滨海盐生植物叶片δ13C与主要养分元素化学计量的关系
2020-06-03夏大娟刘秋蓉LiliannZou葛之葳薛建辉彭思利
夏大娟,刘秋蓉, Liliann Zou,葛之葳,*,薛建辉,3,彭思利
1 南方现代林业协同创新中心, 南京林业大学生物与环境学院, 南京 210037 2 Saint John′s School, San Juan 00907, USA 3 江苏省中国科学院植物研究所, 南京 210014
滨海湿地高盐环境与土壤贫瘠限制植被生长,盐生植物遗传多样性对环境产生特殊适应性[1- 5]。依据盐生植物耐盐特性,将盐生植物分为稀盐盐生植物(salt-dilution halophyte)、拒盐盐生植物(salt-exclusion halophyte)、泌盐盐生植物(salt-secretion halophyte)[5]。稀盐盐生植物叶片或茎肉质化发育,增加水分储存,缓解生理干旱[1, 5];拒盐盐生植物,对K+选择性吸收,或将Na+贮藏于根等特殊部位,保持体内K+/Na+平衡,以维持体内渗透压,缓解Na盐毒害[3, 5-6];泌盐盐生植物特殊泌盐结构能将盐分排除出体外[1, 5],这些特征是研究植物遗传适生性的基础。
植物碳稳定性同位素δ13C反映细胞间与环境CO2浓度比(Ci/Ca),由叶片光合能力(A)与气孔导度(gs)之间平衡决定[7- 8]。δ13C因与水分利用效率(Water use efficiency: WUE)正相关成为研究植物长期WUE的有效替代[9]。滨海湿地高盐环境与养分胁迫严重阻碍植被生长[10-11],多数学者关注盐分因子,认为盐生环境植物δ13C与土壤盐度[12-13]或自身耐盐性有关[13]。盐生环境养分胁迫继渗透胁迫和离子毒害后成为限制植物生长的因素[14- 16],但不同盐生植物叶片养分元素与植物碳同位素组成(δ13C)的关系及植物养分利用策略与WUE之间的权衡模式仍然未知。
叶片N、P、K养分元素对植物生长、发育有着重要作用,通过影响植物生理过程,如氨基酸、蛋白质、核酸等物质合成(N)[17]、卡尔文循环酶活性与合成蛋白相关酶活性(P)[17-18]、细胞渗透压调节与Na盐毒害缓解(K)[2-3, 19],直接或间接影响δ13C[18, 20- 22]。叶片C/N、C/P表示植物固定单位C消耗的N和P量,反映了植物的N利用效率(Nitrogen use efficiency: NUE)和P利用效率(Phosphorus use efficiency: PUE)[23]。N/P常用来表征植物养分亏缺状况,植物N/P<14,表明植物生长受N限制;植物N/P>16,表明植物生长受P限制[11, 24-25]。植物C/N、C/P、N/P与蒸腾相关,δ13C与WUE正相关成为研究碳-水-养分循环的有力工具[21-22, 25]。
叶片δ13C与养分元素化学计量特征的关系,因研究区域环境差异与物种差异而不同[18, 20- 22, 26]。众多研究表明,叶片N含量与叶片δ13C正相关[18, 22, 26];叶片P通过影响光合相关酶活性与δ13C正相关[26],或通过质流效应与δ13C负相关[21-22]。K+在渗透压调节中起重要作用,被证明与δ13C正相关[19],而部分研究也表明K+还可以通过气孔导度调节与δ13C负相关[22]。植物在低N环境通过高效水分利用代偿减少的N利用率,揭示δ13C(WUE)与NUE负相关[20, 22-23]。Dijkstras等[23]认为由于微生物对土壤P具有很强的固定作用,导致WUE与PUE之间没有相关关系。Zhou等[21-22]研究得出,由于质流效应,P吸收受蒸腾压控制,WUE与PUE正相关,即δ13C与PUE正相关。Cernusak等[25]研究热带树种叶片N/P与WUE关系,发现WUE与N/P正相关。而在滨海特殊生境(高盐环境、土壤贫瘠)下WUE与NUE之间的关系尚未见直接报道,这对于解释不同生理生态策略的盐生植物应对环境压力的生理机制具有重要意义。
目前δ13C与叶片养分元素化学计量特征之间的关系被成功应用于干旱和半干旱区研究[18-19, 22, 26-27],关于滨海湿地研究较少。本研究在对江苏盐城滨海湿地草本植物群落进行调查的基础上,以常见的稀盐盐生、拒盐盐生植物为研究对象,分析两种耐盐型植物叶片δ13C与养分元素化学计量特征的关系,揭示不同耐盐型植物遗传适生性,为滨海湿地生态系统恢复、重建与保护提供理论基础。
1 材料与方法
1.1 研究区概况
研究区位于江苏省盐城滨海淤泥质海岸带湿地(32°52′16.30″—33°38′21.62″N,120°34′33.85″—120°55′1.44″E),濒临黄海,为射阳县、大丰区、东台市近海滩涂湿地。该地位于北亚热带和暖温带交界处,气候具有明显过渡性,属北亚热带季风气候,雨热同季、雨量集中在6—8月,四季分明,年均气温13.7—14.4℃,冬季较冷,夏季较热,无霜期为209—218 d,年降水量1025 mm,年均日照时数2169.6 h,白天日照充足。土壤质地为砂质壤土,pH在7.89—8.50之间[28-29]。植被类型为滩涂湿地植被,主要优势科为禾本科(Poaceae)、藜科(Chenopodiaceae)、菊科(Compositae),原生植物包括盐地碱蓬(Suaedasalsa)、碱蓬(Suaedaglauca)、盐角草(Salicorniaeurpaea)、芦苇(Phragmitesaustralis)等[30]。
1.2 样品的采集
1.2.1样品采集与预处理
样品采集:沿海岸线自北向南设置间隔大于18 km的3个样点(图1),运用样线结合样方调查法[30],在植物生长季(2018年6月),对滩涂湿地草本植物进行实地踏查。每个样点调查10个1 m×1 m样方,每个样方间隔20 m以上,共30个样方,记录群落特征。依据赵可夫[5]盐生植物分类系统对调查结果进行分类汇总(表1)。
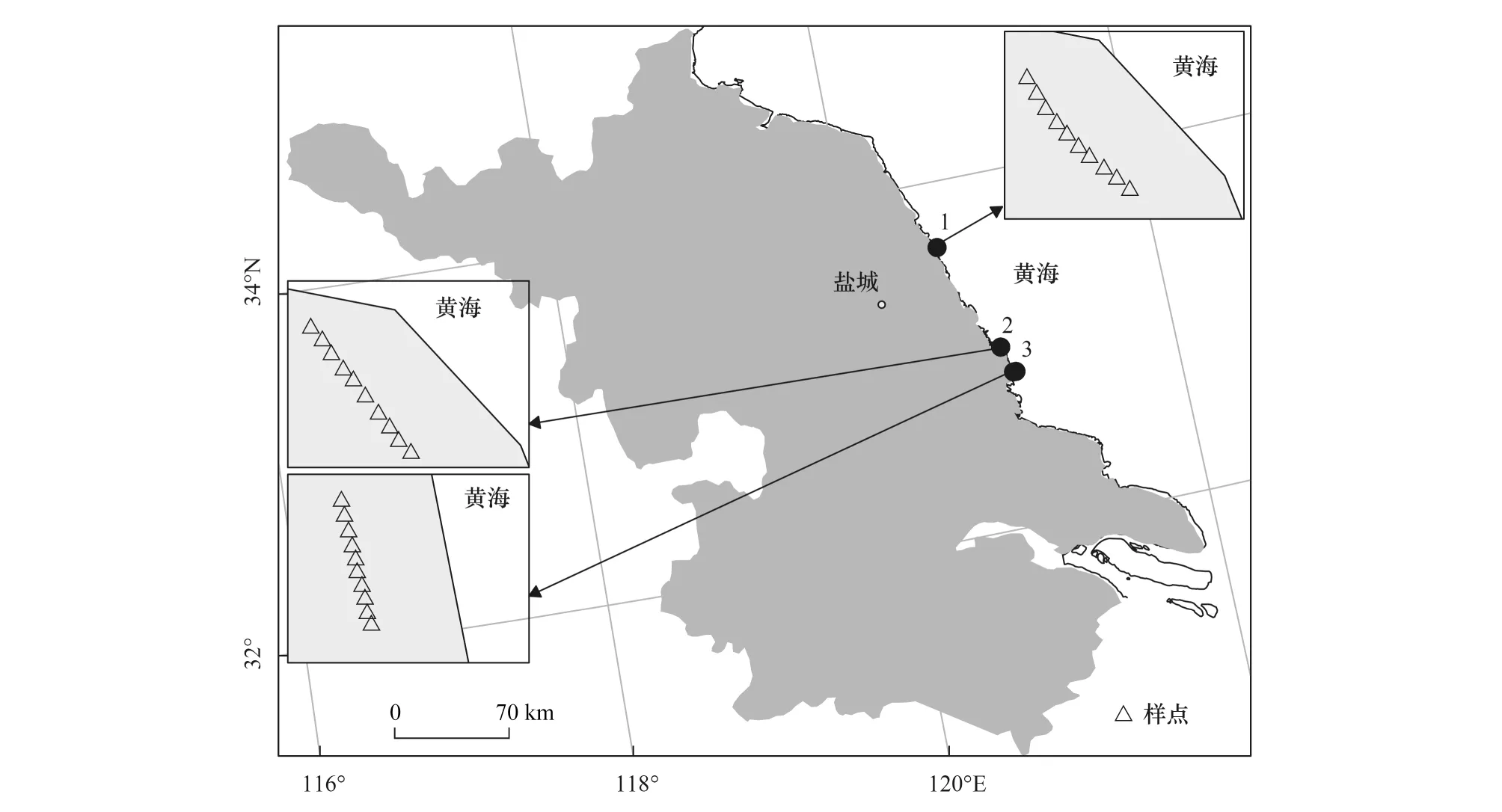
图1 研究区与采样点Fig.1 Study area and sampling site
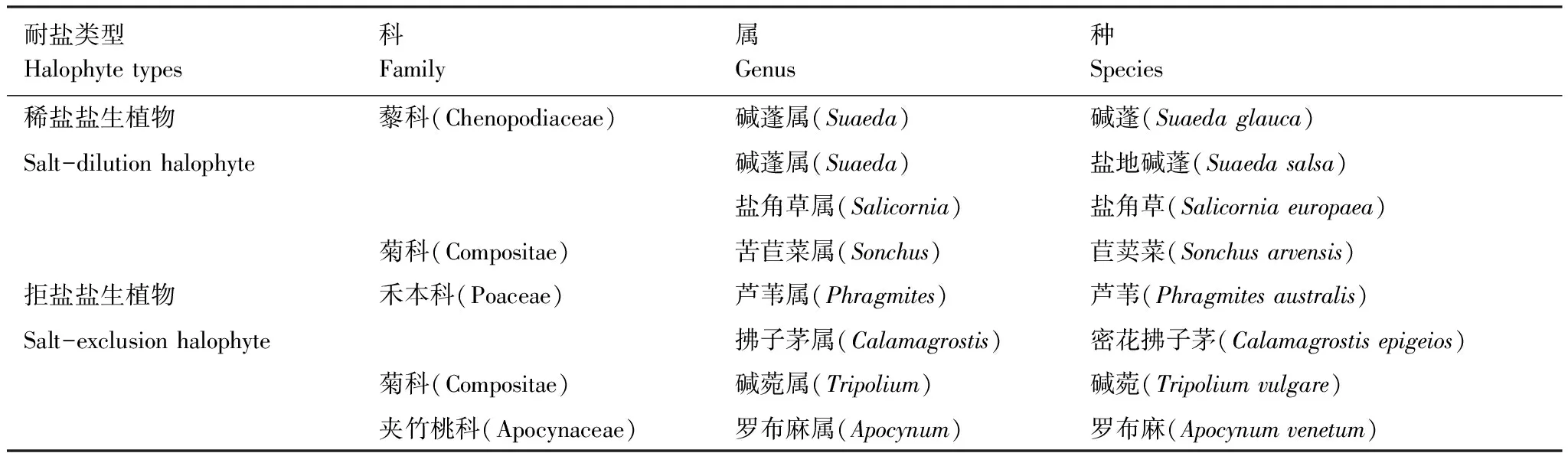
表1 盐生植物样本列表
预处理:对样方植物进行采集,整株带回,分离出根、茎、叶,用去离子水洗净各部分,105℃杀青30 min,并于55—65℃烘干至恒重,用药用粉碎机或全自动粉碎仪研磨,过100目筛,待测。根际土壤样品剔除枯枝落叶及碎渣,于阴暗处风干15 d,采用玛瑙研钵进行研磨,过100目筛,装袋封存,待测。
1.2.2指标测定
元素测定:叶片全碳、全氮浓度(%)采用元素分析仪(Elemental Analyzer, Perkin-Elmer 2400 II,USA)测定;叶片全磷、全钾含量采用王水-微波辅助消解-ICP-AES法测定。
稳定碳同位素自然丰度测定:采用稳定同位素比例质谱仪(DELTA V Advantage,USA)测定,叶片碳稳定性同位素组成(δ13C,‰)表达式如下:
δ13C(‰)=[(Rsample-Rstandard)/Rstandard]×1000
(1)
式中,Rsample、Rstandard分别为叶片、标准样品中13C/12C; Rstandard采用国际标准物质VPDB。
δ13Cp=δ13Ca-a-(b-a)Ci/Ca
(2)
式中,Ca:大气中CO2浓度,根据网站http://www.esrl.noaa.gov查阅可得;δ13CP:植物组织碳同位素比率,可通过(1)式计算而得;δ13Ca:大气CO2碳稳定性同位素比率,计算方法依据Feng[31]提供的计算式:
δ13Ca= -6.429-0.0060exp[0.0217(t-1740)]
(3)
式中,t为研究年份2018,δ13Ca= -8.93‰;a、b: CO2扩散、羧化反应产生的分馏效应(4.4%、27%)。
WUE=A/gs=(Ci-Ca)/1.6=Ca/1.6x(δ13Cp-δ13Ca+b)/(b-a)
(4)
式中,A: 光合速率;gs: 气孔导度;Ca、δ13Cp、δ13Ca、a、b含义同上。
1.3 数据处理与分析
研究采用Microsoft Excel 2016 软件对数据进行整理、SPSS 22.0统计分析软件进行统计分析(ver.22.0; SPSS Inc.,美国)、利用Origin 8.5制图。线性回归分析应用于研究物种δ13C与养分元素化学计量特征关系分析,相关性分析采用Pearson相关,显著性水平P<0.05。
2 结果与分析
2.1 滨海湿地盐生植物叶片δ13C与元素含量化学计量学特征
稳定性同位素自然丰度(δ13C)在不同植物间存在显著差异(P<0.05)(表2),不同耐盐植物遗传适生性不同,叶片对养分富集程度不同(表2)。拒盐生植物叶片δ13C、C含量均显著大于稀盐盐生植物(P<0.05),叶片N含量((30.8±9.6) g/kg)大于稀盐盐生植物((23.4±10.2) g/kg),但未达到显著性水平(P>0.05)。稀盐盐生植物叶片P含量比拒盐盐生植物高0.4 g/kg,但差异性不显著(P>0.05)。拒盐盐生植物叶片K+含量较稀盐盐生植物高2.6 g/kg,但未达到显著水平(P>0.05)。 C/N、C/P、N/P化学计量学统计结果见表2,结果表明,叶片C/N、C/P元素比在两种盐生植物之间差异性不明显,但N/P在两种盐生植物之间存在显著性差异(P<0.05),稀盐盐生植物N/P<14,拒盐盐生植物N/P>16。
滨海湿地盐生植物叶片元素含量及化学计量学特征与其他研究区差异比较见表3,盐生植物C含量较其他区域都低,但N、P较其他区域高。因叶片C含量较小,C/N与C/P表现出比其他区域小的特征。研究区N/P平均值为15.3,大于全球范围、中国东部南北带森林生态系统102个优势种植物及中国东黄海海岛植物,但小于荒漠地区植物。
2.2 盐生植物叶片δ13C与元素N、P、K关系
叶片养分元素通过影响植物生理过程与δ13C相关(图2)。无论是盐生植物总体样本结果,还是两种不同耐盐类别的盐生植物,δ13C与N均呈显著正相关性(P<0.05);但由于不同盐生植物δ13C丰度分布差异明显(表2),其与P的相关性关系只在两种耐盐类型中被发现,两种植物δ13C与P均呈显著正相关性(P<0.05),而盐生植物总样本相关性不明显。除稀盐盐生植物外,拒盐盐生植物、总样本δ13C与K+浓度显著正相关(P<0.05)。
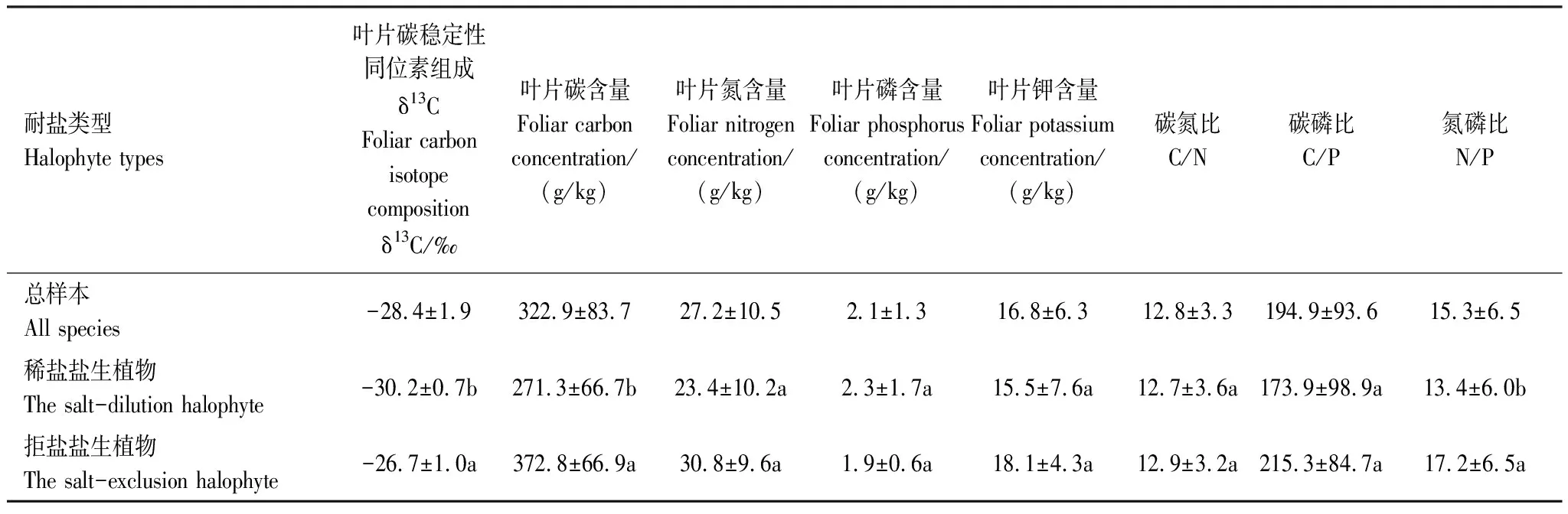
表2 不同盐生类型植物叶片δ13C与元素含量及化学计量特征(平均值±标准偏差)
同列不同小写字母a、b表示数据间差异达到显著水平(P< 0.05),稀盐盐生植物n=29,拒盐盐生植物n=30
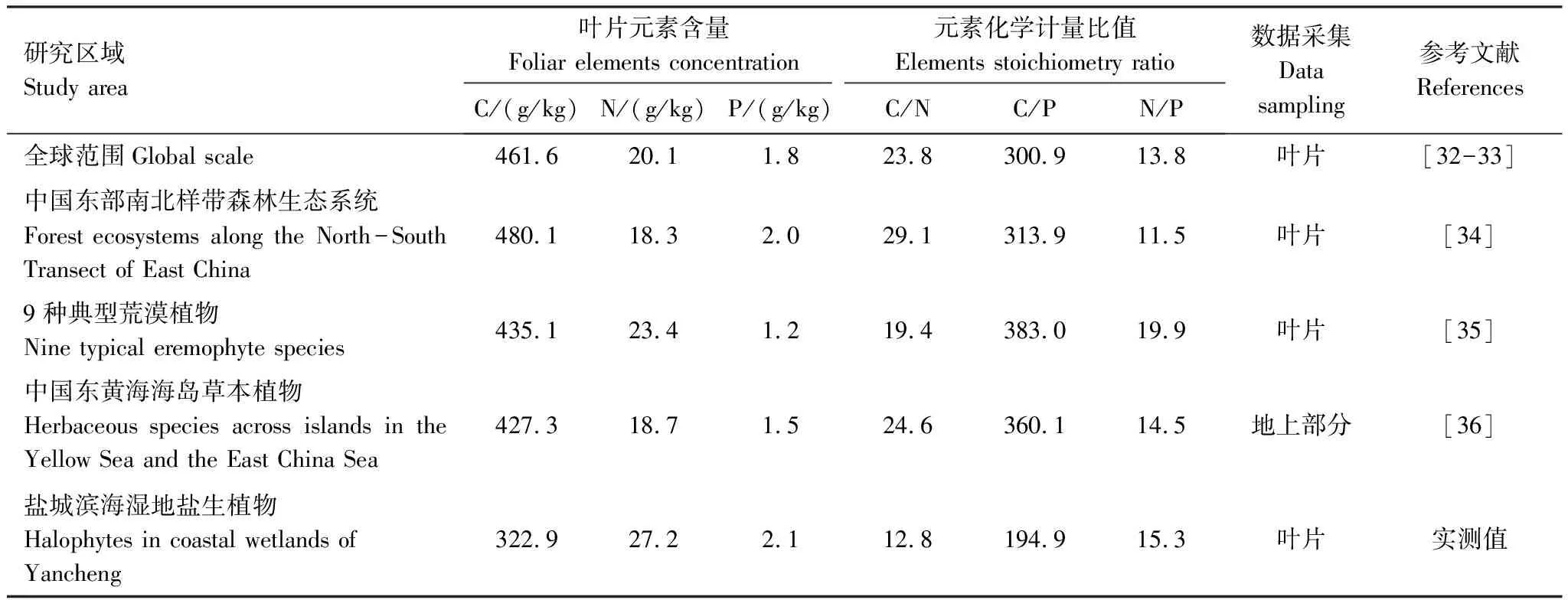
表3 本研究结果与其他区域研究结果列表
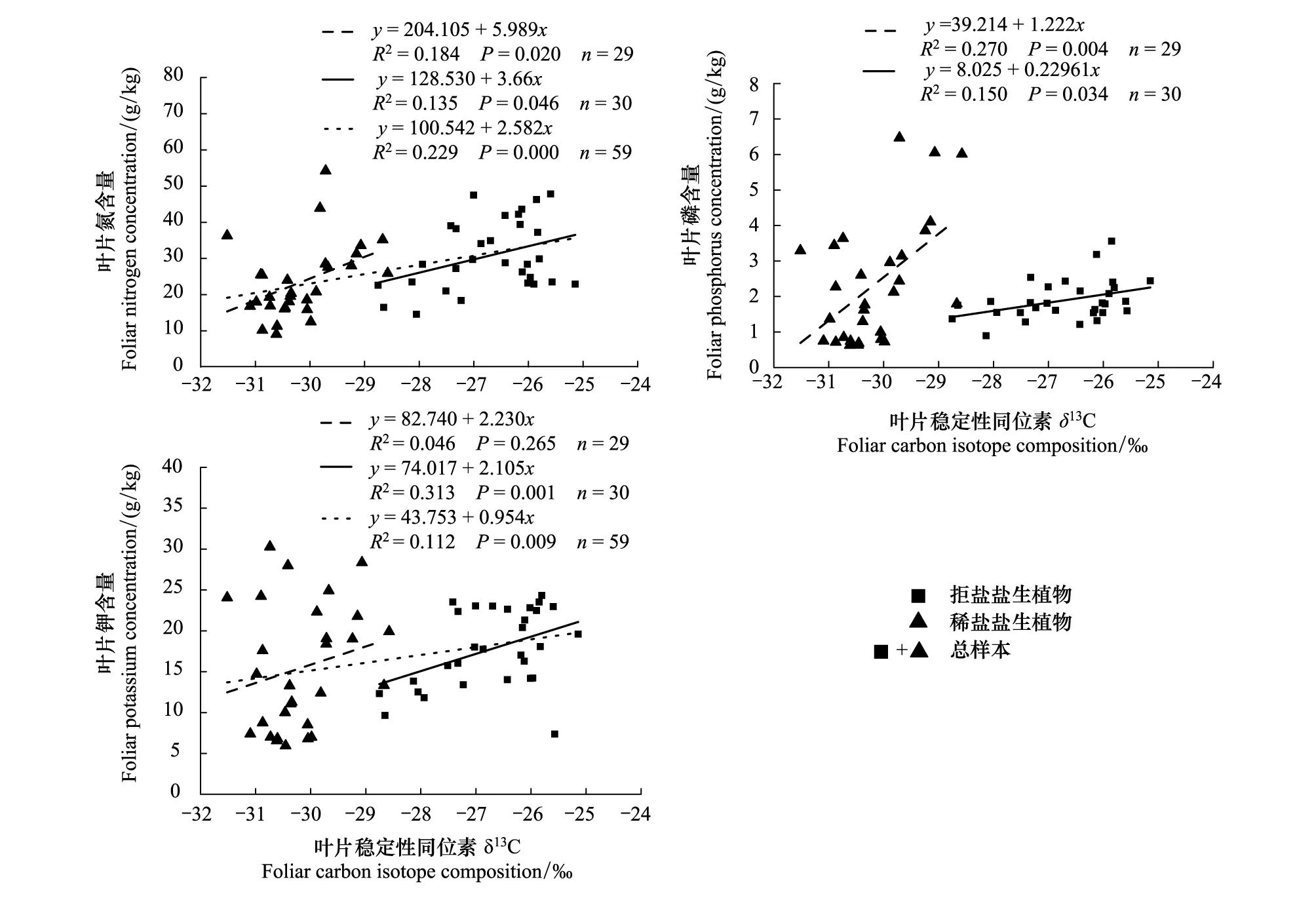
图2 不同盐生植物叶片δ13C与叶片养分元素之间的关系Fig.2 Correlation between foliar δ13C and foliar nutrient elements in different halophytes
2.3 盐生植物叶片δ13C与C/N、C/P、N/P关系
盐生植物总体分析结果表明(图3),δ13C与C/N、C/P与N/P不存在相关性关系(P>0.05)。但在稀盐盐生植物中,δ13C与C/N和C/P比均呈现显著负相关关系(P<0.05),与N/P之间没有相关性(P>0.05)。但对稀盐盐生植物叶片δ13C与N/P关系进行分类回归分析,发现当N/P<16时,δ13C与N/P呈现显著负相关性(P<0.05)。拒盐盐生植物叶片δ13C与C/N存在显著负相关关系(P<0.05),但这种关系在δ13C与C/P和δ13C与N/P分析中不显著(P>0.05)。
3 讨论
3.1 滨海湿地盐生植物叶片δ13C、元素含量及化学计量学特征
本研究得出拒盐盐生植物叶片δ13C显著大于稀盐盐生植物,因WUE与δ13C正相关,见式(4)[9],结果揭示拒盐盐生植物叶片WUE大于稀盐盐生植物。原因可能与植物耐盐特性有关,稀盐盐生植物叶片肉质化发育,能稀释体内过多盐分以缓解生理缺水现象,拒盐盐生植物缺乏这种结构,必须采取高效水分利用手段,才能维持在盐生环境中的生存[1, 5]。
本研究结果得出盐生植物C含量((322.9±83.7) g/kg),均低于各陆生研究区[32- 34]。与前人对滨海湿地生态系统研究相比,研究结果低于中国东黄海海岛植物地上部分含量[36]。这一结果说明该区域盐生植物C储存能力较弱(表3),这与滨海湿地盐生环境压力大、植物生物量累积困难有关[10, 36]。本研究区叶片N、P含量均高于其他区域(表3),可能与盐生植物对资源特殊获取与分配策略有关,盐生植物通过叶片富集大量养分来适应养分亏缺环境,因而会将大量养分储存[16, 36-37]。Talbi-Zribi等[37]对盐生生境3种禾本科饲用植物P利用率进行研究发现,在P亏缺环境,3种草本植物能保持较高P获取能力。郭超等[36]研究认为东黄海海岛5种植物N、P高于杭州湾,与该地区土壤养分亏缺有关,植物通过富集大量养分来适应土壤贫瘠环境。
观测结果显示盐生植物叶片C/N、C/P平均值低于全球尺度及中国其他研究区域(表3)[32- 36],表明滨海湿地生态系统植物单位养分供应的生产潜力较低,与其特殊生境有关,因滨海湿地土壤盐分含量高、长期受养分限制[10-11],植被养分可利用性差,造成植被生产力低下。植物N/P可用于表示养分供给状况,N/P<14表明植物受N限制,N/P>16,表明植物P相对匮乏[11, 24-25]。本研究中,盐生植物N/P平均值为15.3(表3),较全球陆生植物、中国东部南北样带102个优势种及中国东黄海海岛5种常见植物都要高[32- 34, 36],但比典型荒漠植物低[35]。其中稀盐盐生植物N/P<14受N限制、拒盐盐生植物N/P>16受P限制(表2),因生境条件特殊性(高盐环境、土壤贫瘠),江苏滨海盐生植物表现出N、P共同限制的特征。
3.2 不同耐盐型植物叶片δ13C与N、P、K关系
叶片养分元素对植物生长、发育有着重要作用,能通过植物生理过程,如气孔导度(gs)或光合速率(A),直接或间接影响δ13C[18, 20- 22, 26]。因此,探讨盐生植物叶片养分化学计量特征与δ13C(WUE)之间的关系,对于揭示滨海湿地盐生植物适生机理与生长评价指标筛选具有重要意义。本研究得出,植株δ13C随叶片N、P、K含量增加而升高的规律。
研究表明叶片元素N与δ13C在所有盐生植物样本、稀盐盐生植物及拒盐盐生植物中都存在正相关关系(图2),结果与大多数研究结果一致,即δ13C随叶片N浓度的增加而增加,原因是植物光合能力随叶片N浓度的增加而增加,因δ13C与光合能力呈正比,所以叶片N与δ13C正相关[18, 22, 26]。马剑英等[26]对荒漠生态系统红砂(Reaumuriasongarica)植物叶片δ13C与叶片N浓度关系进行研究,发现两者之间存在正相关关系,野外观察还发现在养分限制地点,植株生长稀疏、生物量相对较低,生境条件较好的地点,植物生长较好,说明养分是限制红砂光合生长的因子,进而引起δ13C变化。但Zhou等[21]研究表明,因叶片N合成用于抵御高压、高寒环境,在高海拔地区植物光合能力并未随叶片N浓度的增加而增加,因此在青藏高原低温及低气压环境没有发现叶片N与δ13C之间存在相关关系。
关于叶片P含量与δ13C之间关系的研究存在不一致的结论。马剑英等[26]研究表明红砂植物叶片P与δ13C正相关,因P与光合过程和能量转化过程相关,叶片P含量越多光合能力越强,δ13C随植物光合能力的增强而增大。然而,Zhou等[22]研究锡林郭勒草原植被养分元素与δ13C之间关系时发现,δ13C与P在所有植物样中存在负相关关系,但对物种分类分析时没有发现负相关关系,他们认为P吸收与质流效应有关,其中蒸腾作为P吸收动力,驱使土壤可溶P向根表面移动,蒸腾作用越大WUE越低,δ13C值越小,所以δ13C与P呈反比关系。本研究表明δ13C与叶片P元素在两种耐盐型植物中呈正相关关系,说明在盐生环境下,P通过影响光合产物合成过程影响δ13C,高盐环境导致土壤可溶P低,植物无法通过加大蒸腾作用来富集P。但对所有盐生植物样本进行分析,却没有发现相关性关系,结果与Ma等[18]研究结果一致。表明δ13C与P的关系与植物耐盐类型有关,不同耐盐特性δ13C与P相关性不同,需要对盐生植物类型进行分类分析。
对于盐生植物而言,K+在调节细胞渗透压、缓解Na盐毒害及ROS氧化应激伤害中起着重要作用[3]。Si等[19]认为K+作为渗透调节产物之一,在干旱胁迫中大量积累,以减少细胞渗透势,增大水分利用率,因此K+在干旱区与δ13C正相关。Zhou等[22]发现K+通过蒸腾流被动吸收进入植物组织,被证明与δ13C负相关。本研究得出,δ13C与K+在盐生植物总样本及拒盐盐生植物中均存在极显著正相关关系(P<0.01),这主要与植物叶片积累大量K+,在高盐环境下利于减少细胞渗透势、缓解Na盐毒害及ROS氧化应激伤害有关[2-3, 5, 14, 19]。但在稀盐盐生植物中,这种正相关关系未被发现,原因可能是稀盐盐生植物叶片肉质化发育,叶片内水分会对内部盐溶液进行稀释,依赖K+调节需求比拒盐盐生植物小[1, 5]。
3.3 不同耐盐型植物叶片δ13C与C/N、C/P、N/P关系
研究发现不同耐盐型盐生植物δ13C与C/N呈现负相关关系(图3),表明在滨海湿地高盐胁迫与养分亏缺环境,固定单位C需要获取更多N,即NUE较低,植物必须扩大WUE以代偿减少的N利用率[20, 22-23]。本研究中发现稀盐盐生植物叶片中C/P与δ13C之间存在显著负相关关系,但拒盐盐生植物未发现(图3)。可能与稀盐盐生植物叶未受P限制有关(表2),植物可以增大P利用率以实现低WUE时对C的合成。而拒盐盐生物种中,δ13C与C/P未发现相关性存在,这可能与微生物对P的固定作用大于对植物P供应有关[23],研究缺乏微生物相关研究证据证明。
研究并未发现叶片δ13C与N/P之间存在相关性,Zhou等对青藏高原和锡林郭勒草原研究也并未发现[21-22]。但以N/P=16为分界线,对稀盐盐生植物进行分段分析,发现当N/P小于16时,N/P与δ13C显著负相关。因植物N/P<14, 植物生长主要受N素限制,14 滨海湿地盐生植物受N、P共同限制,盐生植物为适应N、P限制环境,采取N、P光合相关型策略,通过叶片高效吸收与利用N、P元素增加自身光合生产潜力。在盐生环境,K+依赖型渗透调节策略在拒盐植物中更为重要,通过采取K+渗透调节相关型策略,以减小细胞渗透势、增加水分吸收、减轻植物体内盐分毒害。研究结果得出δ13C与C/N成反比,表明盐生植物采取提高WUE以代偿减小的NUE策略。稀盐盐生植物δ13C与C/P成反比,且叶片具有较高P含量,表明稀盐盐生植物可以提高PUE以增大低WUE环境的C合成和生物产量。滨海湿地高盐环境与养分亏缺成为植被恢复的限制因子,本研究结果可为滨海湿地生态系统恢复与重建提供理论依据。4 结论
