松材线虫galectin-1基因的克隆、原核表达和表达模式分析
2020-07-07任万兰侯有明赵莉蔺福建农林大学植物保护学院闽台作物有害生物生态防控国家重点实验室福建福州5000福建农林大学植物保护学院福建省昆虫生态重点实验室福建福州5000中国科学院动物研究所农业虫鼠害综合治理研究国家重点实验室北京000中国科学院生物互作卓越创新中心北京0009
任万兰, 张 驰, 周 娇, 侯有明, 赵莉蔺,*福建农林大学植物保护学院,闽台作物有害生物生态防控国家重点实验室,福建福州5000;福建农林大学植物保护学院,福建省昆虫生态重点实验室,福建福州5000;中国科学院动物研究所,农业虫鼠害综合治理研究国家重点实验室,北京000;中国科学院生物互作卓越创新中心,北京0009
松材线虫Bursaphelenchus xylophilus(Steiner and Buhrer)Nickle隶属滑刃目Aphelenchida寄生滑刃科(滑刃科)Parasiaphelenchidae(Aphelenchidae)伞滑刃亚科Bursaphelenchiae伞滑刃属(张星耀和骆有庆,2004),是一种世界性重大入侵害虫,在世界范围内引起严重的森林病害,特别是在亚洲和欧洲的国家(Futai,2008; Shinyaet al.,2013;Zhao&Sun,2017)。松材线虫所引起的松材线虫枯萎病,是松属树种最具破坏性的疾病之一,造成全球环境和经济损失总计达数千万美元(Tóth,2011)。
在夏季,松材线虫会经过卵、2龄、3龄、4龄、成虫各个阶段,此为繁殖周期(L1—L4)。到秋末时,松材线虫会遇到低温、缺少食物、种群密度过大等不利环境,这时,松材线虫会由繁殖周期进入扩散周期。松材线虫会先形成扩散型3龄(LⅢ),当遇到枯死在松树里的沟胫天牛族的天牛幼虫时,松材线虫LⅢ就会形成扩散型4龄(LⅣ),LⅣ进入沟胫天牛族天牛的气管内,随着天牛羽化转移到新的寄主上,危害寄主健康。松材线虫在松墨天牛Monochamus alternatusHope气管中被携带,两者间会产生免疫互作(Zhouet al.,2018)。研究表明,携带松材线虫的松墨天牛中,miR-14、miR279和miR-312表达量均显著升高,其功能大多指向代谢、免疫等方面(宁静等,2018)。
半乳凝集素与早期发育和组织再生、恶性肿瘤分期、侵袭性或转移潜能、脂肪生成和Ⅱ型糖尿病、调节先天和适应性免疫稳态有关,以及对感染性和过敏性挑战的反应和癌症方面起关键作用(Blidneret al.,2015; Craiget al.,2010; Fenget al.,2015;Hillet al.,2010; Liuet al.,2012; Pejnovicet al.,2013; Rabinovichet al.,2012; Vastaet al.,2004;Yanget al.,2011)。galectin既参与寄生虫的形态维持、生长发育等过程,还与其寄生、在宿主内存活及免疫调节功能有关(Haoet al.,2007; Kielet al.,2007; Kimet al.,2010; Wanget al.,2007; Younget al.,2004),现已在多种寄生性线虫中发现,如捻转血矛线虫Haemonchus contortusRud.、马来丝虫Brugia malayi(Brug)Buckley、丝状网尾线虫Dictyocaulus filariaRudolphi、哥斯达管圆线虫Angiostrongylus costaricensisMorera and Cespedes、广州管圆线虫Angiostrongylus cantonensisChen。
松材线虫的galectin-1基因在2011年首次被发现(Leeet al.,2011),其结构和功能的研究仍是空白。本实验纯化了galectin-1蛋白,并对其进行了生物信息学分析,并检测了其在松材线虫不同龄期的表达量,为进一步研究松材线虫发育、免疫、与松墨天牛的免疫互作奠定了基础。
1 材料与方法
1.1 供试线虫
松材线虫品系来自中国陕西,并在实验室饲养了数代。采用马铃薯葡萄糖琼脂(PDA)平板培养灰葡萄孢,温度25℃,无光照,7 d左右灰葡萄孢基本长满平板培养基,接入大约500条松材线虫,在同样条件下培养线虫,8 d左右松材线虫将灰葡萄孢吃完,用带有抗生素的磷酸盐吐温缓冲液(PBST)将松材线虫冲出。
各个龄期的获得:收集松材线虫的卵,在水中培养24 h,发育成2龄幼虫(L2);将2龄线虫接种到灰葡萄孢上培养24 h,得到同步化繁殖型3龄松材线虫(L3);2龄线虫接种灰葡萄孢上,培养48 h,得到同步化繁殖型4龄松材线虫(L4);将2龄线虫接种到长满灰葡萄孢的PDA平板上,培养72 h,得到同步化成虫松材线虫。用贝尔曼漏斗法收集各个龄期线虫。扩散性3龄(LⅢ)来自安徽疫区树木,用贝尔曼漏斗法检出。扩散性4龄(LⅣ)来自室内天牛气管中。
将所收集的繁殖型线虫(Ln)、LⅢ和LⅣ线虫离心(5000 r·min-1,5 min),除去多余的 PBST,加入1 mL 35%的蔗糖,6000 r·min-1离心 6 min,将上层的线虫转移到新的离心管中,加入1 mL带有抗生素的 PBST,7000 r·min-1离心7 min,除去 PBST,收集线虫提RNA。
1.2 线虫总RNA的提取,cDNA的合成
根据Trizol试剂盒(Thermo Fisher公司)说明书,提取各个龄期繁殖型线虫(Ln)、LⅢ和LⅣ松材线虫的 RNA,使用 cDNAFast Quant RT Kit(with gDNase)(TIANGEN公司)反转试剂盒合成cDNA,并作为荧光定量PCR的模板。
1.3 galectin-1基因的扩增和测序
根据NCBI上松材线虫galectin-1基因的mRNA完整编码序列(GenBank登录号:GU130138),设计合成特异性引物Galec-F:GATGACTGAGGAAAAGAAAAC和Galec-R:GCTT GGGTTAATGGATCTGGATGCCAG(单下划线为酶切位点EcoR1和Hind3)。使用Phusion High-Fidelity PCR Kit高保真DNA聚合酶(NEB公司)做载体PCR和目的基因片段PCR,反应体系:cDNA模板 1 μL,上、下游引物各 1 μL,dNTP 混合液 0.4 μL,Taq 酶 0.2 μL,Taq 缓冲液4 μL,补充 ddH2O 至20 μL。 PCR 反应条件:95 ℃1 min ;94 ℃30 s,60℃ 30 s,72℃ 30 s,循环 40次;72 ℃ 5 min。 PCR产物经l%琼脂糖凝胶电泳鉴定,送深圳华大基因科技有限公司测序鉴定。
1.4 基因的生物信息学分析
ProtParam(http:∥www.expasy.ch/tools/protparam.html)预测蛋白质的分子量和等电点,Clustal Wprogram(http:∥www.clustal.org/clustal2/)进行蛋白质序列比对,TMHMM分析蛋白质跨膜结构域(http:∥ www.cbs.dtu.dk/services/TMH-MM/),SMART软件(http:∥smart.embl-heidel berg.de/)分析蛋白质功能结构域,PredictProtein(https:∥ppopen.Informatik.tu-muenchen.de/)预测二级结构,SWISS-MODEL软件(https:∥swissmodel.expasy.org/)预测三级结构和同源建模,在NCBI数据库中对松材线虫的galectin进行氨基酸序列的同源性比对,下载线虫的galectin的氨基酸序列,并用MEGA 7(neighbor-joining,NJ法)软件在1000次bootstrap检测后构建进化树。
AB135-S型十万分之一电子天平(瑞士梅特勒-托利多仪器有限公司);LC-20A型高效液相色谱(HPLC)仪(包括DGU-20A5R在线脱气机、LC-20AT输液泵、SIL-20A自动进样器、CTO-20A柱温箱、SPD-M20A紫外检测器及LC-Solution色谱数据处理系统,日本岛津公司);Multiskan MK3型全波长酶标仪(美国Thermo Electron公司);分样筛(上虞市五四仪器厂);TGL-16型高速离心机(金坛市中大仪器厂);GRP-9270型隔水式恒温培养箱(上海森信实验仪器有限公司);HH-S型数显恒温水浴锅(常州翔天实验仪器厂)。
1.5 重组质粒pET-28a-galectin-1的构建
用双酶切法处理目的基因与载体pET-28a,限制性内切酶为EcoR1和Hind3,使用DNA Ligation Kit Ver.2.1(TaKaRa)试剂盒链接目的基因与载体,再将产物热激转化到大肠杆菌DH5α感受态细胞。把细胞均匀涂布在含卡那霉素(100 μg·mL-1)的LB平板上,筛选阳性克隆,阳性菌液送深圳华大基因科技有限公司测序鉴定。
1.6 重组galectin-1蛋白的诱导表达和纯化
接种空载菌株和单克隆pET-28a-galectin-1重组质粒菌株分别于5 mL的LB培养基(含氨苄青霉素 50 μg·mL-1)中,在温度 37 ℃、转速 200 r·min-1的培养箱中培养12 h。加入异丙基-β-D-硫代半乳糖苷(IPTG)至终浓度分别为0、0.1、0.5和1.0 mmol·L-1,分别在 16、37 ℃ 诱导 16 h,5000 r·min-1离心 20 min,沉淀用 100 μL PBS 混匀,超声破碎后,12000 r·min-1离心 10 min,收集上清和沉淀,SDS凝胶电泳后,考染30 min,脱色过夜,检测样品。采用镍柱亲和层析法纯化蛋白。将上清和沉淀的蛋白收集做 western blot。首先将蛋白跑SDS-PAGE电泳,电泳结束后,在转膜buffer浸泡20 min,转到硝酸纤维素膜(PVDF membrane)上,在TBST浸泡10 min,使用快速封闭液封闭10 min,一抗为鼠抗His标签,其浓度为1∶5000,4℃,过夜孵育,使用TBST洗膜5 min,洗3次;再将二抗鼠抗,浓度为1∶10000,加到封闭液中摇床孵育2 h,再用TBST洗5 min,洗3次;用ECL显色液均匀涂抹在PVDF膜上,在黑暗中静止5 min,再用医用X线胶片曝光,置于显色液2 min,水洗,定影液2 min,洗净晾干,扫描数据。
1.7 松材线虫各个龄期galectin-1基因定量表达
使用Peimer premier 5.0设计galectin-1基因的荧光定量PCR引物以及β-actin基因的引物,序列如表1所示:

表1 引物信息Table 1 Primers used in this study
反应体系:SybGreen 10 μL,Rox 0.4 μL,上、下引物各0.8 μL,ddH2O 6 μL,模板2 μL。 反应条件:95 ℃ 3 min;95 ℃ 5 s;56 ℃ 30 s;72 ℃ 30 s;40个循环。以L2作为对照,比较松材线虫其他各个龄期的定量数据,采用2-△△Ct相对定量法计算相对表达量。使用SPASS软件分析数据,分析方法为比较均值单因素方差。
2 结果与分析
2.1 松材线虫的galectin-1基因序列及特点
扩增松材线虫的galectin-1基因并进行测序分析,该序列与 NCBI上所提交的基因序列(GU130138.1)一致,松材线虫galectin-1的 Genbank编号为ACZ13331.1,氨基酸残基数为278,由ProtParam查询可知该蛋白的理论分子质量为31.87 ku,等电点为 6.27。
2.2 松材线虫galectin-1蛋白的功能结构域分析
由SMART软件分析可知,松材线虫galectin-1蛋白的功能结构域有2个,即第10~141位氨基酸处的GLECT结构域(CRD)和第147~276位氨基酸处的GLECT结构域(CRD),由5个氨基酸残基的肽链接(图1)。
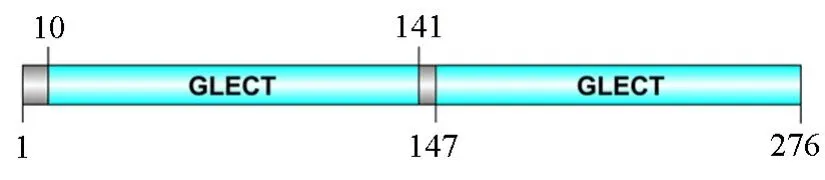
图1 松材线虫galectin-1蛋白结构域的预测Fig.1 Predicted domains in galectin-1 of B.xylophilus
2.3 松材线虫galectin-1蛋白二级和三级结构
由PredictProtein软件分析可知,galectin-1蛋白的二级结构中无规卷曲占 54.35%,β折叠占45.65%,没有α螺旋。使用SWISS-MODEL软件对galectin-1蛋白同源建模,显示出该蛋白主要由无规卷曲和β折叠组成,结果与Predict Protein软件一致。每个半乳凝素中,2个CRD的结构相似,每个CRD具有典型的半乳凝素折叠,该褶皱由反平行的β-折叠构成,β-折叠由 β-三明治组合组成,没有任何 α-螺旋(图 2)。
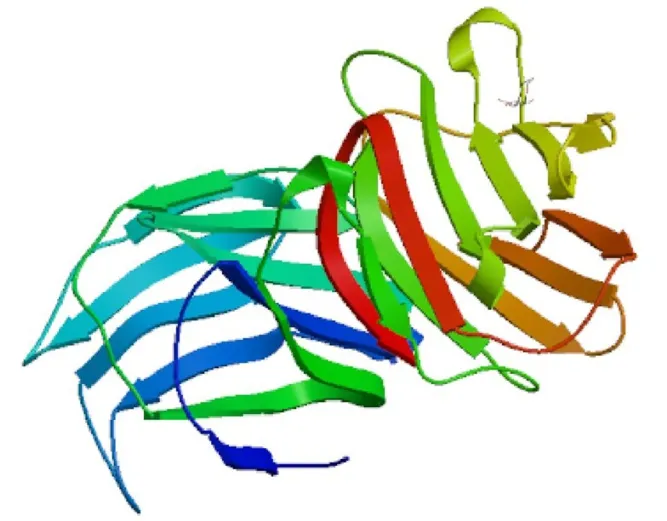
图2 松材线虫galectin-1蛋白的三级结构预测Fig.2 Predicted tertiary structures of galectin-1 of B.xylophilus
2.4 松材线虫galectin-1同源序列对比及系统发育树
通过对松材线虫galectin-1蛋白序列的blast与氨基酸系列分析(图3),发现松材线虫与小卷蛾斯氏线虫Steinernemacarpocapsae(Weiser)(TMS35925.1)的相似性最高,为84.36%;与鼠类圆线虫Strongyloides rattiSandground(XP_024498502.1)相似性为82.01%。以日本血吸虫Schistosoma japonicumKatsurada为外群,用MAGE 7.0建立成系统发育树(图4),线虫都聚集在一个大分支上,而小卷蛾斯氏线虫、鼠类圆线虫和秀丽杆线虫Caenorhabditis elegans在同一个小分支上,与blast的结果一致。

图3 松材线虫与其他物种中galectin-1蛋白氨基酸序列比对Fig.3 Alignment of galectin-1 amino acid sequences of B.xylophilus with other species
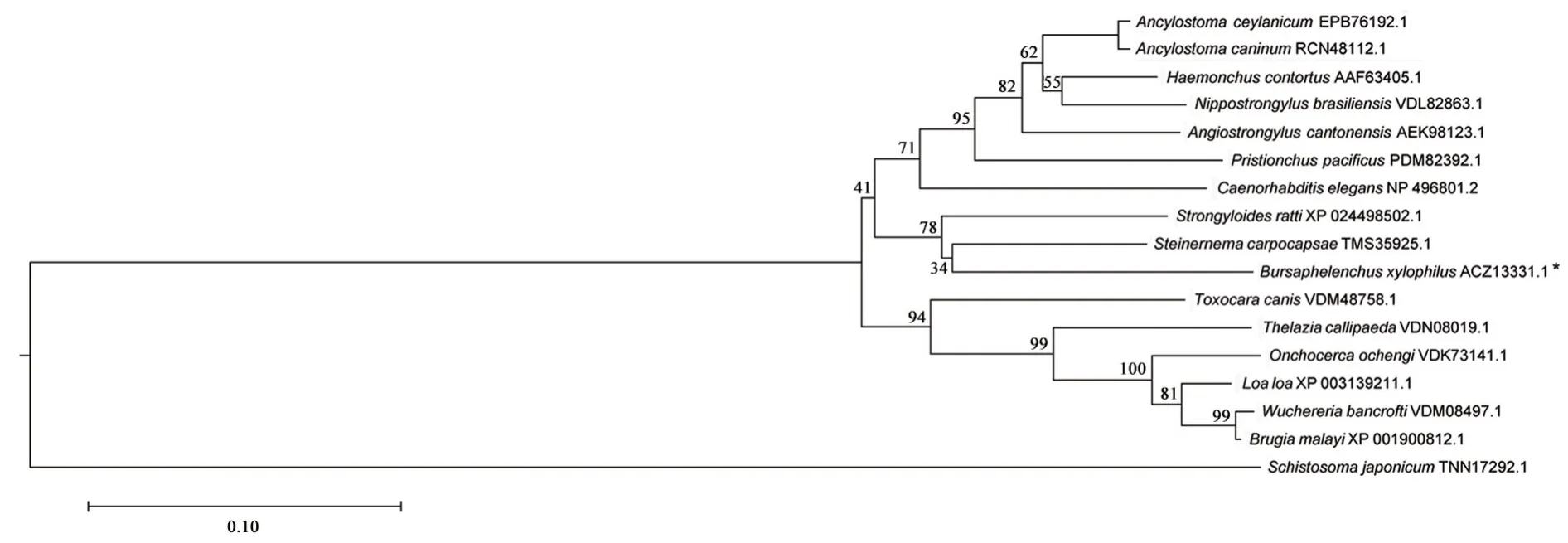
图4 galectin-1与其同源基因基于氨基酸序列构建的进化树(邻接法)Fig.4 Phylogenetic tree of galectin-1 and its homologues based on amino acid sequences(neighbor-joining method)
2.5 松材线虫galectin-1蛋白的原核表达和Western blot
镍柱亲和层析纯化蛋白的Western blot结果与查询所得蛋白分子质量一致。通过构建galectin-1/pET-28a重组表达载体,并转化到DH5α宿主菌中,使用不同浓度的IPTG,分别在37、16℃条件下诱导16 h表达重组蛋白(图5),结果显示,在37℃条件下,表达的galectin-1蛋白较多,沉淀中蛋白的含量比上清多,并且在有IPTG诱导后,表达的galectin-1蛋白会更多。
2.6 各个龄期松材线虫galectin-1基因的定量表达
收集松材线虫各个龄期的cDNA,做galectin-1基因的RT-qPCR,结果表明,相比繁殖型2龄,繁殖型3龄、繁殖型4龄、扩散型3龄和扩散型4龄的galectin-1基因表达量都高,并且呈显著差异(n=3,P>0.05,t检验),与雌、雄成虫没有显著差异;扩散型3龄与繁殖型3龄的表达差异显著(n=3,P>0.05,t检验),扩散性4龄与繁殖型4龄没有显著差异,雌、雄成虫之间没有显著差异(图6);扩散型3龄中galectin-1基因的表达量最高。

图5 重组galectin-1蛋白诱导表达与蛋白纯化Fig.5 Recombinant galectin-1 protein induced expression and protein purification

图6 松材线虫各个龄期的相对表达定量Fig.6 Quantitative expression of relative expression of B.xylophilus at various ages
3 讨论与结论
本文通过原核表达体系,表达和纯化松材线虫galectin-1的蛋白,并预测其蛋白结构,通过与同源基因系统发育树的分析及其在松材线虫不同虫龄的表达量,初步探讨松材线虫galecctin-1基因的可能生理功能。
松材线虫的 galectin-1蛋白有 2个 GLECT(CRD)结构域,这2个结构域可结合碳水化合物,从而发挥生长发育、免疫或其他生理功能。松材线虫是一种植物寄生线虫,其传播需要与松墨天牛相互作用,而通过blast以及氨基酸序列比对,系统发育树结果显示,松材线虫与动物寄生线虫更为接近,特别是昆虫寄生线虫小卷蛾斯氏线虫,说明松材线虫Gal-1的功能可能在与天牛相互作用时发挥作用。通过对松材线虫各个龄期做实时荧光定量PCR,发现 L2、L3、L4和 LⅢ的galectin-1 基因的表达量依次增高,可能与松材线虫的生长发育有关;LⅢ和LⅣ表达量差异显著,LⅢ的galectin-1基因含量最高,可能与天牛蛹室周围的复杂环境有关,因为LⅢ受天牛幼虫所产生的萜烯物质吸引,从而聚集在天牛蛹室周围(Zhaoet al.,2007),而天牛蛹室周围有许多真菌和细菌,环境复杂,线虫从而产生更多的galectin-1提高免疫力。而LⅣgalectin-1基因表达量的减少可能与天牛的免疫逃避相关;雌雄成虫表达量没有差异,说明与性别无关。
从2011年发现松材线虫的galectin-1基因,便再无其他报道。本研究克隆了松材线虫的galectin-1基因,并对其进行了生物信息学分析和各个龄期的RT-qPCR,通过原核表达的方式表达该蛋白,最终获得了纯化后的蛋白,为今后研究松材线虫的galectin-1蛋白与松墨天牛之间的互作提供了参考,为松材线虫的防治提供了新方向。
