基于CLIMEX预测黑角负泥虫在中国的潜在地理分布
2020-07-07刘孝贤王振霖吕昭智中国科学院新疆生态与地理研究所中国科学院干旱区生物地理与生物资源重点实验室新疆乌鲁木齐800中国科学院大学资源与环境学院北京0009阿拉山口海关技术中心新疆阿拉山口88新疆农业大学农学院新疆乌鲁木齐800青岛农业大学植物医学学院山东青岛6609
刘孝贤, 李 兰, 王 婷, 张 苹, 张 鑫, 王振霖, 吕昭智,*中国科学院新疆生态与地理研究所/中国科学院干旱区生物地理与生物资源重点实验室,新疆乌鲁木齐800;中国科学院大学资源与环境学院,北京0009;阿拉山口海关技术中心,新疆阿拉山口88;新疆农业大学农学院,新疆 乌鲁木齐800;青岛农业大学植物医学学院,山东青岛6609
黑角负泥虫Oulema melanopusL.又名谷叶甲虫、小麦负泥虫,属鞘翅目Coleoptera叶甲科Chrysomelidae负泥虫属Oulema。主要寄主植物为小麦Triticum aestivumL.、大麦Hordeum vulgareL.、燕麦Avena sativaL.和禾本科 Gramineae杂草(EPPO,2002;Mason&Gillespie,2013)。该虫是国外谷类作物上的一种常见害虫,主要以成虫和幼虫在叶脉间纵向取食叶肉组织进行为害,受害后的叶片出现白色条斑或全叶发白枯焦,严重时甚至会造成植株枯萎,导致谷类作物减产(Hyanes&Gage,1981)。据统计,1995年黑角负泥虫在北美地区暴发,致使春小麦减产55%,冬小麦减产23%。在欧洲,该害虫在谷物幼苗期严重危害旗叶,导致谷物产量下降幅度从波兰的3%~8%到荷兰的95%不等(Mason&Gillespie,2013)。黑角负泥虫成虫一般在白天活动,偶尔飞行,具有群栖性特征,其自然扩散相对缓慢,可随谷物、秸秆及其制成的包装物、草皮、树木和其他可能的载体进行远距离传播,风和流水也能促其扩散(陈乃中,1996)。
入侵物种在入侵地的适生性分析是入侵生物科学风险评估的基础。目前基于生态位理论,应用相关的数学模型来进行适生区决策已经广泛应用于外来入侵物种的扩散、濒危物种的保护以及全球气候变化对物种分布区的影响等研究中(王聪等,2018)。生态位模型是根据物种特殊的生存环境,从目标已知分布区出发,利用数学模型归纳或模拟其生态位需求,然后将其投射到目标地区得到物种适生区分布的模型(蔡静芸等,2014)。潜在地理分布模型和软件主要包括气候相似距模型及数据库系统、地点比较模型与CLIMEX软件、MAXENT模型与软件、GARP模型与软件以及有害生物实验模型与GIS软件等(李志红和秦嘉誉,2018)。
CLIMEX是通过物种在已知地理分布区域的气候参数来预测物种潜在地理分布和相对丰度,分析气候对物种生长发育的影响,能够比较全面评估生物在新的栖息地适生能力的动态模拟模型(Kriticoset al.,2015)。自CLIMEX出现以来,国内外应用其开展了大量的有害生物潜在地理分布预测工作。在国外,澳大利亚、新西兰等国家和地区的学者首先应用CLIMEX地点比较模型进行了有害生物的潜在地理分布进行研究(李志红和秦嘉誉,2018)。例如,对乌桕Triadica sebifera(L.)Roxb.、东方果实蝇Bactrocera dorsalis(Hendel)、茶翅蝽Halyomorpha halysStål等先后进行了潜在地理分布研究(Kriticoset al.,2017; Pattison&Mack,2007; Stephenset al.,2007)。我国于20世纪90年代引入CLIMEX模型,并开展了大量农林业有害生物适生性的研究,并对纳尔塔实蝇Ceratitis rosaKarsch、卷叶毒蛾Epiphyas postvittana、甜菜夜蛾Spodoptera exiguaHübner等多种有害昆虫的潜在地理分布进行了研究(张哲等,2011; Heet al.,2012; Zhenget al.,2012)。此外,CLIMEX还与未来气候模拟数据相结合,特别预测了未来气候变化条件下西印度果实蝇Anastrepha obliquaMacquart、埃及伊蚊Aedes aegypti和马缨丹Lantana camaraL.等多种有害生物的潜在地理分布变化(Fuet al.,2014;Khormi&Kumar, 2014; Taylor&Kumar,2013)。
预测黑角负泥虫在全球范围内的潜在地理分布,对科学防控黑角负泥虫、保护小麦可持续生产及生态安全具有重要意义。本研究利用CLIMEX模型和ArcGIS软件,基于黑角负泥虫的生物学特性,建立黑角负泥虫的生态气候模型,预测黑角负泥虫在全球及我国的潜在地理分布,以期对该虫在我国不同地区的传播风险进行定量评估,同时也为制定谷物生产的防控策略提供科学依据。
1 材料与方法
1.1 研究软件
1.1.1 适生性分析软件CLIMEX模型 CLIMEX是1985年由澳大利亚昆士兰大学Sutherst教授和Maywald博士所研发的商业化软件。本次研究使用的是Hearne software开发的CLIMEX 4.0软件,使用的功能模块为单物种地区比较(Compare Location,1 species)。CLIMEX地点比较模型通过使用物种生长发育的生物学数据进行一系列运算得到生态气候指数(ecoclimatic index,EI)来表示某种特定昆虫在特定地理区域内潜在适宜性(王聪等,2018)。生态气候指数变化范围为0~100,当生态气候指数为0时,表示该物种不适宜长期在该地区生存定殖;生态气候指数越高,表示该物种在该地的气候适宜程度也就越高。
1.1.2 地理信息系统软件ArcGIS 地理信息系统(geographic information system,GIS)是用于存储、管理和显示地理空间数据的计算机系统。主要包括地理空间数据获取、管理、显示、探查和分析等功能(张康聪,2016)。本研究使用的地理信息系统软件是由美国环境系统研究公司开发的ArcGIS 10.2软件,主要使用了其中的插值分析、重分类、制图综合等模块。
1.2 气象数据
本研究使用了CliMond气候数据共享网站以1975年为中心的 30年的空间分辨率为 10′的CSIRO 气候数据集(Kriticoset al.,2012),主要包括月平均降水量,日最高,最低气温,9:00和15:00的空气相对湿度等要素。
1.3 生物学资料
黑角负泥虫一般一年发生一代,卵、幼虫、蛹的发育适宜温度分别为12~32、8~32、8~32℃。 若以7℃为发育起点温度,卵、幼虫(4个龄期)和蛹的发育积温为 105、41、36、43、46 和 282 日度(陈乃中,1996)。越冬成虫最早出现于春天,取食交配,交配过一周后开始产卵,产卵位置多在植株中部叶子的中脉位置(杨静飞,2001)。卵上覆盖有胶状分泌物,暴露于空气中,迅速硬化后固着在叶子上。卵期7~15 d,幼虫逐叶取食,背负分泌物,且只在每次蜕皮后和化蛹前的短时间内光裸。幼虫4龄,经历12~20 d,多发生于4月下旬—6月中旬(刘芳政等,1988)。在4 cm左右深的土室内化蛹,20~25 d羽化出成虫。羽化出的成虫,当年不交配、不繁殖,秋末进入草地、林间、灌丛下和麦田3~5 cm深的土层等处越冬(Websteret al.,1982)。
1.4 气象数据
黑角负泥虫分布数据来源于全球生物多样性信息网 络 (Global Biodiversity Information Facility,GBIF)中发生记录,以及欧洲和地中海保护组织(European and Mediterranean Plant Protection Organization,EPPO)中记载的分布国家和地区。此外,查询一些相关文献对黑角负泥虫在我国新疆、内蒙古等地的分布进行了补充(董淑芳,2016;吉别克等,2012;沙代提力·阿不都等,2018)。
黑角负泥虫主要分布在欧洲的奥地利、比利时、保加利亚、塞浦路斯、捷克、丹麦、芬兰、法国、德国、希腊、匈牙利、爱尔兰、意大利、马耳他、荷兰、挪威、波兰、葡萄牙、罗马尼亚、俄罗斯、塞尔维亚、西班牙、瑞典、瑞士、乌克兰、英国,亚洲的土耳其、以色列、伊朗、土库曼斯坦、乌兹别克斯坦,非洲的阿尔及利亚、摩洛哥、突尼斯,北美洲的美国、加拿大(图1)。
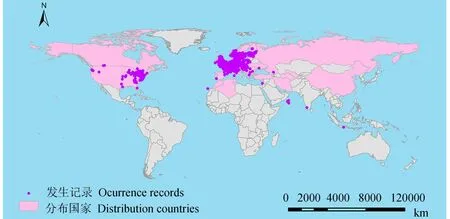
图1 黑角负泥虫在全球的地理分布Fig.1 O.melanopus geographical distribution across the globe
1.5 参数确定
CLIMEX共预设了42个生物学参数,根据黑角负泥虫的实际地理分布和生物学资料,选取其分布集中的西欧和北美作为参照地,反复进行调试,使其分布范围的最大程度与实际相吻合。经过调试,本次研究选用其中的24个参数项,并最终确定黑角负泥虫的预测参数(表1)。
1.5.1 生长指数 生长指数主要包括温度、湿度、光照、滞育等要素,本研究选择与黑角负泥虫分布相关的温度、湿度、滞育3个要素作为生长指数。昆虫的生理活动对温度的变化有直接的响应,温度指数能够反映出这种响应(Shabaniet al.,2012)。根据Olfertet al.(2004)的模型数据与黑角负泥虫的实际分布记录,将 DV0、DV1、DV2、DV3 分别设置为6.5、7、26和35℃。湿度指数指昆虫的寄主植物对土壤湿度变化的响应,它通过影响昆虫寄主植物的地理分布从而影响昆虫的地理分布(Kriticoset al.,2015)。本研究主要参照Olfertet al.(2004)的模拟对北美地区的湿度参数,分别将SM2、SM3设置为1和1.5。为了模拟在中亚荒漠地区黑角负泥虫无法生存的情况,本研究将SM0和SM1提高到0.1和0.15。根据黑角负泥虫的滞育发生情况及其相应的研究,本次研究继续沿用Olfertet al.(2004)的模型参数,分别将 DPD0、DPT0、DPT1、DPD 和DPSW 设置为 14、11、6、120、0。
1.5.2 胁迫指数 冷胁迫中,TTCS的值保持不变,由于CLIMEX模型版本调整的原因,将THCS的值更新为-0.0015。冷胁迫还有一个重要影响因素就是积温,最低积温阈值DTCS和低于此阈值情况下的冷胁迫的积累速率DHCS,分别将其设置为3和-0.0005。热胁迫和湿胁迫的参数因子则继续保持Olfertet al.(2004)的模型参数。在干胁迫中,本次研究将SMDS和HDS更新为0.02和-0.1,湿胁迫的SMWS和HWS分别设置为1.5和0.0005。
1.5.3 有效积温 昆虫的在发育期间需要一定的热量积累,以完成昆虫基本的生长需要(Kriticoset al.,2015)。根据文献中的实验数据,将有效积温PDD的值设置为550。
1.5.4 灌溉参数 灌溉参数值主要影响其在欧洲和亚洲中国新疆地区的分布,GBIF记录其在欧洲地区有大规模的分布,相关文献也曾报道其在中国新疆的巴里坤、裕民等地发生严重危害,因此,灌溉参数值设为冬季0 mm·d-1、夏季 1.0 mm·d-1(Avilaet al.,2019)。

表1 本研究所选取的各生物气候变量信息表Table 1 Environment factors used in this study
1.6 模拟流程
Olfertet al.(2004)对加拿大西部的黑角负泥虫的潜在地理分布及其变化趋势进行了预测,本研究在其参数基础上,根据现有的黑角负泥虫的发生记录(GBIF,2019)对参数进行了补充、更新和完善,使其在全球的适生区模拟与实际分布相吻合;后将调整好的参数输入模型,得到黑角负泥虫在中国不同地区的生态气候指数。将CLIMEX模型模拟出的生态气候指数导入ArcGIS进行插值,绘制黑角负泥虫在中国的潜在地理分布示意图,并通过ArcGIS综合制图模块进行成图。本次生态气候指数的分类以Olfert分类标准为基础,再结合CLIMEX的相关资料,设置如下分类标准:非适生区,EI=0;低度适生区,0<EI≤10;中度适生区,10<EI≤20;高度适生区 a, 20<EI≤30;高度适生区 b, EI>30。
2 结果与分析
2.1 黑角负泥虫潜在地理分布范围
黑角负泥虫的潜在地理分布区域主要集中在北半球亚热带北部和温带地区。欧洲大部分地区均是黑角负泥虫的潜在地理分布,其高度适生区主要分布在法国的巴黎盆地和阿基坦盆地、亚平宁半岛、巴尔干半岛及黑海沿岸地区;中度适生区主要分布在伊比利亚半岛、大不列颠和爱尔兰岛南部、西欧平原、东欧平原大部分地区;低度适生区主要分布在东欧平原北部和东部、斯堪的纳维亚半岛南部和英伦三岛北部地区。非洲黑角负泥虫的潜在适生区面积相对较小,主要集中在摩洛哥、阿尔及利亚北部、突尼斯和南非高原南部地区;此外,在埃塞俄比亚的高原和山脉也有零星分布。在亚洲,黑角负泥虫的高度适生区主要在高加索山脉及里海沿岸一带;中度适生区主要分布在西伯利亚平原南部和东亚的温带地区;低度适生区则主要分布在中亚地区、我国的西部地区、俄罗斯西伯利亚平原和蒙古高原南部。在北美洲,黑角负泥虫的适生区主要集中在美国东部的大西洋沿岸平原和大平原地区,适生程度由东南向西北递减;在落基山脉西侧的狭长地带和墨西哥高原地区也有小面积的分布。南美洲黑角负泥虫的适生区范围相对较小,主要集中在潘帕斯草原及其以南的地区,适生程度由北向南降低;除此之外,西部安第斯山地中也有零星分布。大洋洲的黑角负泥虫适生区主要集中在澳大利亚大陆东南部、新西兰和塔斯马尼亚岛(图2)。

图2 黑角负泥虫在全球的潜在地理分布Fig.2 The potential geographical distribution O.melanopus across the world
黑角负泥虫在我国的适生区分布范围广阔,除海南、香港、澳门、广西、广东、台湾等少数几个省(自治区)之外,其他省级行政区内均有不同程度的潜在适生区域。其中,高度适生区主要集中黄土高原南部、华北平原南部、长江中下游平原及四川盆地周围山地和云贵高原北部地区;中度适生区主要集中在华北平原北部、黄土高原北部、东北平原南部和南方丘陵的北部山区;低度适生区则多分布在中高度适生区的过渡区域、河西走廊、内蒙古西部地区和新疆北部及南部塔里木盆地周围的绿洲地区(图2)。
生长指数描述了某种生物在特定的气候条件下的增长速率,对生态气候指数的变化有着直接的影响(Kriticoset al.,2015)。我国南方地区有着更为良好的水热条件,黑角负泥虫的生长周期长,生长指数高,因此,其生态气候指数也比北方地区高。北方地区则由于温度较低,降水较少,黑角负泥虫生长周期短,生态气候指数也相对较低。我国黑角负泥虫发育较快的时期主要集中在较为凉爽的春季和秋季,而夏季由于气温远远高于黑角负泥虫适宜的温度发育阈值,因此生长指数大幅度降低(图3)。

图3 我国主要小麦产区黑角负泥虫生长指数变化Fig.3 Change of growth index of O.melanopus in major wheat producing areas in China
2.2 不同胁迫因子对黑角负泥虫潜在地理分布的影响
CLIMEX模型中设定限制潜在适生区分布的主要胁迫因子为干胁迫、湿胁迫、热胁迫和冷胁迫(Kriticoset al.,2015)。图4结果显示,对黑角负泥虫在我国潜在地理分布的主要因素是冷胁迫和干胁迫,而热胁迫和湿胁迫则对黑角负泥虫潜在地理分布影响较小。冷胁迫在我国的分布分布范围较广,中度、重度胁迫主要集中在我国的青藏高原、天山及天山以北的新疆地区和内蒙古东部及东北平原的北部地区;黑角负泥虫适生程度较高的东部季风区也受到轻度胁迫的影响;无胁迫的影响区域主要集中在我国南部的岭南地区及海南等。干胁迫主要为中度和轻度胁迫,主要集中在青藏高原南部、新疆天山以南地区、新疆东部、内蒙古和甘肃西部、青海的西北部地区。而热胁迫和湿胁迫则对黑角负泥虫潜在地理分布的胁迫面积小,胁迫的程度低。热胁迫主要分布在新疆吐鲁番盆地及塔克拉玛干沙漠东部,湿胁迫则主要分布在我国岭南地区南部、武夷山区、西藏山南地区和台湾岛。
3 讨论
3.1 黑角负泥虫的潜在地理分布及主要限制因素
黑角负泥虫的潜在地理分布主要集中在25°N-55°N之间的温带亚热带地区,这些地区年均温度普遍在0℃以上,有着良好的降水条件或灌溉条件,是全球小麦等谷类作物的主要种植区域。潜在适生区的南部具有更好的水热条件,适生程度由南向北递减。我国黑角负泥虫的适生区分布趋势与世界分布一致,主要分布在25°N-48°N之间,适生程度由南向北逐渐降低。从气候区划上看,黑角负泥虫的潜在地理分布范围主要集中在我国的东部温带季风气候区和亚热带季风气候的北部地区;从地形上看,黑角负泥虫的潜在分布区在我国南方主要是高原、山地等海拔较高、气候凉爽的地区,在我国北方则主要是平原、盆地等水热条件较好的地区;特别是在降水稀缺的新疆南部地区,黑角负泥虫的适生区紧紧围绕沙漠周围的狭长绿洲带分布。
影响黑角负泥虫在我国潜在地理分布的主要因素为寒冷和干旱,寒冷主要集中在我国的西部和北部地区,干旱影响则集中在我国西部的荒漠地区。根据秦大河等(2005)的预测,未来气候将会向更温暖、更湿润的方向发展,这将使受到干胁迫和冷胁迫地区的胁迫程度降低,生态气候指数增加,更适合黑角负泥虫的定殖、传播。在未来的气候条件下,黑角负泥虫的潜在地理分布会进一步向西和向北地区,同时也会向山脉、高原等海拔更高的地区扩展,如天山山脉、青藏高原等。
3.2 潜在地理分布与实际地理分布的差异
Olfert&Weiss(2006)对黑角负泥虫在北美地区的潜在地理分布进行了预测,但其实验模型仅使用了黑角负泥虫在北美地区的地理分布数据,样本数量较少。本次研究将黑角负泥虫的分布较多的东欧、北欧等地区也列入采样范围;依据其在北欧、蒙古等地的传播,对模型中的温度阈值和冷胁迫参数进行调整,更符合实际分布情况;依据黑角负泥虫寄主的小麦的种植情况,增添了灌溉参数。因此,本次研究对黑角负泥虫的潜在地理分布预测的准确性和参考价值更高。但CLIMEX模型是针对昆虫在气候上的生态适应性来预测其潜在地理分布,实际情况中影响一个物种分布的因素还要考虑自然地理隔离、竞争者、天敌等对物种扩散分布的限制(王聪等,2018);此外,人类对农业生态系统的改造,会改变当地的水热条件,促使黑角负泥虫向低度适生区和非适生区的农业区扩散,这些动态变化因素将会是影响黑角负泥虫分布的重要因素。
我国在20世纪90年代就已有黑角负泥虫分布在新疆地区的记录(陈乃中,1996),但与俄罗斯小麦双尾蚜Diuraphis noxiaM.等许多新疆地区的入侵害虫一样,仅在新疆局部地区传播,并没有传入我国气候适宜度较高的东部小麦主要产区。主要由于自然条件下黑角负泥虫扩散能力较弱(Hyanes&Gage,1981),再加上我国边疆与内地存在着荒漠或高山的阻隔,致使其扩散的范围有限。黑角负泥虫进行远距离的传播主要依靠成虫隐藏在谷物的秸秆、杂草中随人类活动而进行传播(陈乃中,1996),而我国边疆与内地并无相应的大宗商品的贸易流,使其不能凭借贸易等进行远距离的传播。近年来东西部之间贸易流、交通流、货物流的不断增加,致使黑角负泥虫等各种入侵害虫随人为因素的扩散可能性不断增加。因此,相关检疫部门和科研机构应未雨绸缪,建立相应的检疫防控机制,合理规划农业活动,针对黑角负泥虫传播、生物学特性及危害特点采取相应的预防措施。
3.3 全球气候变暖对温带昆虫的影响
全球变暖已经是全世界科学家形成的一个共识,气候变暖将使大多数昆虫物种的地理分布向更高纬度地区扩展(Virkkala&Lehikoinen,2014)。同样的,更加温暖的气候条件使黑角负泥虫逐渐向纬度较高的寒冷地区扩展,很多关于蝴蝶、甲虫、鸟类地理分布的研究也证实了这一点(Lehmannet al.,2015; Parmesanet al.,1999; Virkkala&Lehikoinen,2014)。另一方面,全球出现的暖冬对黑角负泥虫的生活史也将产生重要的影响。滞育是许多温带昆虫度过寒冷冬天的关键机制,它在特定的温度和昼长条件诱发。对于黑角负泥虫这样的温带昆虫,滞育是生活史中的一个必要过程。没有滞育,黑角负泥虫将不能成功越冬。理论研究也表明,滞育在害虫的扩散过程中有着重要的推动作用(Mahdjoub&Menu,2008)。全球变暖导致的温暖冬天将会无法满足黑角负泥虫的滞育条件,因此,未来气候条件下原先适宜程度较高的温带南部和亚热带北部地区适宜程度将会降低甚至不再适宜。
4 结论
黑角负泥虫在我国的潜在地理分布主要集中在25°N-50°N之间的亚热带北部和温带地区,其潜在地理范围面积大、气候适宜程度高。但黑角负泥虫自身扩散能力弱,我国分布的黑角负泥虫和我国小麦主产区又存在着天然的地理隔离,因此对我国小麦生产造成的影响有限。随着近些年西部地区逐渐开发,西部地区与东部地区的贸易流、交通流、货物流不断增多,黑角负泥虫随人为因素传入的可能性不断增加。在全球变暖的气候条件下,黑角负泥虫的潜在适生区未来将进一步向高纬度地区扩展;低纬度地区则由于滞育等因素的影响,其潜在地理分布将会进一步缩减。
致谢:特别感谢中国科学院中亚与生态环境研究中心在黑角负泥虫分布数据与生物学资料等方面,澳大利亚学者Myron教授在模型使用及调整等方面提供的帮助与支持。
