壳寡糖在水产动物上的研究与应用
2020-07-09张义张志浩张继平
张义 张志浩 张继平

摘要:壳寡糖(COS)是近年来新开发的具有增强免疫力的生物活性物质,具有抗菌、增强免疫力、降血脂及抗肿瘤等多种生物活性,现已被广泛应用于食品及医药等领域。综述了壳寡糖主要生物学功能及其对水产动物生长性能、非特异性免疫及肠道微生物的影响,并对其影响机制进行了简要阐述,同时就其应用前景进行了展望,旨在为壳寡糖广泛应用于水产养殖业提供参考。
关键词:壳寡糖(COS);生长性能;免疫细胞;免疫酶;肠道微生物
中图分类号:S9 文献标识码:A
文章编号:0439-8114(2020)06-0005-05
DOI:10.14088/j.cnki.issn0439-8114.2020.06.001 开放科学(资源服务)标识码(OSID):
Research and application of chitosan in aquatic animals
ZHANG Yi,ZHANG Zhi-hao,ZHANG Ji-ping
(School of Life Science and Engineering, Foshan University,Foshan 528231,Guangdong,China)
Abstract: Chitosa(COS) is a newly developed biologically active substance with immuno potentiating effect. It has various biological activities such as antibacterial, immune enhancement, hypolipidemic and anti-tumor, and has been widely used in food and medicine fields. The main biological functions of chitosan and their effects on aquatic animal growth performance, non-specific immunity and intestinal microorganisms were reviewed, and the mechanism of their effects was briefly explained, the prospect of its application was prospected,in order to provide a reference for the wide application of chitosan in aquaculture industry.
Key words: chitosan(COS); growth performance; immune cells; immune enzymes; intestinal microbes
甲壳素是自然界最丰富的天然多糖,壳聚糖是其最重要的衍生物,甲壳素和壳聚糖的分子量高、溶解性差、黏度高,限制了其在许多领域的应用。壳寡糖(COS)是甲壳素和壳聚糖经过物理、化学或酶促聚解反应生成聚合度为2~20的低聚壳聚糖,是自然界大量存在的带正电的碱性氨基多糖。COS的分子链与母体相比要短很多,使得分子间不易形成较强的氢键,提高了COS在水中的溶解性,同时无细胞毒性,可以直接被肠道内上皮细胞吸收。COS具有广泛的生物活性,包括抗菌消炎、抗肿瘤、抗高血压、抗氧化、增强免疫力及调节肠道微生物结构等,由于其多方面优于母体聚合物,COS被广泛应用于各个领域[1,2]。有关COS在禽畜身上的应用已经取得显著成效,Huang等[3]在对肉鸡的研究中发现,日粮中添加壳寡糖能提高肉鸡的免疫水平,且添加量为100 mg/kg时效果最好。Zhou等[4]在对猪的研究中发现,日粮中添加2 000 mg/kg COS能够提高猪的生长性能,改善表观消化率,减轻猪的腹泻症状。COS作为一种纯天然的活性物质,在促进水产动物生长、增强免疫力及改善肠道微生物等方面具有较高的研究价值和广阔的应用前景,本文综述了COS的生物学功能及其作用机制,以期为这方面研究提供参考。
1 壳寡糖的促生长功能
1.1 壳寡糖在生长方面的应用研究
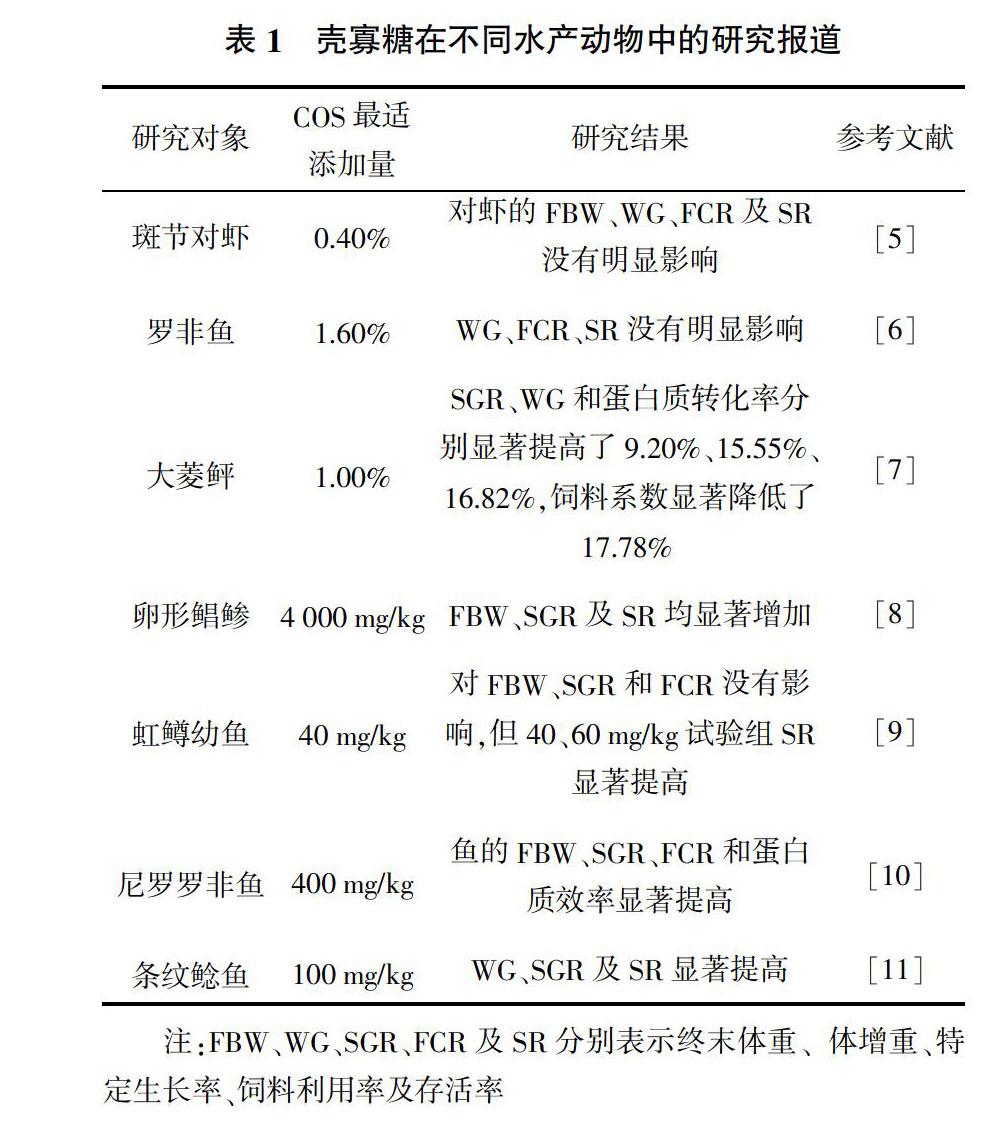
近几年来,COS在水产动物中的研究报道逐渐增多,在饲料中添加一定水平的COS,对斑节对虾、罗非鱼与卵形鲳鲹等水产动物的体增重、特定生长率、饲料转化率及成活率等生长性能有一定的影响。大多数研究表明,COS对水产动物的生长性能有明显的促进作用,且在不同的水产动物中有不同的适宜添加水平,具体的研究报道见表1。
1.2 壳寡糖对水产动物的促生长作用机制
肠道是鱼类消化与吸收营养物质的主要部位,也是鱼类最大的黏膜免疫器官,正常的肠道形态结构是营养物质吸收和肠道免疫的基础[12]。COS可通过改善肠道组织形态、缓解肠道炎症及降低应激反应来提高动物的生长性能,田娟等[13]研究发现,在日粮中分别添加0.30%和0.50%的COS,幼鱼前肠绒毛长度、宽度及密度显著增加,各添加组的幼鱼肠壁厚度明显减小;孙飞等[14]报道了COS添加量为0.02%時,异育银鲫肠道形态结构改善效果最为明显,肠道绒毛排列整齐均匀,与对照组相比,肠皱襞高度与肠壁厚度分别增加了86.84%和20.45%,肠皱襞宽度减少了12.18%。动物肠道绒毛高度、宽度、密度及肠皱襞高度的增加,有利于增加小肠对食物的接触面积,提高对营养物质的吸收能力。Qin等[6]还发现,COS可以显著减轻罗非鱼肠道炎症反应和应激反应,表现为促炎蛋白肿瘤坏死因子-a和应急反应蛋白热休克蛋白70的mRNA表达水平降低,转化生长因子TGF-b水平升高。
肠道内消化酶活性的高低决定了营养物质的分解程度,消化酶活性的增强有助于水产动物对养分的消化和吸收,对于一些无胃鱼(鲤科类)来说,对营养物质的分解更依赖于消化酶的消化作用。COS可通过提高肠道内胰蛋白酶、胰脂肪酶及胰淀粉酶的活性,促进水产动物对养分的吸收利用,从而提高水产动物的生长性能。潘金露[15]在对大菱鲆的研究中发现,饲料中添加0.50% COS时,肠道内胰蛋白酶和胰脂肪酶活性显著高于对照组,添加量为0.02%时,肠道胰淀粉酶活性显著高于对照组;Su等[16]在对红鳍东方鲀的研究中发现,随着COS水平的增加,肠道内胰蛋白酶和胰脂肪酶活性也随之上升,并在0.20% COS组达到最大值,与对照组差异显著,这与胡晓伟等[17]的研究结果一致。
矿物质与维生素的缺乏会引起水产动物产生营养性疾病,钙元素缺乏导致神经活动受到抑制,同时引起虾蟹壳软,缺铁性贫血,缺乏维生素C也会引起贫血,并导致脊椎及鳃软骨畸形、肌间出血、水肿等症状。饲料中添加一定水平的COS可以促进矿物质及维生素的吸收。COS因分子结构中含有-NH2和-OH等活性基团,是一种良好的网状载体,许多金属离子能与COS中的氨基N原子和羟基O原子形成配位键,从而生成一类具有独特环状结构的螯合物(COS-M),可以促进矿物质(钙、磷)吸收,而且可以提高微量元素(锰、硒)的吸收利用率,从而促进动物健康生长[18]。王卓[19]研究发现,COS与Cu离子形成螯合物,能显著提高鲫鱼肠道对铜的吸收量。Pallela等[20]还发现,COS还可以与维生素C形成配合物,提高维生素C的利用率。
动物的生长性能与肠道微生物存在密切联系,COS改善肠道环境的同时,也有利于机体对养分的消化吸收。COS本身可以作为促生长因子被有益菌(双歧杆菌)所利用,不仅能促进双歧杆菌的增殖,而且促进其产生B族维生素,促进肠道蠕动,提高蛋白质的吸收利用率[21];Lee等[22]研究也发现,COS对双岐杆菌和大多数乳酸杆菌的生长具有明显的益生作用,其本身可作为生长刺激因子被双岐杆菌和乳酸杆菌所吸收利用。乳酸菌能够定殖在肠道内,有效地抑制有害菌的生长,减少肠道内有害菌产生的毒素对水产动物的毒害,同时能促进蛋白质、单糖及矿物质(钙、镁)的吸收,合成B族维生素等大量有益物质。还有学者研究发现,COS对脂多糖(LPS)诱导的肠上皮细胞炎症反应及伴随的过氧化损伤具有保护作用等[23]。
2 壳寡糖的免疫调节功能
2.1 壳寡糖在免疫调节方面的研究与应用
已有研究表明,COS是一种安全有效的绿色饲料添加剂, 可作为饲料抗生素的替代物广泛应用于水产养殖中。有关学者在对大口鲈、太平洋对虾、虹鳟及糙海参等品种的研究中发现,在基础日粮中添加一定水平的COS,对水产动物免疫细胞活性、吞噬能力及血清免疫酶活性有明显的提升效果。有关COS对水生动物非特异性免疫力的研究报道见表2。
2.2 壳寡糖的免疫调节机制
鱼类是较低等的脊椎动物,先天免疫在鱼类的免疫防御中发挥着重要作用,鱼类的特异性免疫系统受环境(温度、生理状态)影响较大,特异性免疫应答反应进程较长,水生无脊椎动物由于缺乏真正的抗体和特异性免疫细胞,机体防御机制依赖于非特异性的细胞免疫,因此,水产动物抵抗疾病主要依赖先天免疫系统[31,32]。血液中免疫细胞数量和吞噬活性可以反映出水产动物机体的免疫状态,血清LZM、ACP、AKP、SOD及GSH-Px(谷胱甘肽过氧化物酶)等免疫酶是鱼类机体非特异性免疫反应的重要指标,其活性的升高表明机体的非特异性免疫力增强[33]。
免疫系统中的免疫细胞、免疫酶及免疫调节因子在水产动物抵抗病原菌入侵中起決定性作用。以往研究表明,COS对水产动物非特异性免疫具有良好的提升效果。COS可以刺激免疫细胞释放细胞因子,细胞因子具有调节非特异性免疫、特异性免疫、细胞生长以及损伤组织修复等多种功能,并形成复杂的细胞因子调节网络。曹秀明[34]研究发现,50~100 μg/mL的COS能显著提高巨噬细胞的吞噬能力,能刺激巨噬细胞产生NO、TNF-α等细胞因子;吴海明等[35]也报道了COS可以刺激巨噬细胞分泌IL-1β、TNF-α和IL-18等细胞因子,分泌的细胞因子又反作用于巨噬细胞和NK细胞,形成网状的反馈调节系统,进一步提高机体的免疫功能;COS分子结构中带有的活性基团与免疫细胞表面的受体结合而介导免疫调节,侯丽娜等[36]研究发现,COS激活巨噬细胞可能是经由巨噬细胞表面的甘露糖受体介导结合而实现的,并刺激巨噬细胞产生IL-1β;韩燕萍[37]也指出,壳寡糖激活巨噬细胞是通过壳寡糖与巨噬细胞之间结合、内吞及转运这一过程实现的,巨噬细胞膜表面的甘露糖受体在这一过程中发挥重要作用,甘露糖受体可选择性识别含有N-乙酰氨基葡萄糖基团的多糖。由于COS自身具有抗氧化活性,可作为一种抗氧化剂来增强机体免疫力,能通过清除部分自由基(·OH)、超氧阴离子自由基(O2-·)等活性氧自由基(ROS),抑制ROS通过脂质过氧化损伤细胞膜,进而破坏蛋白质和DNA的作用,达到缓解机体氧化损伤的目标[38]。COS还通过提高LZM、SOD、ACP、AKP、GSH-Px等酶活性来增强机体免疫力,目前有关COS对免疫酶活性影响的机理尚不清楚,据推测可能是促进免疫酶相关基因的表达。
3 壳寡糖对肠道微生物的调节功能
3.1 壳寡糖在调节肠道微生物方面的研究与应用
肠道微生物是机体必不可少的,在水产动物免疫和生长发育过程中发挥着重要作用。鱼类肠道微生物是由有益菌群(乳酸杆菌)和有害菌群(大肠杆菌)组成,有益菌群常定殖在肠道黏膜上皮表面,阻止有害菌在肠道黏膜组织上的附着,有害菌具有一定的致病性,会引起鱼类一些细菌性疾病的发生。有害菌数量的增加,会引起肠道内菌群平衡失调,影响正常的消化道环境[39]。日粮中添加一定水平的COS可以改善肠道微生物群落多样性,促进肠道内有益菌的增殖,减弱有害细菌对水产动物的毒害作用并缓解肠道炎症,增强肠道的生理功能。Su等[16]在对红鳍东方鲀的研究中发现,0.20%的COS使肠道内的有益菌(枯草芽孢杆菌)明显高于其他组,有害菌(大肠杆菌)明显少于其他组。田娟等[13]在对吉富罗非鱼的研究中发现也有类似现象。胡晓伟等[17]对花鲈的研究中,除与Su等[16]研究结果类似外,0.60%~1.00% COS显著降低了沙门氏菌的数量。蔡雪峰等[40]对虹鳟幼鱼的研究中发现,COS试验组的肠道细菌总数之间无显著差异,但优势菌群有所变化,且各试验组肠道微生物多样性降低,肠杆菌科和假单胞属减少了。Qin等[6]对杂交罗非鱼的研究中发现,1.60%和2.40% COS组显著改善了肠道菌群,并减轻了肠道炎症。
3.2 壳寡糖调节肠道微生物的作用机制
有益微生物可以调节非特异性的体液免疫,将失调的微生物群恢复到正常状态,还通过与肠道上皮细胞的相互作用和调节抗炎细胞因子的分泌来改变宿主的免疫反应,从而减少肠道炎症[41]。Lee等[22]研究发现COS对双歧杆菌和乳酸菌有益生作用,但其生长促进作用的机理尚不清楚。可能是动物肠道内吸收一定量的壳寡糖后,COS就会被有益菌群分解,从而产生大量的挥发性脂肪酸,双歧杆菌和乳酸杆菌等有益菌利用这些挥发性脂肪酸进行大量增殖。肠道有益微生物可以将COS作为发酵底物,诱导产生有机酸,从而降低肠道pH,pH降低可以抑制肠道病原体的繁殖[22,42,43]。Lillo等[44]认为COS的主要抗菌机制是能改变有害菌细胞膜的通透性,阻止外界营养物质的渗入,同时COS分子表面所带的正电荷与有害菌细胞膜上的负电荷位点相结合,降低了有害菌从外界摄取养分的代谢活性,最终导致细菌死亡,这与Choi等[45]的研究结果类似。Mei等[46]还发现,COS分子表面所带的正电荷能够与微生物细胞中存在的大分子负电荷组分结合,并吸收到微生物的细胞壁中,这种吸收导致其渗透到DNA并阻断RNA转录。COS是自然界中惟一带正电荷的碱性氨基多糖,游离氨基在酸性条件下能够被质子化,能够与有害菌表面的膜结合,使细胞质流失并干扰有害菌与肠壁的结合能力,促进病原菌随粪便排出体外,同时对真菌和微生物生长起到一定的抑制作用[47]。
4 展望
近年来,随着水产养殖业的规模空前扩大,养殖业在追求产量的同时也饱受水产疾病的困扰,最终导致药物滥用现象频发,为了提高水产品质量,目前国内外正在积极研发具有免疫增强作用的饲料添加剂。COS是由甲壳素和壳聚糖进一步降解生成的活性物质,具有多种生物学功能,原料来源广泛。目前在比目鱼、虹鳟、刺参及南美白对虾等品种上已有关于COS应用的研究报道,并具有较好的应用效果。虽然COS发挥生物学功能的分子机制尚不完全清楚,有待进一步深入研究,但COS开发作为具有免疫增强作用的绿色饲料添加剂,在水产养殖领域具有广阔的应用前景。
参考文献:
[1] LIAQAT F,ELTEM R. Chitooligosaccharides and their biological activities:A comprehensive review[J].Carbohyd Polym,2018, 184:243-259.
[2] MUANPRASAT C,CHATSUDTHIPONG V. Chitosan oligosaccharide:Biological activities and potential therapeutic applications[J].Pharmacol Ther,2017,170:80-97.
[3] HUANG R L,DENG Z Y,YANG C B,et al. Dietary oligochitosan supplementation enhances immune status of broilers[J].J Sci Food Agr,2010,87(1):153-159.
[4] ZHOU T X,CHO J H,KIM I H. Effects of supplementation of chito-oligosaccharide on the growth performance, nutrient digestibility, blood characteristics and appearance of diarrhea in weanling pigs[J].Livest Sci,2012,144(3):263-268.
[5] NIU J,LIN H,JIANG S,et al. Comparison of effect of chitin,chitosan,chitosan oligosaccharide and N-acetyl-d-glucosamine on growth performance,antioxidant defenses and oxidative stress status of Penaeus monodon[J].Aquaculture,2013,372-375:1-8.
[6] QIN C B,ZHANG Y T,LIU W S,et al. Effects of chito-oligosaccharides supplementation on growth performance,intestinal cytokine expression,autochthonous gut bacteria and disease resistance in hybrid tilapia Oreochromis niloticus ♀×Oreochromis aureus ♂[J].Fish shellfish immunology,2014,40(1):267-274.
[7] 蔡勝昌,张利民,张德瑞,等.壳寡糖与低聚木糖对大菱鲆(Scophthalmus maximus)幼鱼生长、体组成和血液生化指标的影响[J].渔业科学进展,2015,36(6):29-36.
[8] LIN S,MAO S,GUAN Y,et al. Dietary administration of chitooligosaccharides to enhance growth,innate immune response and disease resistance of Trachinotus ovatus[J].Fish shellfish immunology,2012,32(5):909-913.
[9] LIN L,CAI X F,HE C A,et al. Immune response, stress resistance and bacterial challenge in juvenile rainbow trouts Oncorhynchus mykiss fed diets containing chitosan-oligosaccharides.[J].Curr Zool,2009,55(6):416-422.
[10] MENG X,WANG J T,WAN W J,et al. Influence of low molecular weight chitooligosaccharides on growth performance and non-specific immune response in Nile tilapia Oreochromis niloticus[J].Aquacult Int,2017,25:1265-1277.
[11] NGUYEN N D,DANG P V,LE A Q,et al. Effect of oligochitosan and oligo-β-glucan supplementation on growth,innate immunity,and disease resistance of striped catfish (Pangasianodon hypophthalmus).[J].Biotechnol Appl Biochem,2017,64(4):564-571.
[12] 周小秋.營养物质与鱼肠道健康的关系[A].动物营养研究进展[C].北京:中国畜牧兽医学会动物营养学分会,2012.246-260.
[13] 田 娟,孙立威,文 华,等.壳寡糖对吉富罗非鱼幼鱼生长性能、前肠组织结构及肠道主要菌群的影响[J].中国水产科学, 2013,20(3):561-568.
[14] 孙 飞,何 杰,叶元土,等.壳寡糖对异育银鲫生长性能、肠道组织结构和非特异性免疫功能的影响[J].动物营养学报,2018,30(12):5153-5163.
[15] 潘金露.饲料中壳寡糖和褐藻酸寡糖对大菱鲆(Scophthalmus maximus)消化及肠道菌群的影响[D].大连:大连海洋大学, 2016.
[16] SU P,HAN Y,JIANG C,et al. Effects of chitosan-oligosaccharides on growth performance,digestive enzyme and intestinal bacterial flora of tiger puffer(Takifugu rubripes Temminck et Schlegel,1850)[J].J Appl Ichthyol,2017,33(3):458-467.
[17] 胡晓伟,上官静波,黎中宝,等.饲料中添加壳寡糖对花鲈(Lateolabrax japonicus)幼鱼的生长、消化和血清生化指标的影响[J].海洋学报,2018,40(2):69-76.
[18] 曾令智,孙大维,何昇鸿,等.壳寡糖微量元素螯合物在养殖领域中的应用[J].中国饲料,2018(11):66-70.
[19] 王 卓.壳寡糖及其螯合铜对动物铜吸收的比较研究[D].大连:辽宁师范大学,2009.
[20] PALLELA R,VENKATESAN J,KIM S K. Synthesis and radical scavenging properties of chitooligosaccharide-vitamin C complex[J].Journal of chitin & chitosan science,2014,2(1):55-61.
[21] 竺国芳,赵鲁杭.几丁寡糖和壳寡糖的研究进展[J].中国海洋药物,2000(1):43-46.
[22] LEE H W,PARK Y S,JUNG J S,et al. Chitosan oligosaccharides,dp 2-8,have prebiotic effect on the Bifidobacterium bifidium and Lactobacillus sp.[J].Anaerobe,2002,8(6):319-324.
[23] 杨院平,罗和生,仝巧云,等.壳寡糖对脂多糖诱导肠上皮细胞炎症的保护作用[J].中华生物医学工程杂志,2015(4):325-329.
[24] LIN S M,JIANG Y,CHEN Y J,et al. Effects of astragalus polysaccharides(APS) and chitooligosaccharides(COS) on growth, immune response and disease resistance of juvenile largemouth bass,Micropterus salmoides[J].Fish shellfish immunology,2017, 70:40-47.
[25] LIU L,YANG Z,ZHAO X,et al. Oligochitosan stimulated phagocytic activity of macrophages from blunt snout bream (Megalobrama amblycephala) associated with respiratory burst coupled with nitric oxide production[J].Dev Comp Immunol,2014,47(1):17-24.
[26] LIN S,MAO S,GUAN Y,et al. Effects of dietary chitosan oligosaccharides and Bacillus coagulans on the growth,innate immunity and resistance of koi(Cyprinus carpio koi)[J].Aquaculture,2012,342-343:36-41.
[27] LI R,YANG X,LI Q,et al. Enhanced immune response and resistance to edwardsiellosis following dietary chitooligosaccharide supplementation in the olive flounder(Paralichthys olivaceus)[J].Fish shellfish immunology,2015,47(1):74-78.
[28] HEO G,SHIN G. Effect of chitosan oligosaccharide on non-specific immune parameters in catfish(Silurus asotus)[J].Philipp J Vet Med,2011,48(1):22-26.
[29] RAHIMNEJAD S,YUAN X,WANG L,et al. Chitooligosaccharide supplementation in low-fish meal diets for Pacific white shrimp(Litopenaeus vannamei):Effects on growth,innate immunity,gut histology,and immune-related genes expression[J].Fish shellfish immunology,2018,80:405-415.
[30] 刘均玲,康静玮,符中环,等.壳寡糖对海南热带糙海参免疫相关酶的影响[J].海洋科学,2017,41(12):26-31.
[31] ALCORN S W,MURRAY A L,PASCHO R J. Effects of rearing temperature on immune functions in sockeye salmon (Oncorhynchus nerka)[J].Fish shellfish immunology,2002,12(4):303-334.
[32] CERENIUS L,S?魻DERH?魧LL K. 2-Crustacean immune responses and their implications for disease control[A].AUSTIN B.Infectious disease in aquaculture[M].Woodhead publishing limited,2012,69-87.
[33] 闫 飞,赵红月,刘来亭.壳寡糖在水产领域的应用开发[J].饲料研究,2015(7):51-56.
[34] 曹秀明.壳寡糖及衍生物抗肿瘤作用、免疫调节作用及其机制的研究[D].山东青岛:中国海洋大学,2010.
[35] 吴海明,华晓阳,何登全.壳寡糖抗肿瘤作用及免疫调节机理的研究[J].中华医学研究杂志,2005,5(8):729-731.
[36] 侯丽娜,赵鲁杭.壳寡糖结合并激活巨噬细胞机制的研究[J].中国医科大学学报,2006,35(2):124-127.
[37] 韩艳萍.膜受体介导的内吞在壳寡糖激活巨噬细胞中的作用[D].杭州:浙江大学,2005.
[38] 钟 佳,刘进辉,肖定福,等.壳寡糖及其衍生物的抗氧化活性[J].动物医学进展,2015,36(7):118-121.
[39] 宋增福,吴天星.鱼类肠道正常菌群研究进展[J]. 水产科学, 2007,26(8):471-474.
[40] 蔡雪峰,罗 琳,战文斌,等.壳寡糖对虹鳟幼鱼肠道菌群影响的研究[J].中国海洋大学学报(自然科学版),2006,36(4):606-610.
[41] G?魨 MEZ G D,BALC?魨 ZAR J L. A review on the interactions between gut microbiota and innate immunity of fish[J].Fems Immunol Med Mic,2010,52(2):145-154.
[42] LI X J,PIAO X S,KIM S W,et al. Effects of chito-oligosaccharide supplementation on performance, nutrient digestibility, and serum composition in broiler chickens[J].Poult Sci,2007,86(6):1107-1114.
[43] MIKKELSEN L L,JAKOBSEN M,JENSEN B B. Effects of dietary oligosaccharides on microbial diversity and fructo-oligosaccharide degrading bacteria in faeces of piglets post-weaning[J].Anim Feed Sci Tech,2003,109(1):133-150.
[44] LILLO L,ALARCON J,CABELLO G,et al. Antibacterial activity of chitooligosaccharides[J].Z Naturforsch C,2008,63(9-10):644-648.
[45] CHOI B K,KIM K Y,YOO Y J,et al. In vitro antimicrobial activity of a chitooligosaccharide mixture against Actinobacillus actinomycetemcomitans and Streptococcus mutans[J].Int J Antimicrob Ag,2001,18(6):553-557.
[46] MEI Y X,DAI X Y,YANG W,et al. Antifungal activity of chitooligosaccharides against the dermatophyte Trichophyton rubrum[J].Int J Biol Macromol,2015,77:330-335.
[47] 秦 朋,徐小龍,张 斌,等.海洋生物新材料壳寡糖在动物领域研究现状及应用前景[J].粮食与饲料工业,2018(2):54-58.
