不同光温条件谷子光温互作模式研究及SiCCT 基因表达分析
2020-07-02贾小平袁玺垒李剑峰王永芳张小梅全建章董志平
贾小平 袁玺垒 李剑峰 王永芳 张小梅 张 博 全建章董志平,*
1 河南科技大学农学院, 河南洛阳 471023; 2 河北省农林科学院谷子研究所 / 国家谷子改良中心, 河北石家庄 050035
光周期和温度是决定植物开花时间的重要环境因素, 对农作物的生态适应性和产量有重要影响[1]。光周期和温度存在复杂的互作效应, 如拟南芥在长日照(16 h)诱导条件下温度从23℃升高到27℃能够有效促进开花[2]; 大麦在长日照条件下25℃比15℃培养能达到更高级的生殖发育期, 而在短日照条件下结果则相反[3]; 长日照加高温是最不利于开花诱导的光温组合, 长日照条件下, 高温抑制大豆的生殖发育, 低温减弱长日照对发育的抑制作用; 低温处理的大豆能正常出现花序, 而在高温处理下, 大多数植株一直处于营养生长期或部分发生花序的逆转, 表明高温增强了长日照对大豆生殖生长的抑制[4-6]。虽然目前通过对拟南芥、水稻的研究, 光周期调控植物开花的分子机制已经较为清晰[7-8], 有关环境温度调控植物开花分子机制的报道也逐渐增多[9], 但主要集中在光周期或者温度单一因素对植物开花调控作用, 对光温互作分子机制的研究极少。目前的研究初步发现拟南芥CO 蛋白在光温互作调控机制中发挥着重要作用, 短日照条件低温(16 ℃)增加了组蛋白H2A.Z 在成花素FT位点的占领,阻止了光敏色素互作因子4 (PIF4)与FT基因启动子的结合, 高温(27 ℃)则使H2A.Z 与FT位点分离, 诱导CO 蛋白在中午积累, 并与PIF4 形成复合体, 结合FT启动子, 促进基因表达, 诱导开花; 而长日照条件低温使短营养期蛋白(SVP)与FT启动子结合抑制其表达, 温度改变使SVP 在黄昏时活性增加, 同时也增加了夜晚CO 的稳定性, 从而使FT基因表达模式改变[10]。
CO作为光周期调控途径的关键基因, 属于CCT 结构域基因家族, 该家族分为CMF、COL 和PRR 三个亚家族, 其中COL 亚家族含有B-box 锌指结构域和一个CCT 结构域, PRR 亚家族含有PRR 结构域和一个CCT 结构域, 而CMF 亚家族只有一个CCT 结构域[11]。研究表明许多CCT 结构域基因都直接或者间接参与植物开花调控, 是光信号途径中的重要组成部分[12-13]。水稻中首先被克隆的CCT 域基因Hd1是拟南芥中CO的同源基因, 长日照(LD)处理可以推迟水稻抽穗开花, 而短日照(SD)处理促进水稻抽穗开花[14]; LD、SD 条件下,Hd1节律性都在夜间达到一个表达峰值, 该基因还可以通过影响每穗的小穗数(粒数)来控制水稻产量[15]。大麦Hd1的同源基因HvCO1不管日照长短, 其过表达都上调了HvFT1的表达, 使大麦提前开花[16]。高粱CO同源基因SbCO是 COL 亚家族一个成员, 在缺失SbGHD7和SbPRR37的背景下无论长日照还是短日照均能促进植株开花[17]。继Hd1之后在水稻中相继克隆了Ghd7、DTH2、OsCOL10、Ghd7.1(OsPRR37)和Ghd2等含有CCT 结构域的基因, 这些基因多数参与光周期调控的水稻开花过程, 但是具体功能有差异, 在LD 条件下Ghd7、Ghd7.1通过抑制Ehd1和Hd3a的表达延迟水稻抽穗期[18-19], 与Ghd7、Ghd7.1功能类似, 高粱SbGhd7、玉米ZmCCT9在LD 条件下同样能够推迟开花[20-21];ZmCCT基因启动子中的一个CACTA 转座子元件可以抑制ZmCCT的表达, 从而降低光周期敏感性, 使玉米在LD 条件下得以传播种植[22]。DTH2则是通过上调成花素基因Hd3a和RFT1的表达来促进水稻提前开花[23];OsCOL10在LD 和SD 条件下均抑制水稻开花, 还受到Ghd7的调控[24];Ghd2参与对叶片衰老和干旱的调节[25]。从以上研究概述可以发现, CCT 结构域基因普遍参与植物光周期开花调控, 但是这些基因是否参与温度调控的开花过程则极少报道, 特别是在C4作物中, 有关CCT 结构域基因参与光温互作调控开花的研究尚属空白。
谷子(Setaria italica)属于禾本科狗尾草属, 是自花授粉的二倍体(2n=2x=18)作物, 具有抗旱、耐瘠和高光效等优良性状, 基因组较小(约515 Mb), 且全基因组序列已被测定[26-27], 可以作为C4禾谷类理想的模式作物[28-29]。作为一个短日照喜温作物, 谷子对光温较敏感, 前期的研究表明不同光周期环境对谷子株高、叶片数及穗部性状均有显著影响[30], 然而自然环境中除了光周期外, 温度的变化对作物生长发育也有作用, 光周期和温度往往存在复杂的互作效应, 因此通过研究光温互作对谷子等C4作物生长、发育及产量的影响, 为深入揭示以谷子为代表的C4禾谷类作物光温互作调控开花机制奠定基础, 具有重要的实践和理论意义。本研究设置长日照低温、长日照高温、短日照低温、短日照高温4个不同光温处理研究了光温互作对谷子抽穗期、叶片数、穗长、穗码数4 个性状的影响, 明确谷子的光温互作模式, 进一步利用同源克隆技术从谷子中分离获得一个CCT 域基因的完整编码序列, 在对其进行生物信息学分析、组织特异性表达分析的基础上, 详细分析4 个不同光温组合条件下该基因的表达模式, 揭示该基因参与谷子光温调控开花的可能机制, 为进一步深入阐明谷子光温互作调控开花机制奠定基础。
1 材料与方法
1.1 试验材料
选用对光周期较敏感的河北省谷子农家品种‘黄毛谷’。
1.2 谷子材料种植及表型性状调查
采用盆栽方法, 于口径10 cm×10 cm 的小方盆中装营养土, 每盆播8~10 粒种子, 二叶期定苗, 每盆保留4 株长势一致的小苗。8 盆‘黄毛谷’从播种至三叶期均在自然条件下生长, 随后移入光照培养箱进行长日照(16 h 光照/8 h 黑暗)处理, 4 盆在处理1周后采集幼嫩叶片用于总RNA 的提取及SiCCT基因克隆, 另外4 盆继续培养至抽穗用于基因组织特异性表达分析, 处理期间温度为25℃。120 盆‘黄毛谷’从播种至三叶期均在12 h 光照/12 h 黑暗、25℃的条件下培养, 三叶期开始将40 盆‘黄毛谷’转入长日照(15 h 光照/9 h 黑暗)、短日照(9 h 光照/15 h 黑暗)2 个培养箱, 每个处理20 盆, 2 次重复, 每个重复10盆, 2 个光周期处理温度均设定为25 ℃, 用于SiCCT基因长、短日照条件的表达分析; 80 盆移入短日照高温[SD (9 h 光照/15 h 黑暗), 27 ℃]、短日照低温[SD(16 h 光照/8 h 黑暗), 22 ℃]、长日照高温[LD (16 h 光照/8 h 黑暗), 27 ℃]、长日照低温[LD (16 h 光照/8 h黑暗), 22 ℃] 4个培养箱, 每个处理20 盆, 2 次重复,每个重复10 盆, 调查抽穗期、株高、穗长及叶片数,用于光温互作模式分析及SiCCT基因不同光温处理表达分析。
1.3 谷子SiCCT 基因的克隆及生物信息学分析
首先利用生物信息学方法从NCBI 的GenBank数据库中检索到一条谷子CCT 域基因mRNA 序列(XM_004983603), 根据此序列利用软件 Primer Premier 5.0 设计1 对特异引物(SiCCT-F: 5′-TGGG AGGAGGAAGAGAGGAG-3′, SiCCT-R: 5′-CTTCTT CAGGTCCGTGGCTA-3′), 扩增范围包含基因的完整编码区域, 预期大小为1160 bp。用康为世纪的Ultrapure RNA Kit 提取总RNA, 用TaKaRa 公司的PrimeScript II 1st Strand cDNA Synthesis Kit 反转录合成第1 链cDNA, 以合成的cDNA 为模板, 用特异引物扩增目的基因, 扩增体系含cDNA 模板2 μL、2×GC buffer 10 μL、10 mmol L-1dNTPs 0.4 μL、2 μmol L-1特异引物4 μL、TaqDNA 聚合酶0.2 μL,以ddH2O 补充至20 μL。PCR 扩增程序为95℃预变性3 min; 95℃变性30 s, 58℃退火30 s, 72℃延伸45 s, 共35 个循环; 72℃延伸10 min, 4℃保存。将RT-PCR 扩增产物纯化后连接到pBM16A 克隆载体上, 再转化大肠杆菌DH5α 感受态细胞, 挑选阳性菌液送生工生物工程(上海)股份有限公司测序。
利用ProtParam、ExPASy、SOPMA 软件分析SiCCT基因的理化性质、二级和三级结构以及保守结构域。利用DNAMAN5.0 软件将SiCCT基因编码的氨基酸序列与NCBI 下载的其他物种CCT 域基因氨基酸序列进行多序列比对, 使用MEGA 7.0 软件构建谷子与几个物种的系统进化树。
1.4 谷子SiCCT 基因的表达分析
将自然条件下生长至三叶期的‘黄毛谷’幼苗转入长日照条件培养, 取营养生长期的叶片、根、茎节、叶鞘、茎结和生殖生长期的幼穗、穗颈作为试验材料, 每个样品取3 个重复, 用剪刀迅速剪下放入2 mL 离心管, 以液氮速冻, 用于SiCCT基因组织特异性表达分析; 25℃恒温条件下, 长日照、短日照处理的谷苗均从三叶期开始取样, 长日照取到十叶期结束, 短日照取到八叶期开始抽穗, 抽穗后7 d、15 d 取样结束, 期间还对长至五至六叶期的谷苗,从晚6 时黑暗处理开始48 h 内取样, 每隔3 h 取一次, 每个时间点重复取样3 次, 取完全展开的顶叶和第2 叶片, 用于SiCCT基因长、短日照条件表达分析; 4 个不同光温处理的谷苗均从三叶期开始取样直到抽穗, 取完全展开的顶叶和第2 片叶, 取样时间为光照3 h 后, 每个叶龄取1 次样, 3 个重复, 样品以液氮速冻, 用于SiCCT基因4 个光温处理的表达分析。所有样品RNA 提取及反转录过程同实验步骤1.3。
根据SiCCT基因cDNA 序列的编码区来设计实时荧光定量PCR 引物, Q-F 为5′-CGTGGACCCGAC GATGGAC-3′; Q-R 为5′-GTCAGGCACCTTGGCG AATC-3′。参考文献用谷子Actin基因作为内参, 引物为SiActin-F: 5′-GGCAAACAGGGAGAAGATGA-3′和SiActin-R: 5′-GAGGTTGTCGGTAAGGTCACG-3′, 由北京鼎国昌盛生物技术有限责任公司合成[31]。
将反转录好的cDNA 作为Real-time PCR 的模板, 用引物Q-F/Q-R 扩增SiCCT基因的特异保守片段, 同时以引物SiActin-F 和SiActin-R 扩增作内参,每个样品做3 个重复, 扩增体系及循环程序参照宝生物公司TB Green Premix ExTaqII (Tli RNaseH Plus)的试剂说明书, 首先配制10 µL 扩增体系, 包含cDNA 模板1 μL、TB Green Premix ExTaqII (Tli RNaseH Plus) 5 μL、10 μmol L-1的Q-F 和Q-R 特异引物各1 μL、ddH2O 2 μL。使用Roche Light Cycler 96 实时定量PCR 仪, 采用两步法PCR 程序, 扩增程序为95℃预变性30 s; 95℃变性30 s, 60℃退火30 s, 共40 个循环。分析得到扩增曲线、溶解曲线,并计算ΔΔCT 值, 采用2-ΔΔCT计算方法进行相对表达量分析。
2 结果与分析
2.1 谷子光温互作模式
光周期对谷子的生长发育起关键作用, 高温条件下, 长日照处理的‘黄毛谷’比短日照处理抽穗时间推迟了60.3 d, 株高增加了30.4 cm, 叶片数增加了7.7 片, 穗长增加了7.6 cm, 长日照相比短日照营养生长期明显延长; 低温条件下, 长日照处理的‘黄毛谷’抽穗时间比短日照处理推迟了21.4 d, 株高增加7.5 cm, 叶片数几乎一样, 穗长增加了4.7 cm, 说明温度的变化并不能改变长日照比短日照延长‘黄毛谷’营养生长期的总体趋势(图1 和图2-a~d)。同时光周期也决定着温度的作用方向, 短日照条件下,高温处理的‘黄毛谷’与低温处理相比株高减少了11.2 cm, 叶片数减少了2.0 片, 抽穗期提前了11.3 d,穗长减少了1.2 cm, 说明短日照条件下, 提高温度可以使‘黄毛谷’快速完成营养生长, 进入生殖生长期; 长日照条件下, 高温处理的‘黄毛谷’抽穗时间比低温处理延长了30 d, 株高增加了18.8 cm, 叶片数增加了5.6 片, 穗长减少了2.1 cm, 说明长日照条件下, 提高温度使‘黄毛谷’营养生长期延长, 推迟进入生殖生长期, 长、短日照条件下温度的作用方向正好相反(图1, 图2-a~d)。
短日照高温处理的谷子抽穗期最短, 缩短营养生长期的作用最强, 其次是短日照低温处理; 长日照低温、长日照高温2 个处理的谷子抽穗期均比短日照低温、高温2 个处理延长, 且长日照高温组合抽穗期最晚(图2-a)。
2.2 谷子SiCCT 基因的克隆及生物信息学分析
提取长日照处理的‘黄毛谷’叶片总RNA, 经1%非变性琼脂糖凝胶电泳检测看到, 28S rRNA 和18S rRNA 条带清晰, 且前者的亮度大约是后者的1~2 倍(图3)。经过超微量分光光度计检测, OD260/280均在1.97~2.10 之间, 说明总RNA 完整性比较好, 基本没有降解, 可以进行后续实验。将提取的总RNA 反转录得到 cDNA, 以此为模板使用基因特异性引物SiCCT-F、SiCCT-R 进行RT-PCR 扩增, 扩增产物经1%琼脂糖凝胶电泳检测得到一条清晰、明亮的特异性条带, 大小在1000~2000 bp 之间, 与预期目的片段1160 bp 相接近(图4)。

图1 不同光温处理对‘黄毛谷’生长发育的影响Fig. 1 Effect of different photo-thermal treatments on growth and development of ‘Huangmaogu’

图2 不同光温处理间‘黄毛谷’4 个性状的比较Fig. 2 Comparison of four traits among different photo-thermal treatments

图3 ‘黄毛谷’叶片总RNA 电泳图Fig. 3 Electrophoregram of total RNA extracted from leaves of ‘Huangmaogu’

图4 SiCCT 基因RT-PCR 产物电泳图Fig. 4 Electrophoregram of RT-PCR products of SiCCT gene
将RT-PCR 扩增产物经纯化后连接到pBM16A克隆载体上, 再转化大肠杆菌DH5α 感受态细胞,挑选阳性菌液送往生工生物工程(上海)股份有限公司进行测序, 获得大小为1154 bp 的cDNA 序列, 该序列的CDS 全长861 bp, 编码286 个氨基酸(附图1, 附图2)。分析SiCCT 蛋白的结构域发现在215~257 aa之间存在1 个保守的CCT 结构域(附图3), 因此本研究所克隆的SiCCT基因属于CMF 亚家族。
用SiCCT 蛋白序列同源搜索NCBI 数据库, 共检索到9 条与SiCCT 蛋白存在直系同源关系的序列, 这些序列来自水稻(XM_015757733.2)、高粱(XM_002464888.2)、玉米(XM_008660049.2)、节节麦(XP_020151451.1)、二穗短柄草(XM_003574283.3)、海枣树(XP_008796123.1)、油棕(XP_010933637)、野生糜(XP_025825012)、菠萝(XP_020093545) 9 个物种。基于包括SiCCT 蛋白在内的10 个CCT 结构域蛋白序列构建系统进化树, 10 个物种可以聚为3 个组,谷子、高粱、玉米、水稻、节节麦、二穗短柄草聚在一个组, 亲缘关系较近, 它们同属于禾本科; 油棕、海枣树、菠萝聚为一组; 野生糜单独聚为一组(图5)。
2.3 谷子SiCCT 基因的组织特异性表达分析
从图6 可以看出,SiCCT基因在根、茎节、叶、茎结、叶鞘、穗和穗颈7 个不同组织中都有表达, 但是在叶片中的相对表达量最高, 其次是穗和叶鞘,在根部的相对表达量最低。
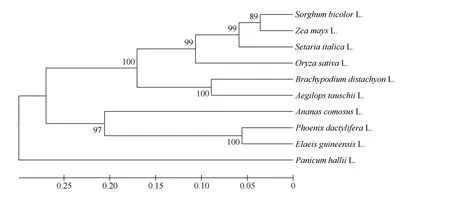
图5 基于CCT 域基因蛋白序列的系统发育树Fig. 5 Phylogenetic tree of CCT-motif genes based on protein sequences

图6 SiCCT 基因在不同组织中的相对表达量Fig. 6 Relative expression of SiCCT in different tissues
2.4 不同光周期条件谷子SiCCT 基因的昼夜表达分析
短日照条件下连续取样48 h, 以研究SiCCT基因在连续2 d 中表达的变化情况, 发现SiCCT基因在‘黄毛谷’叶片中呈现24 h 节律性表达, 黑暗条件0~3 h 几乎不表达, 6 h 之后表达量逐渐增高, 黑暗第12 h 表达达到一个小峰, 随后下降, 在光照开始3 h后达到一个峰值, 然后迅速下降, 在24 h 光照结束后表达量最低; 24 h 之后的下一个循环周期同样SiCCT基因在黑暗6 h (30 h)的时候表达量开始缓慢上升, 在黑暗12 h (36 h)时达到一个峰值, 然后缓慢下降, 见光后开始上升, 当光照3 h (42 h)后再一次达到高峰, 随后又逐渐下降, 在光照快结束时(45 h)达到一个较低的水平。SiCCT基因的表达在1 d 之内有2 个峰, 不过白天表达峰值差不多是晚上峰值的2倍左右(图7-a)。
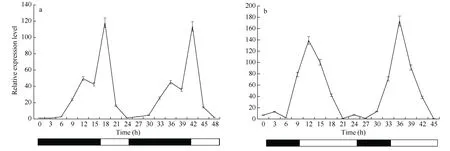
图7 不同光周期条件SiCCT 基因的昼夜表达Fig. 7 Circadian expression of SiCCT gene under different photoperiod conditions
在长日照条件下, ‘黄毛谷’叶片中SiCCT基因的相对表达也呈现出24 h 节律性, 黑暗表达量低, 光照表达量高, 光照开始后表达量迅速增加, 光照3 h后达到一个较高峰值, 随后快速下降, 光照结束后表达降到极低水平; 下一个循环周期与前面 24 h表达模式相似, 同样开始光照后3 h 表达达到峰值(图 7-b)。与在短日照条件不同的是长日照条件SiCCT基因只出现一个表达峰, 且是在光照3 h 后出现, 而短日照条件出现2 个表达峰, 一个是在黑暗12 h, 一个是在光照3 h。
2.5 不同光周期条件、不同生长期谷子SiCCT 基因的表达分析
在短日照条件下, 从三叶期到七叶期‘黄毛谷’叶片中SiCCT基因表达随叶龄的增加而增加, 七叶期达峰值。八叶期谷子开始抽穗,SiCCT基因的相对表达量明显降低, 抽穗后7 d 和抽穗后15 dSiCCT基因的表达也处于较低水平, 说明在谷子营养生长期SiCCT基因表达水平高, 而抽穗后的生殖生长期表达量始终较低(图8-a)。在长日照条件下, ‘黄毛谷’叶片中SiCCT基因从三叶期到十叶期相对表达量均随叶龄增加而增加, 八叶期达峰值, 九叶期、十叶期略下降, 但仍维持较高水平。因为长日照导致营养生长期延长, 十叶期谷子依然处于营养生长期, 没有抽穗, 说明无论短日照还是长日照,SiCCT基因在营养生长期表达量较高(图8-b)。
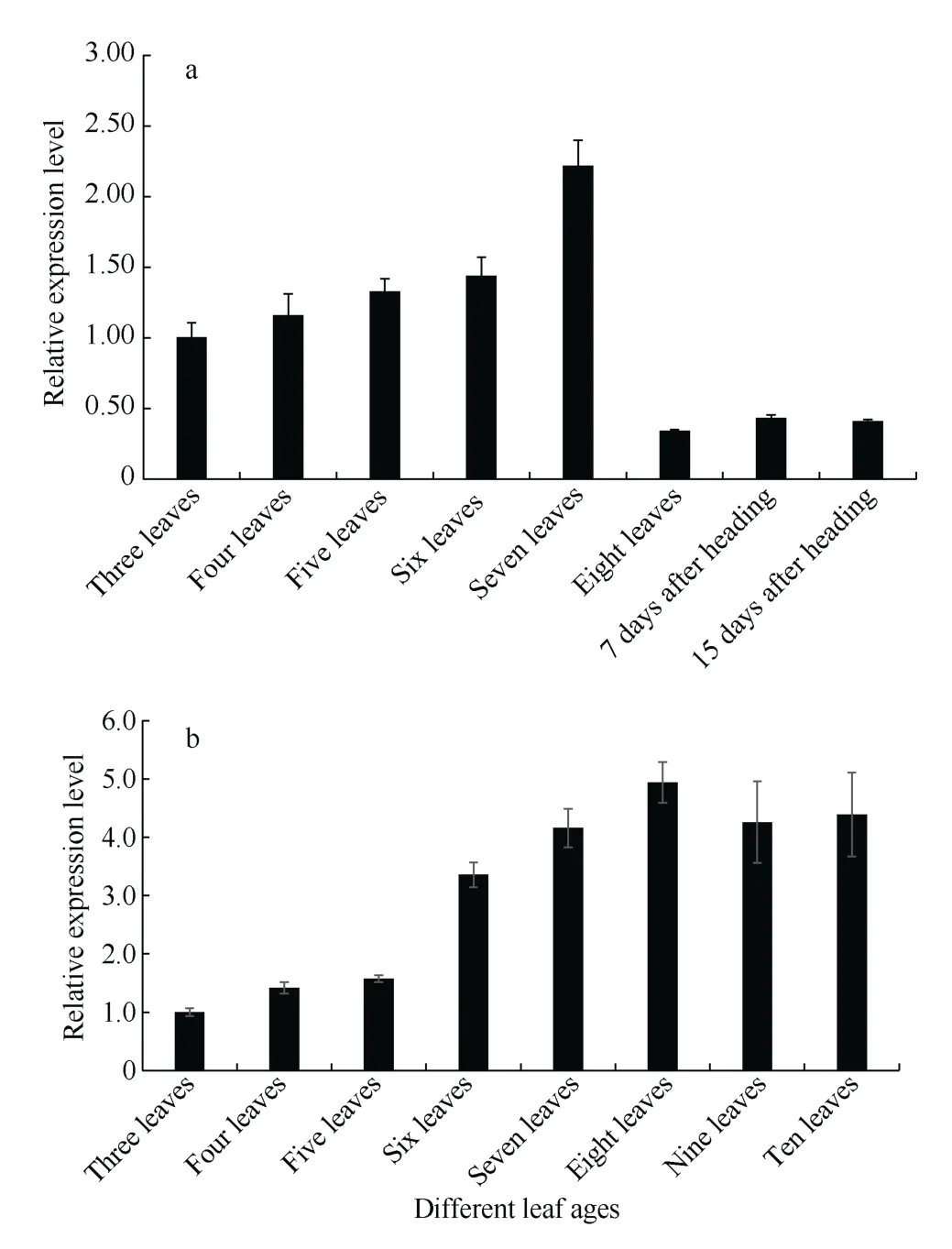
图8 不同光周期条件SiCCT 基因在不同叶龄的表达水平Fig. 8 Expression level of SiCCT gene at different leaf ages under different photoperiod conditions
2.6 不同光温组合条件谷子SiCCT 基因的表达分析
在高温条件下, 三到五叶期长日照处理的‘黄毛谷’叶片中SiCCT基因的表达总体低于短日照处理,从六叶期开始, 长日照处理的SiCCT基因的表达水平明显高于短日照处理(图9-a); 在低温条件下, 三叶期到十叶期短日照处理的‘黄毛谷’叶片中SiCCT基因的表达水平均低于长日照处理(图9-b)。高温短日照处理‘黄毛谷’八叶期已抽穗, 而低温短日照处理‘黄毛谷’十叶期仍处于营养生长阶段, 说明短日照条件下低温对‘黄毛谷’营养生长具有延长作用;在短日照条件下, 三叶期到八叶期营养生长阶段总体上低温处理SiCCT基因的表达水平高于高温处理,只有五叶期高温处理SiCCT基因表达水平高于低温处理, 而八叶期后高温处理的‘黄毛谷’开始抽穗, 进入生殖生长期,SiCCT基因表达水平非常低, 低温处理的‘黄毛谷’仍处于营养生长期, 九叶期、十叶期SiCCT基因的表达水平仍比较高(图9-c); 长日照条件下, 除了三叶期高温处理和低温处理SiCCT基因表达水平接近外, 四叶期到十叶期高温处理‘黄毛谷’叶片中SiCCT基因的表达水平均高于低温处理,与短日照条件表达模式相反(图9-d)。
3 讨论
光周期和温度是影响植物开花的2 个重要环境因子, 而且它们对植物生长发育存在明显的互作效应。本研究发现不论高温还是低温, 长日照条件下的‘黄毛谷’比短日照都晚抽穗, 在短日照条件下, 高温促进‘黄毛谷’抽穗并开花, 低温使‘黄毛谷’持续进行营养生长而延迟抽穗; 长日照条件下, 高温却延迟了‘黄毛谷’的抽穗, 植株长期处于营养生长阶段,而且大多只抽穗很少开花, 结籽很少, 低温能促进‘黄毛谷’进入生殖生长阶段, 提前抽穗并开花。研究结果表明短日照和高温、长日照和低温对谷子的生殖生长都具有促进作用, 而高温加剧了长日照对谷子生殖生长的抑制作用, 低温减轻了长日照对谷子生殖生长的抑制作用, 谷子和大豆具有相似的光温互作模式[5-6]。
目前在水稻、玉米、高粱和甘蓝型油菜中已经克隆的CMF 亚家族成员有Ghd7、NRR、OsCCT01、SbGHD7、ZmCCT9等基因, 这些基因多数受光周期调控, 对作物抽穗期开花期有重要影响[16,22,25,32-33],而谷子CMF 亚家族成员还未有报道, 其与水稻等作物受光周期调控的模式是否相同仍是未知。本研究首次从谷子中分离到一个CMF 亚家族成员, 命名为SiCCT, 组织特异性表达分析表明该基因在叶片中表达量最高, 其次是幼穗, 与水稻Ghd7基因具有相似的组织表达特异性[16], 而多数受光信号调控的基因在叶片中的相对表达量较高, 因此推测谷子SiCCT基因受光周期调控。进一步分析发现SiCCT基因无论短日照还是长日照, 均表现昼夜节律性表达, 短日照条件光照期和黑暗期各有一个表达峰,前者远高于后者; 长日照条件只有光照产生一个表达峰。水稻Ghd7基因无论长日照还是短日照仅在白天有一个表达峰[16], 高粱Ghd7的同源基因SbGHD7在长日照条件下有2 个表达峰, 分别在清晨和夜晚产生[25], 说明谷子SiCCT基因与Ghd7、SbGHD7昼夜表达模式存在差异。然而谷子SiCCT基因与Ghd7、SbGHD7、ZmCCT9也具有相似的表达模式,即无论长日照还是短日照, 整个营养生长期表达量都较高, 短日照条件谷子抽穗及抽穗后SiCCT基因表达量降到极低水平, 而长日照条件一直维持营养生长,SiCCT基因表达量维持在较高水平。因此推测SiCCT基因与Ghd7、SbGHD7、ZmCCT9具有相似的功能, 在长日照条件下高水平的表达抑制抽穗,延长营养生长期。
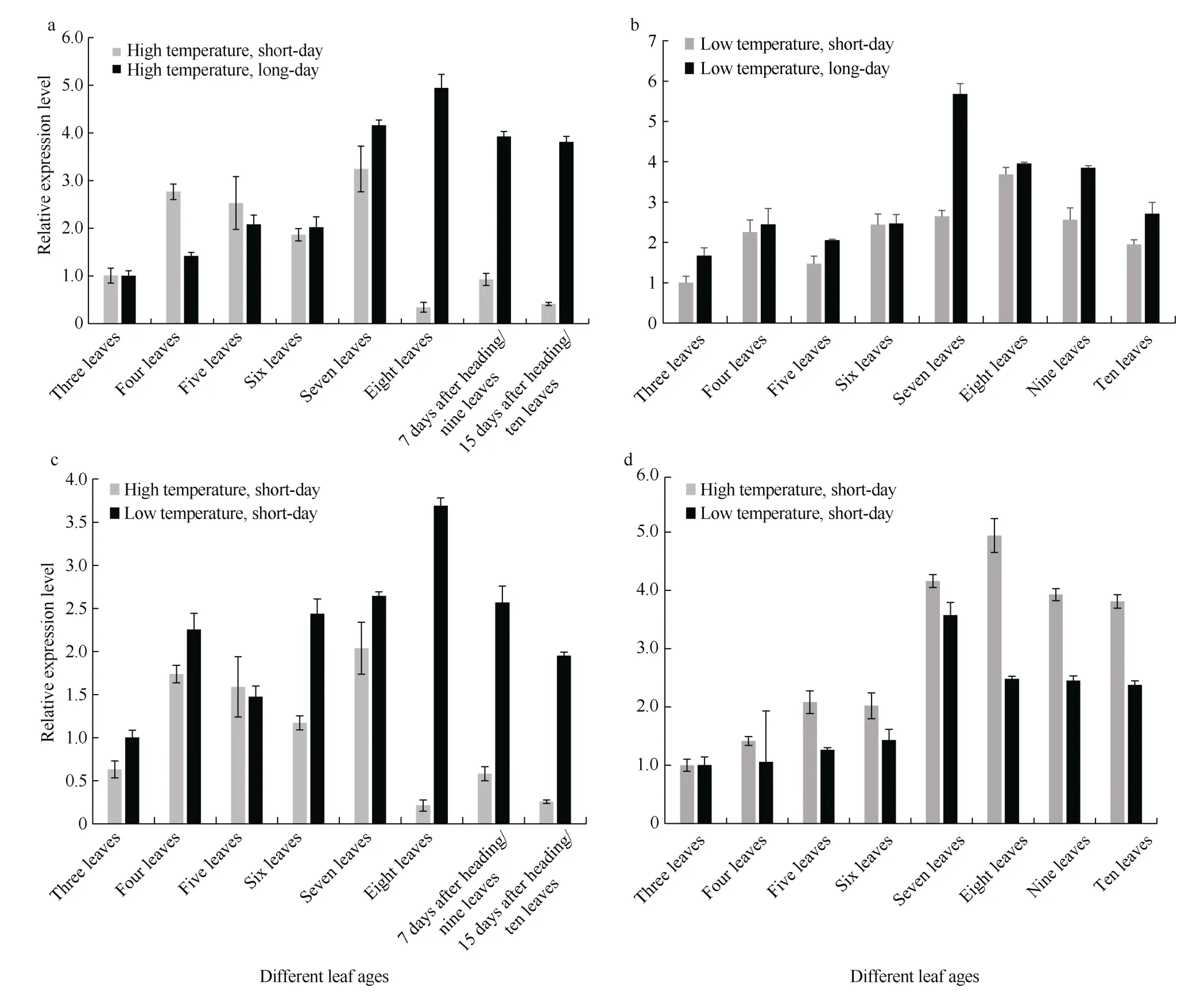
图9 不同光温组合SiCCT 基因的表达特点Fig. 9 Expression feature of SiCCT gene under different photo-thermal combinations
本研究充分说明SiCCT基因和许多CCT域基因一样参与了光周期调控开花途径[34-35], 而光周期和温度对谷子生长发育有明显的互作效应, 因此SiCCT基因是否也受温度调控, 参与光周期途径和感温性途径的交互作用, 是需要弄清楚的下一个问题。通过对拟南芥光温互作机制研究发现,CO基因在光周期途径和感温性途径交互作用中非常关键,短日照条件低温(16 ℃)增加了组蛋白H2A.Z 在成花素FT位点的占领, 阻止了光敏色素互作因子 4(PIF4)与FT基因启动子的结合, 高温(27 ℃)则使H2A.Z 与FT位点分离, 诱导CO 蛋白在中午积累,并与PIF4 形成复合体, 结合FT启动子, 促进基因表达, 诱导开花; 而长日照条件低温使短营养期蛋白(SVP)与FT启动子结合抑制其表达, 温度增加使SVP 在黄昏时活性增加, 同时也增加了夜晚CO 的稳定性, 从而使FT基因表达模式改变[10]。本研究发现SiCCT基因在受光周期调控的同时也受温度调节,长日照条件下, 低温处理的‘黄毛谷’叶片中SiCCT基因的相对表达量明显低于高温处理, 说明在长日照条件下高温促进SiCCT基因的表达, 而低温抑制SiCCT基因的表达, 这与水稻长日照条件下低温(23 ℃)处理促进Ghd7表达的结果相反[36]。而本研究发现高温增加了长日照对‘黄毛谷’生殖发育的抑制作用, 低温减轻了长日照对‘黄毛谷’生殖发育的抑制作用, 这与长日照条件下高温SiCCT基因表达量明显高于低温是相对应的, 更能说明SiCCT基因表达水平与‘黄毛谷’营养生长期存在正相关。无论高温还是低温, 长日照条件下SiCCT基因在不同叶期整体表达水平都高于短日照, 说明SiCCT基因总体上受光周期调控的作用要大于温度。本研究证明了SiCCT基因同时受光周期和温度调控, 而该基因如何与感温性途径的基因相互作用, 并且如何共同作用于谷子成花素基因, 从而实现光温互作对谷子的开花调控, 是后续的主要研究目标。
4 结论
光周期对谷子的发育起关键作用, 不论高温还是低温, 长日照条件下谷子营养生长期要比短日照延长; 温度的作用随光周期的不同而不同, 在短日照条件下, 高温缩短谷子营养生长期而低温促使营养生长期延长, 长日照条件下则相反, 高温延长谷子营养生长期而低温缩短谷子营养生长期; 谷子SiCCT基因在叶片中高表达, 长短日照均表现昼夜节律性表达特点, 整个营养生长期表达量相对较高;SiCCT基因主要受光周期调控, 同时也受温度调控,其表达量与谷子营养生长期具有明显的正相关, 推测SiCCT基因通过参与光周期和温度互作调控机制来调节谷子抽穗期。

附图1 SiCCT 基因cDNA 序列Supplementary Fig. 1 cDNA sequences of SiCCT gene

附图2 SiCCT 基因编码的氨基酸序列Supplementary Fig. 2 The deduced amino acid sequence of SiCCT gene

附图3 SiCCT 基因编码蛋白质的保守结构域预测分析Supplementary Fig. 3 Prediction for the conserved domains of SiCCT protein
