大鼠正畸干预时牙移动对牙周组织中Wnt3a、β-catenin蛋白表达的影响及其意义
2020-06-01高再冕李万顺
高再冕,李万顺
(1.平顶山市平煤总医院 口腔科,河南 平顶山467000;2.郑州大学第四附属医院 口腔科,河南 郑州467000)
正畸治疗中牙移动以牙周组织改建为生物学基础[1]。研究发现[2]wnt/β-catenin信号通路对牙周膜细胞分化、功能及牙周膜、牙槽骨生理代谢调控具有非常重要的作用。wnt3a在骨陷窝中表达,属于wnt蛋白一种重要的亚型[3]。通过与细胞膜受体结合激活该通路,是一种潜在的干细胞,对促进内源性干细胞增殖的生长因子具有诱导作用,从而促进细胞的自我更新及组织修复[4]。同时,可抑制糖原合成激酶-3β对β-catenin的磷酸化,促进细胞质中β-catenin的大量蓄积[5]。β-catenin是一种具有细胞黏附作用以及转录调节作用的多功能蛋白,在经典wnt/β-catenin信号通路中发挥着关键的作用[6,7]。本研究通过探索正畸牙齿过程中加力1、3、5、7、10、14天后张力区、压力区Wnt3a、β-catenin蛋白、 mRNA表达的变化来证实wnt3a、β-catenin对成骨细胞分化、骨量调节的作用及在正畸牙齿移动过程中对牙周组织改建的影响。现将报道如下。
1 材料与方法
1.1 实验动物
选取7周龄健康雄性SD大鼠48只,体质量220-250 g,喂养于自由进食进水、光照12 h、室温23℃-25℃、相对湿度45%-60%的环境中;所有实验大鼠购买自南方医科大学动物实验研究中心。
1.2 动物分组及干预方法
建立大鼠正畸治疗中牙移动的模型,选择7周龄、220-250克左右健康SD大鼠48只(均由哈尔滨医科大学动物实验中心提供),根据加力后第1、3、5、7、10、14天进行分组,每组8只。麻醉采用10%水合氯醛腹腔注射,将麻醉后大鼠固定。用结扎丝将大鼠左上颌第一磨牙与中切牙之间结扎并拴上施加50克力的镍钛螺旋拉簧。将第一磨牙拉向正中,以上颌切牙轴角近牙龈处拉出0.5 mm左右的深槽沟并用树脂固定。对照组为右侧未安装正畸加力装置组。
1.3 组织制备
全麻后采用心脏灌注的方式将大鼠处死,将上颌两侧第一磨牙及其牙周组织完整保留。用4%的多聚甲醛固定48 h后用15%ph=7.4的EDTA微波脱钙6周,流水冲洗24 h,再用系列乙醇-正丁醇脱水,二甲苯透明,浸蜡。包埋面定位上颌三颗磨牙腭侧面,以平行于上颌磨牙牙体长轴为标准,近、远、中向连续切片,切片厚度为4 μm,并进行免疫组化检测。
1.4 免疫组化检测
兔抗大鼠多克隆抗体1∶100由北京博奥森生物试剂公司提供,sv0002-兔IgG两步法免疫组化检测试剂盒及DAB显色盒均购自武汉博士德生物公司。操作步骤严格按照试剂盒及显色盒的规定进行。选取各个时间点第一磨牙近中根近中、远中牙周膜的高倍镜视野进行摄像,所得摄像存于Image-ProPlus6.0系统进行图像分析,计算各组标本阳性染色区域的平均光密度。
1.5 逆转录-PCR法
1.5.1总RNA的提取 取大鼠第一磨牙压力侧、左侧上颌第一磨牙、右侧上颌第一磨牙牙槽骨组织均100 mg放入研磨钵中加液氮进行研磨。取组织粉末至于1.5 ml的EP管中加1 ml Trizol剧烈震荡15 s后室温条件下静置5-10 min加200 ml氯仿在12 000 rpm的条件下离心15 min,取400-500 μl上层水相加入0.5 ml乙丙醇充分混匀,在12 000 rpm条件下离心10 min弃异丙醇。加1 ml 75%乙醇混匀在8 000 rpm离心条件下进行离心5 min倒掉乙醇。滤纸上晒干5 min得到总RNA。
1.5.2逆转录合成cDNA 体系如下:5 μl的5×Reaction Buffer、3 μl 10 mM dNTP Mixture、1 μl oligodT、1 μl RNase Inhitor、1 μl Reverse Transcriptase 、3 μg Total RNA、14 μl DEPC H2O混匀后快速离心1次放入PCR仪中。逆转录反应条件如下:40℃1 h、70℃5 min置于-20℃条件下冰箱保存。PCR反应体系具体为:10 μl 2×Taq PCR Master Mix、0.15 μl rTaq酶、1 μl 上游引物、1 μl 下游引物、3 μl 反转录反应液、4.85 μl ddH2O PCR反应条件为35个循环,95℃30 s,58℃20 s,72℃25 s,GAPDH为内参物,最终得到wnt3a、β-catenin mRNA扩增产物。
1.6 统计学方法
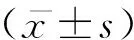
2 结果
2.1 不同组别大鼠牙周组织的Wnt3a、β-catenin蛋白表达情况
不同时间组别压力区、张力区大鼠牙周组织的Wnt3a、β-catenin蛋白表达差异有统计学意义(P<0.05),均随时间的推移呈现先升高后降低趋势,压力区和张力区β-catenin蛋白表达分别于加力后第5天和第7天达高峰,而Wnt3a蛋白表达均于加力后第5天达高峰,见图1,图2;不同时间组别对照区Wnt3a、β-catenin蛋白表达差异无统计学意义(P>0.05)。A、B、C、D、E、F组张力区的Wnt3a、β-catenin蛋白表达均显著高于对照区和压力区(P<0.05); C、D、E、F组压力区的Wnt3a、β-catenin蛋白表达均显著高于对照区(P<0.05),见表1、表2。
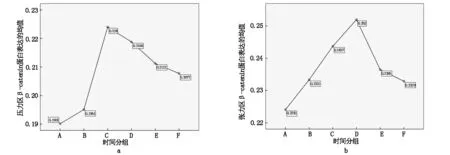
图1 不同组别大鼠牙周组织的β-catenin蛋白表达
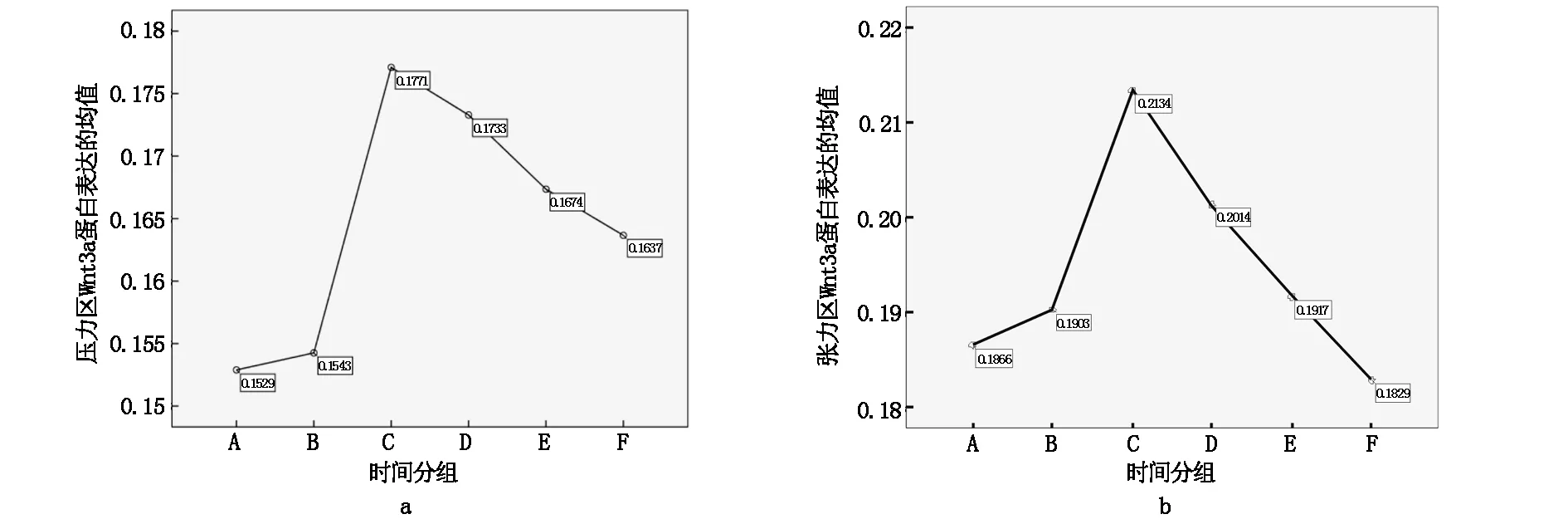
图2 不同组别大鼠牙周组织的Wnt3a蛋白表达

表1 不同组别大鼠牙周组织的β-catenin蛋白表达情况光密度值)
注:与同组对照区比较,*P<0.05;与同组压力区比较,#P<0.05。与同区C组比较,△P<0.05;与同区D组比较,□P<0.05
2.2 不同组别大鼠牙周组织的Wnt3a、β-catenin mRNA表达情况
不同时间组别压力区、张力区大鼠牙周组织的Wnt3a、β-catenin mRNA表达差异有统计学意义(P<0.05),均随时间的推移呈现先升高后降低趋势,压力区和张力区β-catenin mRNA表达均于加力后第7天达高峰,而Wnt3a mRNA表达均于加力后第5天达高峰,见图3,图4;不同时间组别对照区Wnt3a、β-catenin蛋白表达差异无统计学意义(P>0.05)。A、B、C、D、E、F组张力区的Wnt3a、β-catenin mRNA表达均显著的高于对照区和压力区(P<0.05); C、D、E、F组压力区的Wnt3a、β-catenin mRNA表达均显著的高于对照区(P<0.05)。Wnt3a、β-catenin mRNA表达电泳图见图5、图6。
表2 不同组大鼠牙周组织的Wnt3a蛋白表达情况光密度值)
注:与同组对照区比较,*P<0.05;与同组压力区比较,#P<0.05。与同区C组比较,△P<0.05

图3 不同组别大鼠牙周组织的β-catenin mRNA表达

图4 不同组别大鼠牙周组织的Wnt3a mRNA表达

图5 各组不同区域β-cateninPCR电泳图
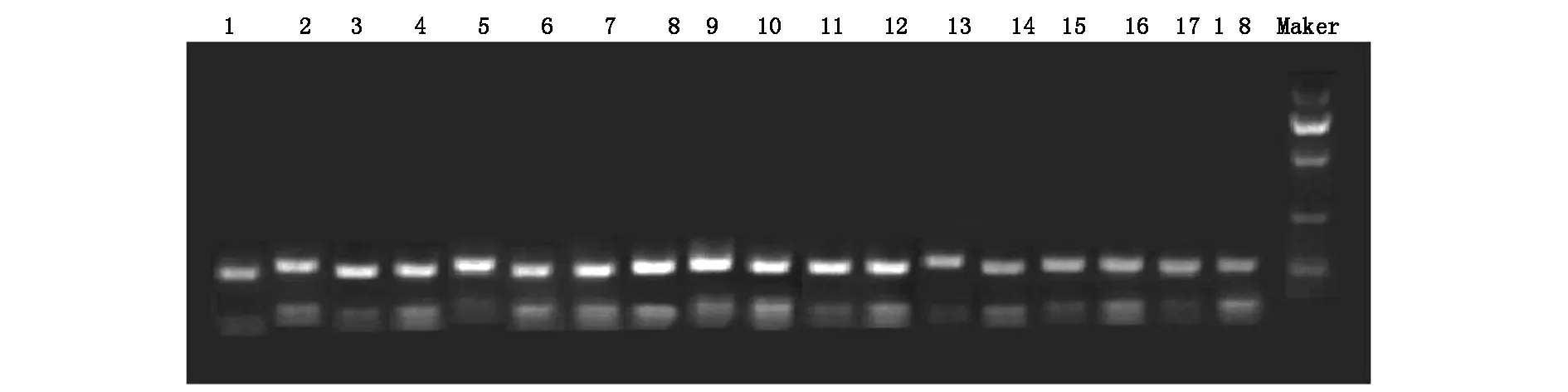
图6 各组不同区域Wnt3am PCR电泳图
表3 不同组别大鼠牙周组织的β-catenin mRNA表达情况相对表达)
注:与同组对照区比较,*P<0.05;与同组压力区比较,#P<0.05。与同区D组比较,△P<0.05
表4 不同组别大鼠牙周组织的Wnt3a mRNA表达情况相对表达)
注:与对照区比较*P<0.05,与压力区比较#P<0.05;与同区C组比较,△P<0.05
3 讨论
正畸作用下压力区牙周膜受压变窄常会引起破骨细胞活跃,并促进相关因子表达,从而增强骨吸收[8]。临床研究发现[9]正畸过程中加注wnt3a信号通路抑制剂DKK1阻断wnt信号通路。主要机制是通过直接或间接手段与wat蛋白竞争性结合低密度脂蛋白受体相关蛋白,从而使信号向细胞内传递的能力减弱,骨形成减少。说明干扰wnt蛋白通路可影响破骨细胞诱导骨吸收程度,进一步阻碍骨重建。wt3a是want信号通路重要的Wnt蛋白亚型,对成骨细胞增殖、分化、成熟起着重要的调控作用,同时还可以促进成骨细胞分泌相关因子发挥对破骨细胞分化的诱导作用,对促进成骨、抑制骨吸收均具有重要意义。外在机械力作用成骨细胞影可响骨改建[10]。有研究发现[11]牙周复合体中某些细胞能够响应wnt刺激。通过激活或抑制wnt/β-catenin信号通路能导致牙周膜中成骨因子的表达水平的改变,从而对牙周膜宽度及牙槽骨更新造成影响。此外,wnt3a作为经典wnt蛋白的配体,通过对成骨细胞分化的直接诱导作用、对破骨细胞分化的矿化及抑制作用以及相关功能的表达来调控骨改建。研究结果显示A、B、C、D、E、F组张力区的Wnt3a、β-catenin蛋白表达均显著的高于对照区和压力区(P<0.05); C、D、E、F组压力区的Wnt3a、β-catenin蛋白表达均显著的高于对照区(P<0.05)与李争争等发现[12]wnt3a蛋白在骨折愈合过程损伤部位的表达上调结果一致。说明wnt/β-catenin信号通路通过上调wnt3a表达提高骨损伤修复能力。wnt3a在张力区表达上调程度显著。其原因主要是由于wnt3a作为促进成骨细胞形成且向骨细胞分化及抑制骨细胞吸收的重要want蛋白亚型,在机械力刺激的条件下表达增强。张力区牙周膜受牵拉增宽,成骨相关转录因子表达增强。本研究结果显示Wnt3a表达上调,可充分说明上调wnt3a mRNA水平使wnt3a分泌增加,促进骨重建。相关研究证实[13]基质金属蛋白酶-13(MMP-13)是骨细胞分化过程中重要的物质,可调控胶原分泌及骨基质成分组成,最终影响骨改建。经典wnt/β-catenin信号通路、骨形态蛋白-2是MMP-13重要的调控蛋白。其水平的增加能够促进成骨细胞的分泌与本研究结果一致。
β-catenin信号通路受wnt蛋白家族的影响,wnt蛋白通过与受体结合后可抑制β-catenin被磷酸化,使其在细胞质中大量蓄积,并转移至细胞核内与相应转录因子相互作用,从而激活下游靶基因转录。β-catenin是内源性骨量调节者,同时也是具有细胞间黏附和转录调控作用的多功能蛋白。本研究结果显示β-catenin蛋白及其mRNA表达上调,充分说明其在正畸牙齿移动牙周组织改建过程中意义重大。其机制主要通过经典wnt/β-catenin信号转导通路来实现。Wnt3a、β-catenin可调节成骨细胞中护骨素(OPG)的表达与杨先炯等报道[14]间充质细胞中want3a激活的经典wnt信号途径使OPG表达上调结果一致。更有体外研究证明[15]缺乏 β-catenin可致成骨细胞成熟和矿化受阻,OPG含量下降,破骨细胞活跃,不利于骨重建。由此推断Wnt信号通过影响OPG生成,决定牙周疾病进展及牙槽骨组织的改建。
综上所述,大鼠行正畸治疗中牙移动中,Wnt3a、β-catenin mRNA蛋白表达上调,参与牙周组织重建,并且主要于第5天、第7天达到高峰。本研究的创新之处在于,通过分析正畸牙齿移动过程中加力1、3、5、7、10、14天后张力区、压力区Wnt3a、β-catenin蛋白、 mRNA表达的变化,进而证实Wnt3a、β-catenin对成骨细胞分化、骨量调节的重要作用。确定其在正畸牙齿过程中牙周组织改建中的影响,对正畸效果的评价提供科学的指标。
