香椿优树半同胞家系苗期生长变异与选择
2020-07-09李成勇索怀俊肖兴翠何芝然杨滨豪
李成勇, 索怀俊, 肖兴翠, 何芝然, 杨滨豪
1. 江安县林业和竹业局,四川 宜宾 644200;
2. 旺苍县国有林场,四川 广元 628200;
3. 四川省林业科学研究院,四川 成都 610081
香椿(Toona sinensis),是楝科香椿属落叶乔木,生长迅速,树干通直,木材色泽红润、花纹美丽、芳香、有“中国桃花心木”之称,是我国特有珍贵速生用材树种[1]。香椿作为用材林发展逐渐得到重视。选育出适合各区域的香椿良种,成为国内学者研究的重点之一。在遗传改良方面,国内外学者对各地种源苗期生长变异[2-5]、家系选择[6,7]及早期选择[8-11]方面进行了大量的研究,少部分学者已经开始进行无性系苗期选择[12]及早期选择[13]方面的研究。四川省2000 年开始把香椿作为优先发展的6 个珍贵用材树种之一,在川中丘陵区、川南地区优先发展,目前人工林面积已有5 万多hm2。四川香椿分布广泛,北至秦巴山区,南至凉山州。四川省香椿良种选育主要是从“十二五”开始,以四川省林科院为主的香椿育种团队先后进行了种质资源收集,对筠连、蓬溪种源的优树半同胞家系进行了苗期比较与选择[7],为香椿超级苗选择及无性系选育奠定了一定的基础,但苗期选择种源范围有限。本文以川北、川中及至川南香椿3 大主要分布区域优树的半同胞家系为研究对象,对1 年生容器苗的苗期生长与变异进行了分析,筛选出苗期优良家系,以期为后期的良种选育提供参考。
1 材料与方法
1.1 试验地概况
试验地位于泸州市泸县的玉蟾山 (105°23′E,29°09′N),海拔350 m~550 m,属亚热带湿润季风气候,年平均温度17.1 ℃,年最高温度39 ℃,年最低温度-1 ℃,年有效积温 5 300 ℃,年降雨量1 110 mm,无霜期320 d 左右。苗圃地的土壤为黄沙土。
1.2 试验材料及来源
参试的18 个香椿半同胞家系主要来源于四川香椿3 大主要分布区域:川北秦巴山区(朝天、万源)、川中丘陵区(蓬溪)以及川南地区(马边、合江、筠连),是四川省林科院在这些地方选择的香椿优树的半同胞家系种子,优树基本情况见表1。
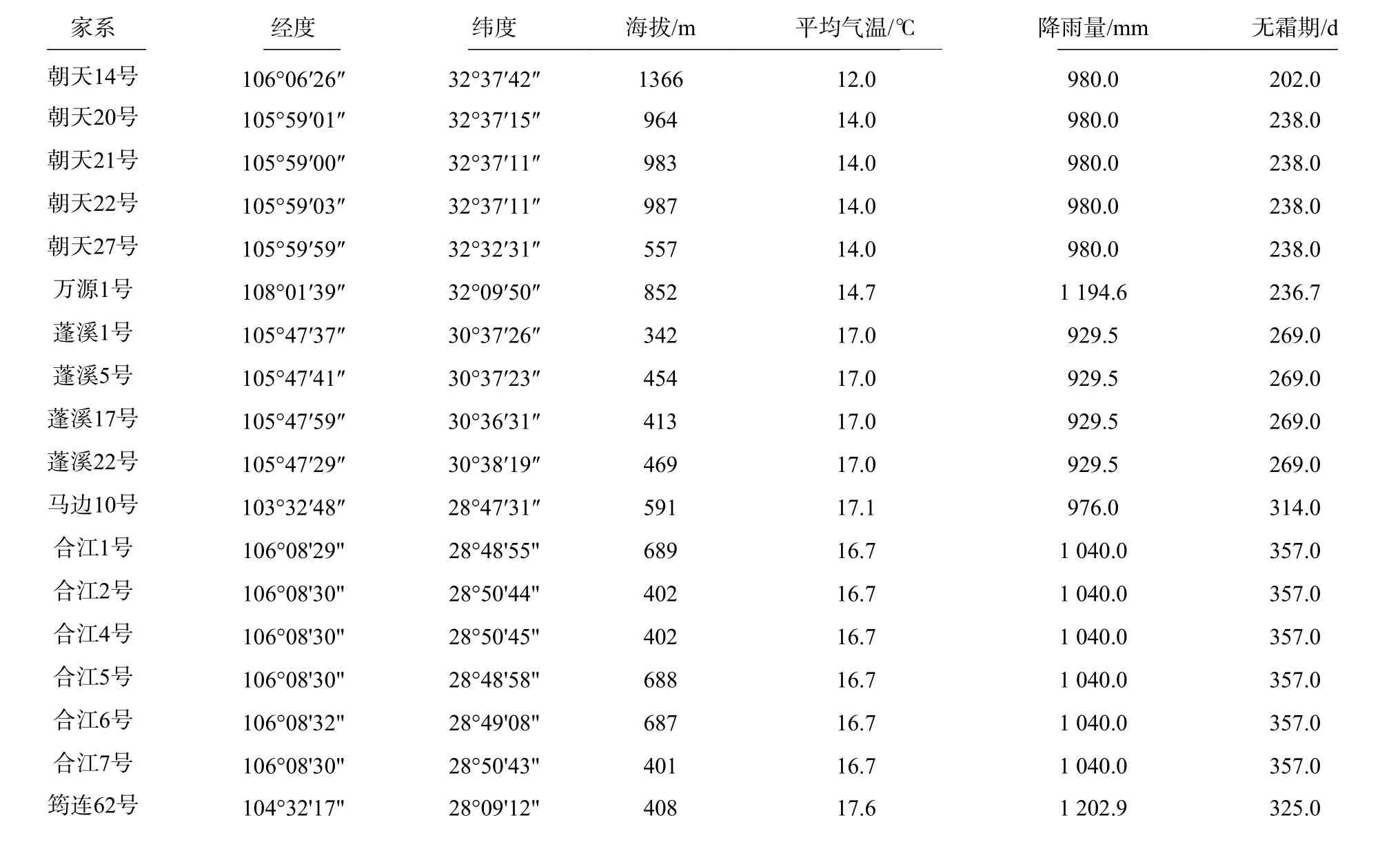
表1 香椿家系所在地理位置及气候条件Tab. 1 Geographical location and climatic conditions of Toona sinensis families
1.3 试验设计及育苗方法
2019 年3 月,对18 个香椿半同胞家系种子进行分家系播种育苗,每个家系育苗50 株/小区,3 次重复,随机区组排列。具体方法是将每个家系种子分别播种到整平的苗床培养芽苗,5 月份幼苗长到4~5 片真叶时,移栽到直径6 cm、高8 cm 的无纺布容器袋中,基质配比为泥炭4:椰糠3:土3;容器袋挨个紧密摆放在整平的地面,四周用土压实,移栽后盖遮阳网1 个月左右。之后采用相同的方法进行苗期管理,苗期管理参照《DB51/T 1156-2010 香椿育苗技术规程》[14]。
1.4 数据测量及分析
2019 年11 月苗木停止生长后,用游标卡尺测量地径,用直尺测量苗高,每个家系均测量中间30 株/小区,3 次重复。采用Excel10 对数据进行整理,并采用SPSS17.0 软件对数据进行方差分析及多重比较。遗传变异系数:C=S/X×100%,式中:S 为标准差;X 为某一性状的群体平均值;家系遗传力h2=(MSv-MSt)/MSv×100,式中:MSv 为处理间均方;MSt 为处理内均方[15];遗传增益△G=h2Si/X,现实增益G=(Xi-X)/X×100%,式中,h2为性状的遗传力,Si为家系选择差,Xi为个体平均数[16]。
2 结果与分析
2.1 不同家系生长与环境相关性分析
对不同家系的苗高和地径与家系所在地区的海拔、气候进行相关性分析,结果见表2。
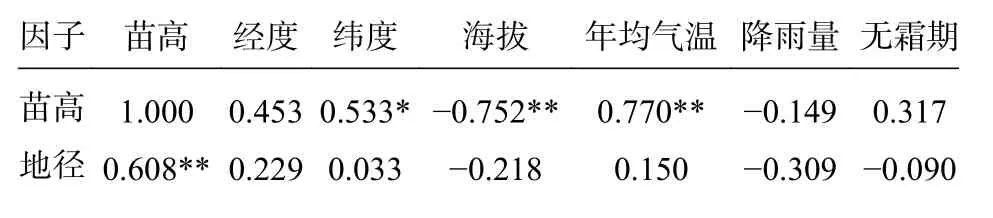
表2 香椿家系苗期生长与环境因子相关性分析Tab. 2 Correlation analysis of seedling growth and environmental factors of Toona sinensis families
由表2 可见,苗高和地径的相关性系数为0.608,两者之间有极显著(P<0.01)的相关性。
苗高在与产地地理位置上与纬度有显著的正相关性,即随纬度的增大而增大,与经度没有显著的相关性;与海拔有极显著的负相关性,即海拔越高,苗高越小;苗高在气候因子上与年均气温有极显著的正相关性,与降雨量和无霜期均没有显著的相关性。地径与产地地理位置及各气候因子均没有显著的相关性。
2.2 不同家系生长变异
对18 个香椿半同胞家系的苗高和地径进行方差分析,结果见表3。

表3 不同家系苗期方差分析Tab. 3 Variance analysis of different families at seedling stage
由表3 可见,18 个香椿半同胞家系间的地径和苗高均有极显著(P<0.01)差异,表明家系间变异较大,可进一步开展苗期优良家系选择。苗高和地径遗传力分别为0.939 和0.762,遗传力均较大,苗高的遗传力大于地径,表明苗高受遗传的控制比地径更大。
2.2.1 不同家系苗高生长变异
18 个香椿半同胞家系苗高生长与变异见表4。
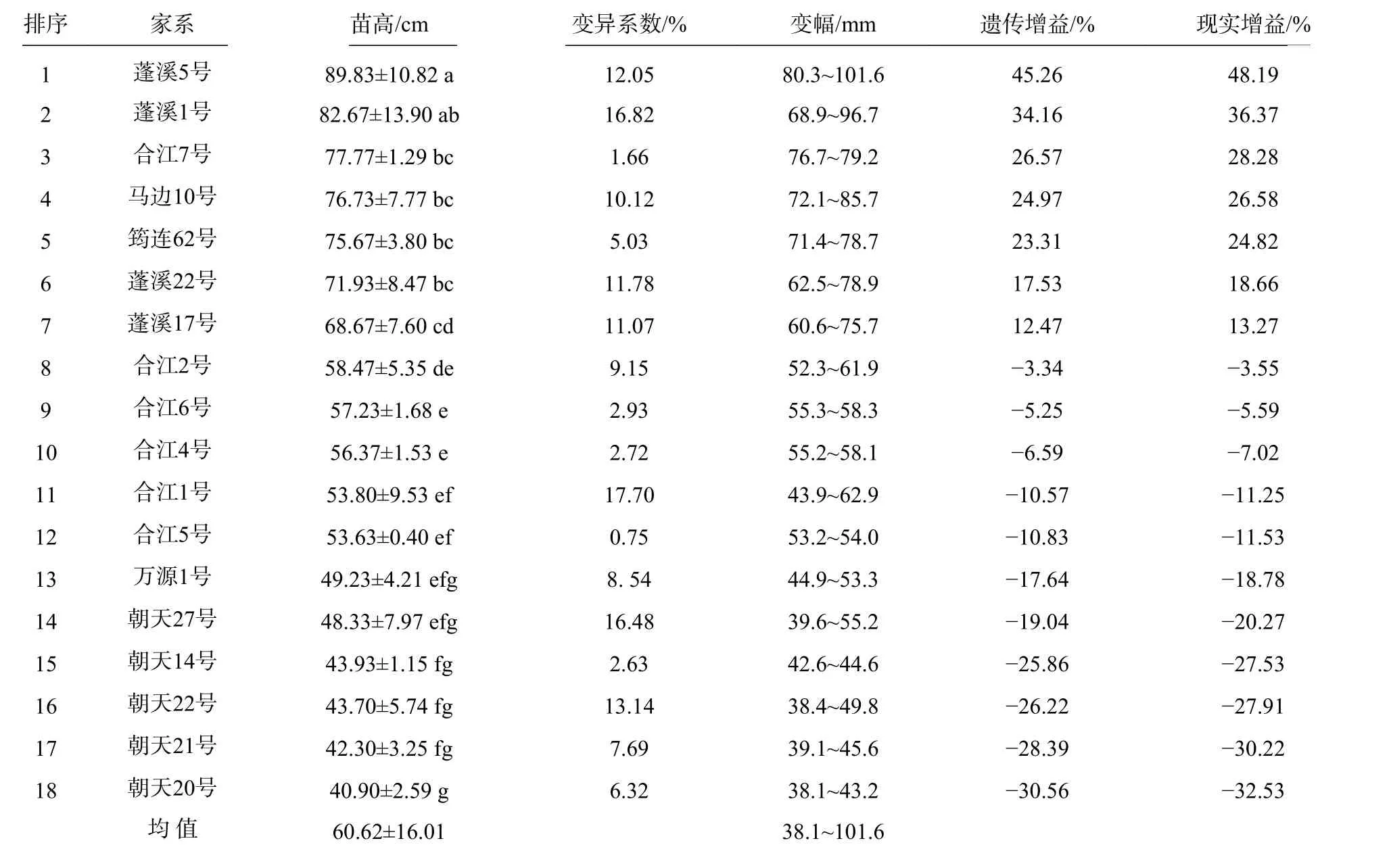
表4 不同家系苗高生长变异Tab. 4 Variation of seedling height growth in different families
由表4 可见,18 个香椿半同胞家系的平均苗高为60.62 cm,变幅为38.1~101.6 cm,超过平均值的家系有7 个,占家系总数的38.89%。蓬溪、合江、马边、筠连4 个地区的苗高高于万源、朝天的种源,在地理上呈现川中、川南高于川北的趋势。18个家系中以蓬溪5 号的苗高最大,苗高为89.83 cm,比平均值高48.19%,变幅为80.3~101.6 cm,遗传增益和现实增益分别为45.26%和48.19%,与蓬溪1 号间没有显著(P<0.05)差异,显著高于其余16 个家系;其次是蓬溪1 号,苗高为82.67 cm,变幅为68.9~96.7 cm,比平均值高36.37%,变幅为68.9~96.7 cm,遗传增益和现实增益分别为34.16%和36.37%,与蓬溪5 号、合江7 号、马边10 号、筠连62 号、蓬溪22 号5 个家系间有差异但差异不显著,显著高于苗高≤68.67 cm 的12 个家系。18 个家系中,蓬溪1 号、合江1 号和朝天27 号的变异系数大于15%,家系内苗高存在丰富的遗传变异,具有苗期选择的潜力,其余15 个家系的变异系数均小于15%,家系内苗高生长较为均与一致。
2.2.2 不同家系地径生长与变异
18 个香椿半同胞家系地径生长与变异见表5。
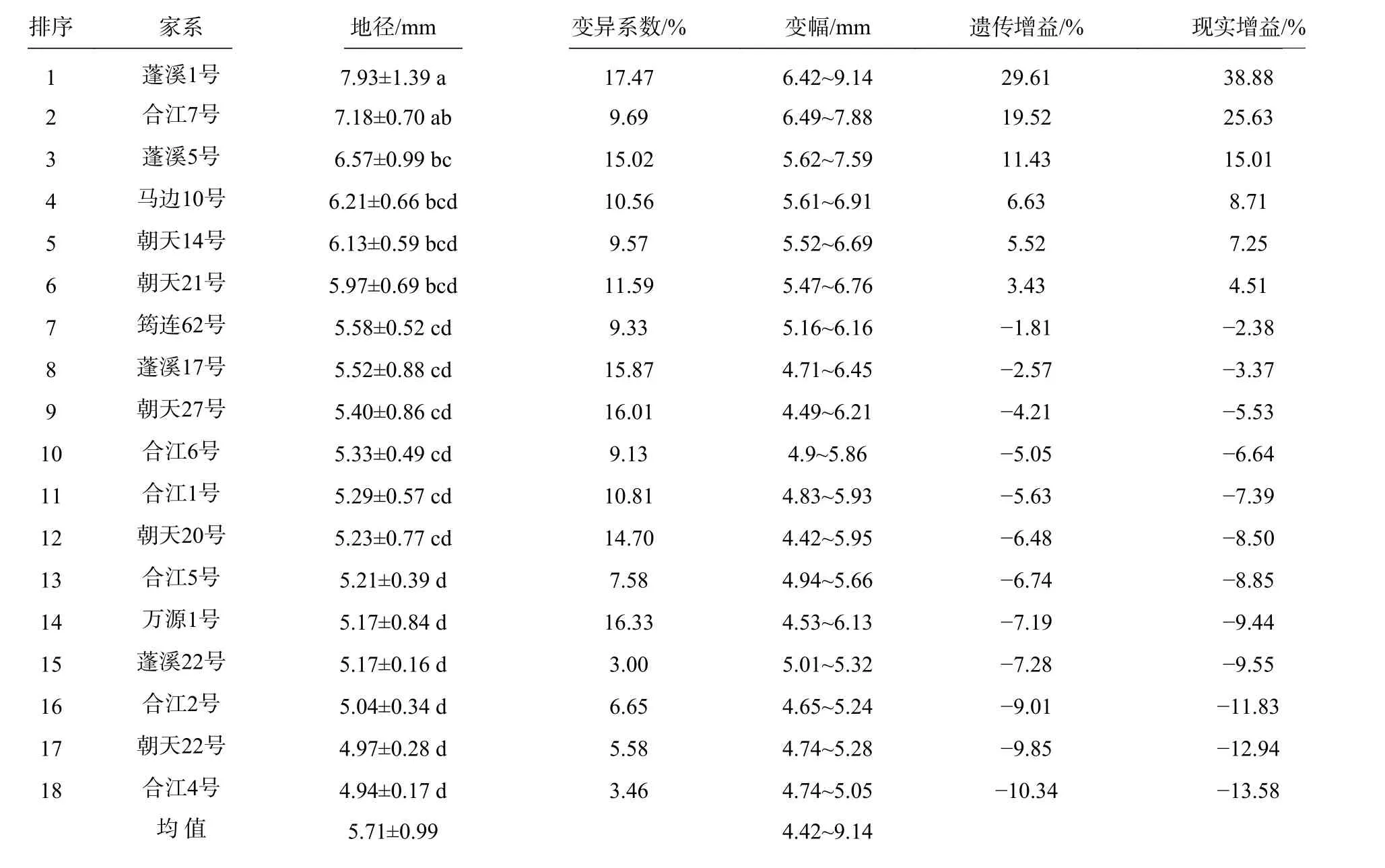
表5 不同家系地径生长变异Tab. 5 Variation of ground diameter growth in different families
由表5 可见,18 个香椿半同胞家系的平均地径为5.71 mm,变幅为4.42~9.14 mm,超过平均值的家系有6 个家系,占家系总数的33.33%。18 个家系的地径与家系所在地理位置没有明显的线性关系。18 个家系中以蓬溪1 号的地径最大,地径为7.93 mm,比平均值高38.88%,变幅为6.42~9.14 mm,变异系数为17.47%,家系内地径有较为丰富的变异,遗传增益和现实增益分别为29.61%和38.88%,其地径与合江7 号没有显著(P<0.05)差异,显著高于其余16 个家系;其次是合江7 号,地径为7.18 mm,比平均值高25.74%,变幅为6.49~7.88 mm,变异系数为9.69%,家系内地径生长较为均匀一致,遗传增益和现实增益分别为19.52%和25.63%,其地径与蓬溪1 号、蓬溪5 号、马边10 号、朝天14 号、朝天21 号5 个家系间没有显著差异,但显著高于地径≤5.58 mm 的12 个家系。18 个家系中,除了蓬溪1 号,还有蓬溪5 号、蓬溪17 号、万源1 号的变异系数大于15%,表明这4 个家系在地径方面存在丰富的遗传变异,其余14 个家系的变异系数均低于15%,家系内地径生长较为均匀一致。
2.3 不同家系苗期比较与选择
由于18 个香椿半同胞家系苗高的遗传力大于地径的遗传力,因此以苗高为主、地径为辅进行苗期家系选择,将18 个家系分为Ⅰ、Ⅱ、Ⅲ、Ⅳ 4 类:遗传增益Ⅰ>Ⅱ>Ⅲ>Ⅳ,分为优、良、中、差4 等,结果见表6。
由表6 可见,Ⅰ类家系共3 个,入选率16.67%,苗高和地径选择差分别为22.80 cm 和1.51 mm,平均苗高和地径分别为83.42 cm 和7.23 mm,遗传增益分别为35.33%和20.19%,苗高和地径增益均较大,为优等家系。Ⅱ类家系只有马边10 号1 个家系,入选率5.56%,平均苗高和地径分别为76.73 cm和6.21 mm,遗传增益分别为24.97%和6.63%,苗高和地径均有一定的增益,为良等家系;Ⅲ类家系包括蓬溪17、22 号、筠连62 号3 个家系,苗高选择差和遗传增益均为正值,但地径的选择差和遗传增益均为负值,苗高生长较好,但地径生长较差,为中等家系;Ⅳ类家系11 个家系,其苗高和地径的选择差均为负值,长势较差,平均苗高和地径均比18 个家系的平均值低,为差等家系。
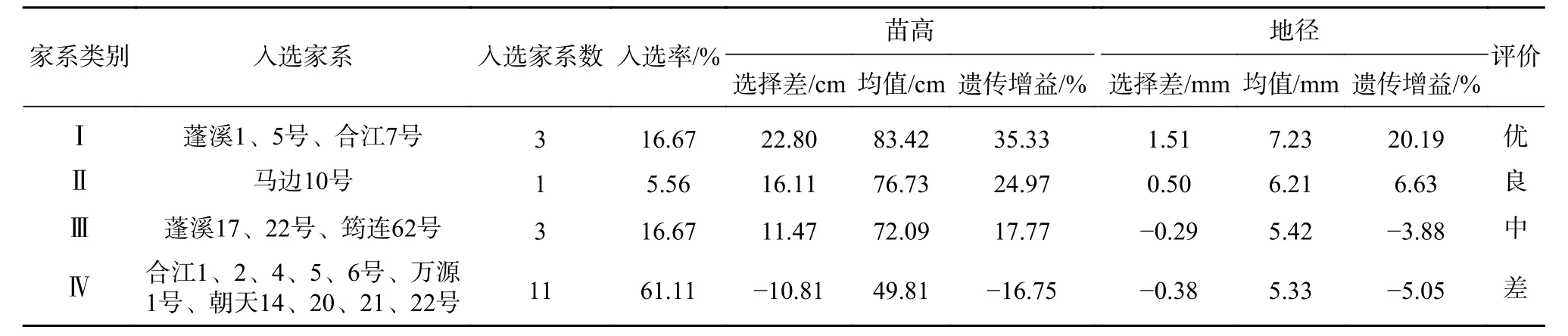
表6 香椿苗期家系选择与遗传增益Tab. 6 Family selection and genetic gain of Toona sinensis at seedling stage
3 结论与讨论
香椿半同胞家系的苗高对原产地的纬度及生态因子表现出非常明显的反应。在原产地纬度上呈现川中、川南高于川北的变异趋势,与梁有旺[17]及刘军[5]等研究结果基本一致;在垂直分布上与海拔有极显著的负相关性,苗高随海拔的增高而减小;在气候因子中与年均气温呈极显著的正相关性,随年均气温的升高而增大,降雨量和无霜期对苗高均没有显著的影响;地径对原产地的经纬度及各环境因子均没有明显的反应。
18 个香椿半同胞家系1 年生轻基质容器苗的平均苗高和地径分别为60.62 cm 和5.71 mm,苗高和地径变幅分别在38.1~101.6 cm 之间和4.42~9.14 mm之间,优劣家系间分别相差1.67 倍和1.07 倍,不同家系间均有极显著差异,与大部分苗期研究结果一致[4-6],具有很好的苗期选择基础。18 个香椿半同胞家系的苗高以蓬溪5 号最大,其次是蓬溪1 号,苗高分别为89.83 cm 和82.67cm,分别比平均值高48.19%和36.37%,变幅分别为80.3~101.6 cm 和68.9~96.7 cm,遗传增益分别为45.26%和34.16%。地径以蓬溪1 号最大,其次是合江7 号,地径分别为7.93 mm 和7.18 mm,分别比平均值高38.88%和25.74%,变幅分别为6.42~9.14 mm 和6.49~7.88 mm,遗传增益分别为29.61%和19.52%。
变异系数的大小决定了苗木生长的均匀程度。18 个半同胞家系的苗高和地径的变异系数总体均小于15%,明显比汉源育苗的14 个家系低[13],家系内生长较为均匀一致,对今后选择优良家系造林有利。蓬溪1 号、合江1 号、朝天27 号3 个家系苗高的变异系数大于15%,蓬溪1 号、蓬溪5 号、蓬溪17 号、万源1 号4 个家系地径的变异系数大于15%,表明这些家系内在苗高或地径中存在丰富的变异,可优先考虑进行超级苗选择与利用,而其余家系的苗高或地径均小于15%,家系内苗木生长均匀一致,对今后选择优良家系造林有利。
遗传力反映亲代将把性状遗传给子代的能力[18],本文18 个家系的苗高和地径的遗传力分别为0.939和0.762,苗高的遗传力比地径大,与表明苗高受到的遗传控制大于地径;苗高和地径有极显著的相关性,因此可以苗高为主、地径为辅进行苗期家系选择。为了提高选择效果,降低入选率,提高选择差是一种很好的方法[19]。从而选择出的蓬溪1、5 号、合江7 号3 个优等家系,入选率16.67%,入选家系苗高和地径的选择差分别为22.80 cm 和1.51 mm,遗传增益分别为35.33%和20.19%,选择差较大,选择率较低,苗高和地径增益均较大,选择效果较好。香椿造林后胸径、树高与苗期的相关性随着年龄的增长而逐渐增大[20],本次选择的3 个香椿优良家系,经造林后有很大成为良种的潜力,对后期香椿良种选育提供了重要的参考价值。
