桑天牛幼虫不同部位肠道组织形态学比较
2020-06-05王帆王松华薛苹苹
王帆,王松华,薛苹苹
(上饶师范学院 生命科学学院,江西 上饶 334001)
桑天牛(AprionagermariHope)属鞘翅目天牛科沟胫天牛亚科,在我国大部分地区均有分布[1]。因其幼虫蛀食树干,对桑、无花果、白杨等危害严重,并可为害柳、构树、苹果、枇杷、樱桃、柑橘等树木,是重要害虫。桑天牛成虫将卵多产于1年生的树皮内,孵化后幼虫在枝干内蛀食。由于树木内含有很多木质纤维素等物质,这些物质不易利用,天牛幼虫必须借助消化系统消化获取营养。一般认为,天牛幼虫肠道微生物中的细菌、真菌具有木质纤维素降解功能,如松墨天牛(Monochamusalternatus)中最优势纤维素降解菌为噬纤维细菌科细菌(Siphonobacteraquaeclarae)[2]。而桑天牛成虫肠道中的优势菌群为产酸雷克伯氏菌(Klebsieelaoxytoca),其次为粘滞沙雷氏菌(Serratiamarcescens),其中前者可能具有降低桑天牛肠道pH的作用[3]。桑天牛肠道是消化吸收营养物质的场所,同时也为肠道微生物提供生存环境。但大多数研究人员关注天牛肠道肠道微生物的研究,却很少关注天牛幼虫肠道结构和细胞生物学特点。
昆虫肠道可分为前肠,中肠,后肠三部分[4]。早在1987年,殷幼平就对天牛科消化道进行了比较解剖学研究,并指出其可作为天牛分类的依据[5]。之后周嘉熹和张克斌也对光肩星天牛和黄斑星天牛幼虫消化道外形进行比较,发现黄斑星天牛的消化道短于光肩星天牛,特别是中肠,但黄斑星天牛的中肠膨大部的皱褶较多[6]。李小平等对桑天牛幼虫进行了消化系统组织和形态学的观察,且对桑天牛前肠、中肠结构特点进行描述[7]。但以上研究,均因技术和方法的限制,没有对肠道细胞进行深入研究来探讨天牛幼虫肠道细胞生物学特征与消化吸收难以利用的木质纤维素。另外,王莹曾对9种天牛幼虫的肠道长度和体长做了研究,发现9种天牛幼虫消化道均长于体长,最长的光肩星天牛肠道是体长的2.4倍[8]。因此,对天牛肠道不同部位进行分段细胞生物学比较研究,十分必要。因此,本试验对桑田牛幼虫进行分段切片观察,试图深入解析桑天牛幼虫的肠道结构,为桑天牛害虫生物防治提供理论依据。
1 材料与方法
1.1 主要仪器
体式显微镜,轮式切片机,烘箱,烤片机,显微镜,数码相机,等。
1.2 主要试剂
石蜡,8.5 g/L NaCl溶液,乙醇,二甲苯,蒸馏水,苏木素-伊红染色试剂盒,甘油明胶封片剂,多巴胺。
1.3 实验方法
1.3.1 桑天牛幼虫的捕捉
桑天牛幼虫于2018年12月在上饶师范学院校园内杨树枝干内捕捉,捕捉之后带回实验室进行实验。
1.3.2 肠道组织多酚氧化酶活性染色
对桑天牛幼虫进行低温麻醉,之后在解剖盘上进行解剖。解剖开皮肤以后,用8.5 g/L NaCl溶液冲洗3次,取出完整肠道。放入含有10 mmol/L多巴胺的30%(体积分数)乙醇溶液中浸泡4~5 h,待前肠和后肠黑化之后拍照。
1.3.3 石蜡切片
取出完整的幼虫肠道,将前肠、中肠1(MG1)、中肠2(MG2)、中肠3(MG3)、中肠4(MG4)、后肠1(HG1)与后肠2(HG2)分别用固定液在4 ℃条件下固定12 h。固定液:30%(体积分数)三氯甲烷、60%(体积分数)无水乙醇,10%(体积分数)乙酸。将固定液去除,加无水乙醇进行脱水3次,每次15 min。二甲苯透明2次,每次10 min。最后在液体石蜡中65 ℃包埋12 h。之后进行修蜡,切片,切片厚度5 μm。将组织切片在载玻片上展片,42 ℃烤片12 h。
1.3.4 苏木素-伊红(HE)染色
将带有组织的载玻片在二甲苯溶液中脱蜡2次,每次15 min。之后将玻片依次在无水乙醇、95%(体积分数)乙醇、80%(体积分数)乙醇、70%(体积分数)乙醇浸泡5 min。之后在ddH2O中浸泡2次,每次3 min。将载玻片从ddH2O中取出,吸去载玻片上多余ddH2O,进行苏木素染色8 min,放ddH2O中浸泡10 min,95%乙醇中5 s。进行伊红染色3 min。在70%乙醇中浸泡2次,每次3 min。最后,滴加永久封片剂,盖上盖玻片进行封片,在显微镜下观察拍照。
2 结果与分析
2.1 桑天牛幼虫消化道形态特征
天牛幼虫的肠道长度长于体长,与其它昆虫肠道长度相比差异明显。对桑天牛幼虫进行解剖观察,发现其形态结构如图1A所示,前肠(FG)很短,中肠最长,将其分为MG1、MG2、MG3、MG4 4段。后肠次之,分为HG1,HG2两段。昆虫肠道中有多酚氧化酶表达,用以对食物中的酚类物质进行解毒与诱导粪便黑化[9-10]。肠道组织中的多酚氧化酶原可以被酒精特异性激活,将桑天牛幼虫肠道在30%(体积分数)乙醇、10 mmol/L 多巴胺溶液中染色,发现FG有多酚氧化酶表达可以染黑,后肠HG1前部可以染黑,HG2可以染黑,但HG1后部不能染黑(图1B),说明此部位没有多酚氧化酶表达。由于后肠表达的多酚氧化酶可以杀死肠道内的微生物[10],推测HG1后部有特异的细菌定殖,且这种菌对桑天牛有益。
2.2 桑天牛幼虫消化道形态特征
昆虫前肠具有暂时储存、磨碎食物的功能,其包括口、咽、食道、嗉囊、前胃等部位。对桑天牛幼虫前肠进行切片,其前肠组织从外到内依次为环肌、纵肌、底模、肠壁细胞、内膜等结构(图2A)。但对桑天牛前肠切片进一步观察发现,前肠内膜上有许多微小的齿状结构(图2B),推测和磨碎坚硬的木质食物有关。

A.桑天牛幼虫肠道分段; B.桑天牛幼虫肠道多酚氧化酶检测。

A.前肠横切图; B.A图中方框放大图; C.MG1组织横切图;D.MG2组织横切图; E.MG3组织横切图; F.MG4组织横切图。图2 桑天牛幼虫前肠和中肠组织结构
中肠是昆虫消化吸收营养的主要场所,上接前肠,下与后肠相连。研究表明,桑天牛幼虫的中肠较长,将其分为4段(图1)。MG1段靠近前肠,为膨大部,其HE染色结果如图2C所示。昆虫中肠细胞主要由肠道干细胞、内分泌细胞和柱状细胞组成。对MG1部位肠道结构观察发现,此处中肠有许多皱褶,推测其可增加与肠道内容物接触的表面积,以致快速吸收食物中的有用物质(图2C)。MG2部位有明显的肠绒毛(图2D),且柱状细胞占比较大。MG3部位和MG2部位差异明显,由图中可以看出,内分泌细胞分泌非常旺盛,整个肠腔内充满分泌小泡,这种分泌小泡内的物质可能是消化酶(图2E)。MG4部位的肠道细胞虽与MG3类似,但稍有不同,表现在肠道细胞中内分泌细胞所占比例明显增多,但中肠肠腔内没有大量分泌物质,推测两部位分泌物质不同,且MG4部位的柱状细胞没有MG3部位典型(图2F)。
后肠是排泄器官,后肠的功能是回收水分和无机盐。通过观察,桑天牛幼虫后肠很长,但HG2比HG1短,HG1的结构如图3A所示,与内膜连接的是一层上皮细胞(图3B);HG2结构如图3C所示,其内膜比HG1要厚,且6个直肠垫结构非常明显;桑天牛幼虫后肠HG2肠道内有隐肾管,有助于水分和无机盐的重吸收。
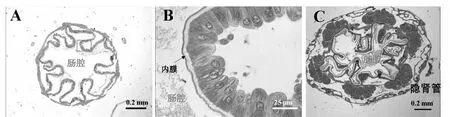
A.后肠前部组织(HG1)横切图; B.图A中红色方框放大图; C.后肠后部组织(HG2)横切图。
3 结论
昆虫肠道是消化、吸收和排泄的器官,是昆虫重要的组织结构。其前肠和后肠由外胚层发育而来,二者在结构上类似,而中肠起源于内胚层。由于不同种类的昆虫生存环境和食性等不同,各类昆虫的消化道有很大变异。果蝇的中肠可分为前中肠(anterior midgut,AM)、铜细胞区域(copper cell region,CCR)和中后肠(posterior midgut,PM)等,其不同部位存在的干细胞也有差别,如CCR区的干细胞为胃肠干细胞,PM区的干细胞是肠道干细胞[11]。天牛幼虫肠道大于体长,与其它昆虫相差较大,而具体某一区段的肠道结构研究还鲜有报道。本文对桑天牛幼虫不同部位的肠道进行解剖、切片、染色、观察,发现桑天牛幼虫前肠和后肠有多酚氧化酶表达来帮助其分解植物酚类物质。根据桑天牛中肠形态和在体腔内分布状况,可分为4个部分。此4个部分的中肠肠道结构有明显差别,如MG1肠道褶皱较多,MG2肠道细胞的肠绒毛丰富,MG3肠腔内充满大量分泌小泡,MG4的内分泌细胞较多。桑天牛幼虫4个部分的中肠肠道结构特征可能与消化木质素有关。Scully等对星天牛转录组幼虫中肠进行转录组测序发现,180个单基因编码糖苷水解酶类(GHs)。包括几种GH5、45和48纤维素酶,GH1木聚糖酶,GH1 b-葡糖苷酶;并且发现有一些植物宿主防御化学物质解毒酶[12]。然而Scully等将整个中肠进行实验,并没有分段取样,鉴于中肠结构的差异,有必要对其蛋白质组或转录组进行分段深入研究。本研究对蛀干类害虫天牛幼虫的肠道结构研究有一定帮助。
