
图1 PSM P1—P20菌株的解磷能力
目前,有关不良环境PSM新菌种的分离筛选研究主要集中在待分离对象的差异性方面,选取特定环境中具有代表性的不同样品,从中筛选出更多新的高效不良环境PSM。然而,现阶段通过自然筛选的菌种可能在解磷量等特性方面仍然不够理想,为了获得新的高质PSM资源,可考虑常压室温等离子体诱变(ARTP)等新兴技术,ARTP技术克服了传统诱变技术的诸多弊端,具有成本低、操作方便,能有效造成DNA多样性的损伤、突变率高,并易获得遗传稳定性良好的突变株等优点,该技术目前在某些领域的微生物、植物甚至动物方面的诱变育种中已经取得了良好成效,利用ARTP技术已经成功选育了包括高产头孢菌素C的突变株[46]以及耐高温性能的酿酒酵母[47]在内的诸多工业微生物生产菌,但该技术在不良环境PSM筛选方面的应用目前暂无报道。
2.2 解磷机制
不良环境PSM种类繁多,解磷机制不尽相同且较为复杂,而从现有报道来看,针对不良环境PSM解磷机制的研究尚浅。以普适PSM解磷机制的相关研究内容整理如表1。
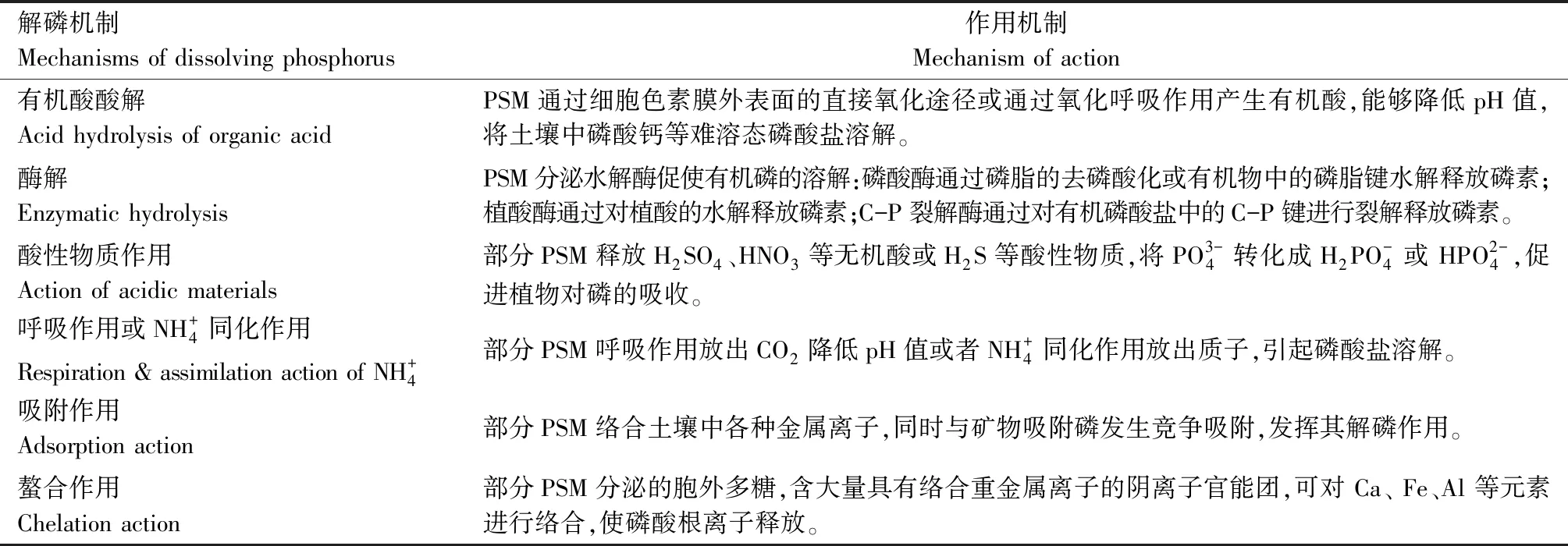
表1 PSM解磷及作用机制

目前,大量解磷机制的研究都集中围绕其中1~2种机制来进行,且多采用较为传统的分析手段,研究还不够全面,不能完整反映微生物的解磷机制。对于大家公认的有机酸解磷机制,在涉及不同菌株的相关试验时仍存在有尚未研究清楚的结论,如YADAV等[55]曾利用HPLC分析PSM菌株BISR-HY65发酵上清液中的有机酸,研究发现,除早前已有报道的柠檬酸、葡萄糖酸、苹果酸和甲酸外,还存在3种未知酸,而这些未知酸可能与该菌株的难溶性无机磷增溶有关。由此可见,不同的PSM菌株产酸解磷机制各有差异,后续应做大量的类比研究,探究其中规律或建立相应的产酸数据库。
现阶段对于不良环境PSM解磷机制的研究热点仍集中于阐述各自与解磷相关的功能产物和代谢途径,还未能将不同种类的解磷机制进行对照研究。实际上,研究各类不良环境PSM作用机制的异同点,能为新型不良环境PSM的挖掘或构建提供思路。
2.3 微生物间相互作用
李阜棣[56]曾提到外源微生物介入土壤必须经过与土著微生物生态位的竞争(包括氧气、空间等),才能在根际定殖并发挥作用。PSM作为外来添加的菌株,将与土壤环境中的土著微生物群落及其他外来微生物群落(如解钾、固氮微生物)发生诸如共生、寄生、拮抗等作用,可能会导致原始的微生态环境和微生物群落结构发生变化,而不同的微生物群落结构将会对溶磷过程产生间接影响,如微生物群落组成的变化会导致微生物代谢产物有机酸的类型和数量发生变化,从而引起正向或负向的反馈作用。所以在研究不良环境PSM相互作用时,一定要重视其与整个生态环境的关系。现阶段的研究一般分别从土壤生物化学、微生物生理生化和基因表达的不同层面出发,对菌株间相互作用的具体过程以及作用机制进行系统研究。PANDEY等[19]将荧光假单胞菌B0应用于玉米,结果发现,玉米植株生长量增长明显,根际的细菌数量也显著提升,但根际真菌数量却受到明显抑制,对微生物间相互作用的深入研究,可以进一步提高PSM的实际应用效果。张林[57]曾研究过丛枝菌根真菌(Arbuscularmycorrhizae,AM)与PSB的相互作用机制,发现AM菌丝分泌物可提高PSB分泌的磷酸酶活性,从而增强PSB活化有机磷的能力,而PSB又可以增强AM根外菌丝吸收和转运无机磷的能力,两者间呈现互利的协同关系。研究还发现,添加适量的无机磷甚至可以调节AM真菌、PSB间的互作关系,因此可通过调控农田生态系统中土著AM真菌来提高土壤有机磷的利用效率。
深入研究不良环境PSM与土著微生物间的相互作用及其机制,能进一步提高不良环境PSM对土壤中难溶性磷的转化率,有助于定向优化不良环境PSM的性能及适用条件,有利于新型复合PSM的研制和开发。
3 展望
本研究以耐寒、耐高温、耐盐PSM为代表,就不良环境PSM的研究策略到实际应用进行了详细阐述,证实了PSM在各类不良环境中的贡献度,如促进高寒等气候恶劣地区的农业发展,以及修复盐碱地生态环境等。实际上,除上述的3种不良环境PSM外,还存在耐旱、耐高压等不良环境PSM,以及能同时耐受多种不良环境胁迫的多功能不良环境PSM。如越南伯克霍尔德氏菌(Burkholderiavietnamiensis)能同时耐受包括低温、高盐在内的环境胁迫因素,再如王聪等[58]从西沙喜盐草根际沉积物中筛选到的星箭头菌(Burkholderiavietnamiensis)和海洋芽孢杆菌是同时具有抗高盐、耐强碱能力的固氮解磷功能菌。因多功能不良环境PSM具有适用范围广、使用便捷等优点,所以其研究将有助于混合微生物菌剂的研制,确保不良环境PSM在复杂多变的土壤环境中仍具有较高的磷转化能力。因此,加强多功能不良环境PSM的研究具有实际意义。
目前,有关不良环境PSM的研究主要侧重在微生物的分离筛选方面,研究深度仅限于解磷性能的分析、田间试验的应用等。但是,欲提高特定环境下微生物对难溶性磷的转化,需要挖掘出更多更高效的PSM,对解磷机制有更为全面的阐述。而就解磷机制的研究可以从基本生物学过程与代谢途径、功能蛋白、解磷基因等多个层面相结合,共同探索不良环境PSM的环境适应机制。笔者拟设计采用宏转录组技术(MT)分析不同条件下(如高温、常温)PSM中显著上调或下调的基因,并通过GO(Gene ontology)对其进行功能注释,利用群体感应(QS)分析揭示该环境下PSM群体效应的调控机制。此后将宏转录组技术与宏基因组(MG)数据结合,通过标准丰度(RPKM)进行物种、功能比较,利用MT/MG比值分析相对转录活性,探究PSM与环境的互作机制。甚至可将转录组和代谢组联合分析,从大量转录组信息中发掘差异基因,快速判断核心调控网络和关键候选基因,明确转录因子功能,阐述PSM响应非生物胁迫的应答机制,最终可在机制的研究基础上选择解磷基因,进行以应用为导向的理论研究和分子改造,将不良环境PSM的促解磷基因转入大肠杆菌、酵母菌等模式生物中,构建得到相应表型的生物体,使宿主解磷性能增强的同时,加快难溶性磷的转化速度,虽然这部分在常温PSM黑曲霉方面已有相关报道[59],但在模式菌株中的表达效果并不理想,需要进一步对其进行深入研究。
自然环境中存在着大量未开发的生物资源,现如今筛选得到的不良环境PSM只是冰山一角,为推广PSM的实际运用,需要根据多变的自然环境,进一步深度开发不良环境下的PSM资源。与此同时,还迫切需要开发一些简单的技术,用来大规模生产不良环境PSM及完善其运输体系,以便PSM能在不同不良环境中得到推广和使用。本研究详细阐述了耐高温、耐寒、耐盐3种不良环境PSM的研究现状,列举了其在农业生产、固废利用和生态治理等方面的应用,分析了现阶段不良环境PSM研究的不足,为未来多角度研究不良环境PSM提供了借鉴,为不良环境PSM及其相关生物肥料制品的研发提供新思路。
