调控植物种子大小的分子机制综述
2020-05-21曹维赵静禹艳坤
曹维 赵静 禹艳坤
摘要:植物种子大小对农业生产和植物自身的发展尤为重要,迄今为止已鉴定出大量的调控种子大小的基因,它们作用于不同的途径,以调节种子不同结构(种皮、胚乳和胚)的生长,从而协调地控制种子大小。从泛素-蛋白酶体途径、HAIKU(IKU)途径、植物激素途径、丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)信号途径以及G蛋白信号途径和转录调控因子等方面综述调控种子大小的基因的分子机制,以及某些途径之间可能存在的联系,以期为探索新的种子大小调控基因和进行分子设计育种工作提供切实可用的理论基础。
关键词:种子大小;分子机制;调控基因;调控途径
中图分类号:Q943.2 文献标志码:A 文章编号:1002-1302(2020)06-0001-07
种子大小是农业生产中最受关注的农艺性状之一,是决定产量的重要因素,同时种子大小对植物的进化至关重要,大种子能够积累足够的营养物质用于发芽,并且对非生物胁迫具有更好的耐受性,而小种子有利于分散和繁殖大量的后代[1-2]。
在各物种中,种子大小差异非常大。然而,关于植物是如何调控其种子大小的研究尚未见系统的报道。在被子植物中,一个精细胞与卵融合形成二倍体的合子,另一个精细胞与中央细胞融合形成初生胚乳核,双受精后由合子发育成胚,中央细胞则发育成胚乳,而胚珠的珠被发育成种皮[3]。因此,种子大小主要由来自孢子体组织和合子组织的遗传信息所控制,但也会受到环境因素的影响[4]。一些控制种子大小的基因已经在模式植物拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa L.)中被鉴定出来了,它们主要参与泛素-蛋白酶体途径、HAIKU(IKU)途径、植物激素途径、丝裂原活化蛋白激酶(MAPK)信号途径、G蛋白信号途径和转录调控因子等途径。
1 泛素-蛋白酶体途径
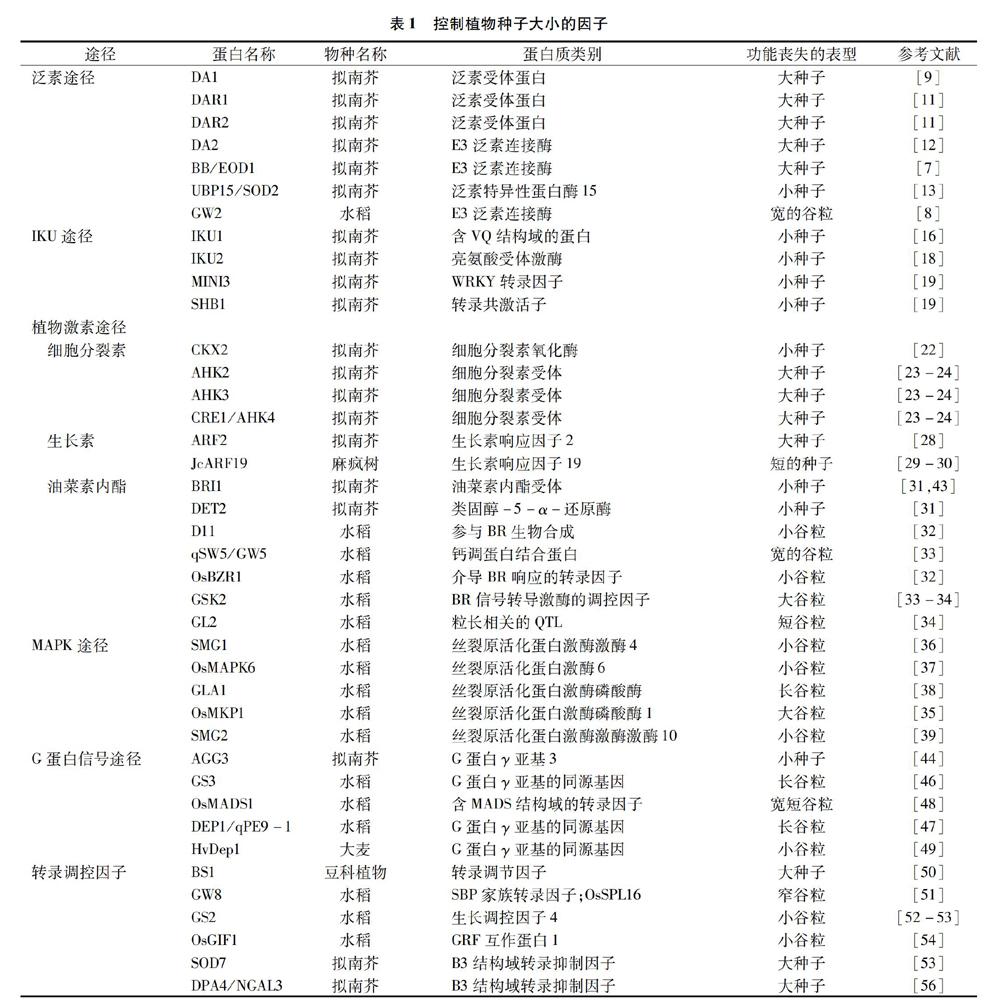
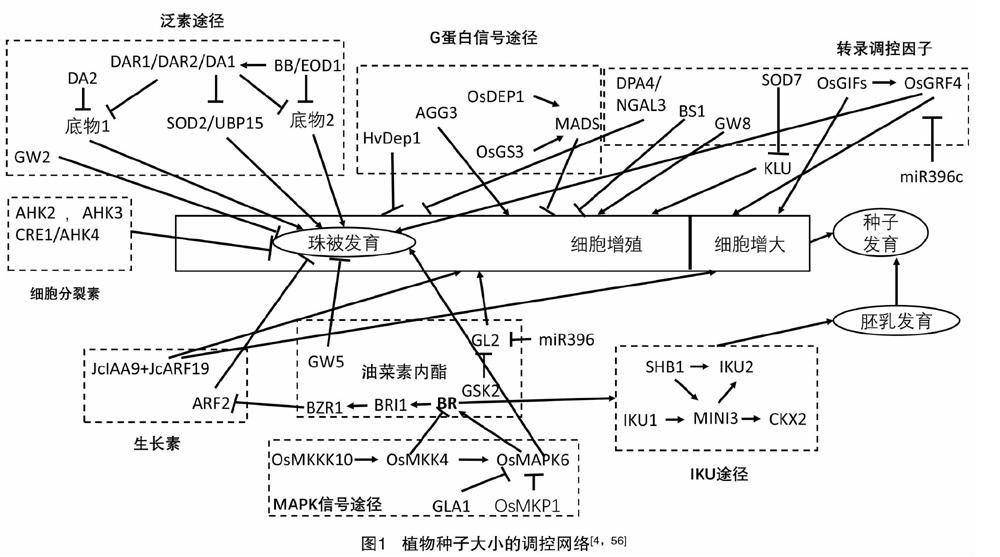
泛素途径在植物种子大小的确定中起重要作用[5-8]。在拟南芥中,DA1编码1个泛素受体,含有2个泛素互作基序(UIMs)和1个结合单链的LIM结构域[6]。DA1蛋白中第358位精氨酸-赖氨酸的突变(DA1R358K)产生了da1-1突变体,da1-1突变体产生的种子比野生型的更大且更重,这是由于促进了孢子体珠被细胞的增殖而产生了较大的珠被[6,9]。AtDA1在甘蓝型油菜(Brassica napus L.)中的同源基因BnDA1,也具有负调控种子和器官大小的功能[10]。DA1-related protein 1(DAR1)和 DA1-related protein 2(DAR2)与DA1蛋白的氨基酸序列很相似,它们在限制种子和器官生长的功能上是冗余的[11]。
拟南芥中的2个RING型E3泛素连接酶,DA2 和BIGBROTHER (BB)/ENHANCER OF DA1 (EOD1)是种子大小的负调控因子,DA2或者BB/EOD1的过量表达都能引起植株器官变小,虽然它们都通过限制珠被细胞的增殖来调控种子大小,但是在控制种子大小的途径中它们不能相互作用;此外,eod1和da2-1的突变协同地增强了da1-1突变体的种子大小表型,表明DA2和EOD1可能通过DA1进行相同的调控来降解不同的生长刺激因子[7,12]。DA1的一个抑制子SUPPRESSOR OF DA1(SOD2),编码去泛素化酶UBIQUITIN-SPECIFIC PROTEASE 15 (UBP15),UBP15作用于DA1的下游,它是种子和器官生长的正调控因子[8,13]。在水稻中,1个谷粒宽度和质量的数量性状位点GRAIN WIDTH AND WEIGHT 2 (GW2),编码1个Really Interesting New Gene (RING)型E3泛素连接酶,在拟南芥中过量表达GW2产生了小的种子和器官;与GW2的RING结构域相比,DA2的RING结构域中一个保守的His残基被Asn残基(Asn-91)取代[7]。
SAPSTERILE APETALA (SAP)/SUPPRESSOR OF DA1 (SOD3)是一種F-box蛋白,参与构成一种SKP1/Cullin/F-box E3泛素连接酶复合物,它通过促进分生组织细胞的增殖来控制器官大小。此外,SAP能够降解PEAPOD1 和 PEAPOD2(分生组织增殖的负调控因子)以控制器官大小[14]。KIX8和KIX9是SAP的底物,SAP与KIX8/KIX9相互作用并调节其蛋白质稳定性,从而通过调控分生组织细胞的增殖来控制器官生长[15]。
2 IKU途径
胚乳发育是决定种子大小的一个重要因素[16],HAIKU(IKU)途径控制植物种子胚乳的早期生长,IKU途径包括1个富含亮氨酸的重复受体激酶IKU2和1个WRKY转录因子MINI-SEED3(MINI3)[17]。
HAIKU1(IKU1)编码一个含有VQ基序(植物所特有的)的蛋白,遗传分析表明VQ基序是IKU1调控种子大小的不可缺少的元件。IKU1在发育早期的胚乳和中央细胞中表达,iku1突变体产生了具有少量胚乳的小种子[16]。在iku1功能缺失突变体中IKU2和MINI3的表达量均减少,在mini3突变体中IKU2的表达水平降低,而iku2突变体中MINI3的表达量没有变化,表明IKU1、IKU2和MINI3作用于同一信号途径并且通过影响胚乳的生长来控制种子大小[18]。此外,SHORT HYPOCOTYLUNDER BLUE 1 (SHB1)能够结合IKU2和MINI3的启动子以调控它们的表达[19]。MINI3结合cytokinin oxidase/dehydrogemase2(CKX2)的启动子并促进其表达从而调控胚乳的生长[20]。因此,胚乳发育过程将IKU途径与细胞分裂素信号途径相关联。
3 植物激素途径
3.1 细胞分裂素
细胞分裂素水平受到生物合成[异戊烯基转移酶(IPT)]、激活[Lonely Guy(LOG)]、失活(O-葡萄糖基转移酶)、再激活(β-葡萄糖苷酶)和降解[细胞分裂素氧化酶/脱氢酶(CKX)]等之间的平衡调节。在果实和种子发育的早期阶段,细胞分裂素水平暂时升高,并与细胞核和细胞分裂一致,细胞核和细胞分裂是种子最终大小的决定因素。细胞分裂素的外源性应用,IPT的异位表达或CKX的下调有时会使种子产量增加,表明细胞分裂素可能限制产量[21]。现代基因组编辑工具可用于靶向和操纵细胞分裂素水平以增加种子产量。
CKX2影响细胞分裂素信号转导,而且是IKU途径的靶向基因,CKX2的表达直接受到IKU转录因子WRKY10的激活然后促进胚乳的生长;同时CKX2的表达还依赖于H3K27me3的沉积,其因母体基因组剂量不平衡和雄配子的DNA去甲基化而产生波动[20]。3种Arabidopsis histidine kinases(AHKs),AHK2、AHK3和CRE1(cytokinin response1)/AHK4是细胞分裂素受体,它们以依赖细胞分裂素的方式负调控拟南芥种子大小,它们形成的三突变体的种子比野生型的大2倍多[22-23]。
3.2 生长素
植物激素生长素,通过调节转录因子与其下游基因的生长素响应元件(auxin-response elements,AuxREs)的相互作用来调控植物生理[24]。AUXIN RESPONSE FACTORS (ARFs)和Aux/IAAs调控生长素响应基因的表达,大多数ARF蛋白在N末端含有高度保守的B3-like DNA结合结构域,其识别生长素响应基因启动子中的生长素应答元件(AuxRE:TGTCTC),而C末端结构域包含2个基序,称为Ⅲ和Ⅳ,在Aux/IAA蛋白中也发现了这2个基序,并且在ARF和Aux/IAAs中能形成同源二聚体和异源二聚体[25-26]。
拟南芥AUXIN RESPONSE FACTOR 2 (ARF2)是一个转录抑制因子,纯合的arf2 T-DNA插入突變体的种子比野生型的大[27]。在木本植物麻疯树(Jatropha curcas L.)中,JcARF19的异位表达上调了某些生长素响应基因,这些基因参与编码种子发育过程中细胞分化和细胞骨架形成,因此JcARF19的过量表达显著增加了拟南芥和麻疯树种子的大小和产量[28]。此外,在麻疯树中ARF19和IAA9相互作用,能影响种子的长度[29]。
3.3 油菜素内酯
油菜素内酯(brassinosteroids,BR)通过激活许多可能增加种子大小的基因表达以促进种子变大,并且抑制负调控种子生长基因的表达,当用BR处理3 h后,种子大小的正调控因子SHB1、IKU1、MINI3和IKU2的表达水平明显升高[30]。Carbon Starved Anther(CSA)基因编码1个MYB结构域蛋白,BRs通过上调其表达来促进水稻花粉和种子的发育;BR合成基因DWARF 11(D11)或BR信号因子Oryza sativa BRASSINAZOLE-RESISTANT1 (OsBZR1)的低表达会导致花粉败育以及种子大小和质量的减小,并积累较少的淀粉,因此,OsBZR1促进CSA的表达,而CSA又调控下游糖类分配和代谢基因的表达[31]。BR-deficientmutant(det2)和BR receptor mutant(bri1-5,bri1突变体的一个弱等位基因)的种子均比野生型的小且轻,形状上短而略宽[30]。水稻SEED WIDTH ON CHROMOSOME 5(qSW5/GW5)基因编码钙调蛋白结合蛋白,它可以与水稻glycogen synthase kinase 2(GSK2)互作并抑制其激酶活性,导致未磷酸化的OsBZR1和dwarfand low-tillering(DLT)蛋白的积累,从而调控油菜素内酯响应的基因表达和生长反应(包括谷粒宽度和质量)[32]。grain-length-associated QTL、GL2在其miR396靶向序列中含有突变,导致它的表达水平适度增加,从而可以通过上调大量的BR诱导基因来激活BR反应以促进谷粒发育;此外,GSK2能够直接与GL2相互作用并抑制其转录激活活性,从而介导激素对谷粒长度的特异性调节[33]。
4 MAPK信号途径
MAPK级联包含至少3种激酶:MAPK激酶激酶(MKKK),MAPK激酶(MKK)和MAPK。MAPK信号通路在防御反应以及与植物生长和发育有关的多个过程中起着重要作用。
LARGE GRAIN 8(LARGE8)编码mitogen-activated protein kinase phosphatase 1(OsMKP1),它通过限制小穗壳细胞的增殖来影响谷粒大小,过表达OsMKP1时会产生小的谷粒;同时OsMKP1能直接与mitogen-activated protein kinase 6(OsMAPK6)相互作用并使其失活,表明OsMAPK6的可逆磷酸化在确定谷粒大小中起着重要的作用[34]。SMALL GRAIN 1(SMG1)编码mitogen-activated protein kinase kinase 4(OsMKK4),smg1突变体由于细胞增殖缺陷而具有小而轻的谷粒、致密且直立的圆锥花序和相对较矮的植株;此外OsMKK4可以抑制BR反应以及BR相关基因的表达,表明MAPK途径与BRs在谷粒生长中可能存在联系[35]。dwarf and small grain1(dsg1)突变体具有小谷粒、植株矮小和叶片直立的表型,DSG1编码OsMAPK6,OsMKK4可能是OsMAPK6的上游MAPK激酶,dsg1突变体中的内源性BR水平降低了,并且当不论是否施加外源性BR时,dsg1突变体中几种有关BR信号转导的基因和反馈抑制基因的表达均被改变,表明OsMAPK6可能影响BR稳态和信号转导[36]。一个隐性突变基因grain length and awn 1 (gla1)的突变位于编码MAPK磷酸酶基因中的单核苷酸多态性(single-nucleotide polymorphism,SNP),GLA1蛋白与OsMAPK6相互作用,通过OsMAPK6的去磷酸化来控制谷粒大小,GLA1的过表达会导致谷粒长度和质量减小,表明GLA1可以作为OsMAPKK4-OsMAPK6级联的负调节因子[37]。SMALL GRAIN 2(SMG2)编码mitogen-activated protein kinase kinase kinase 10(OsMKKK10),smg2突变体具有小而轻的谷粒、短穗和半矮秆植株,OsMKKK10与OsMKK4相互作用并使其磷酸化,综上所述OsMKKK10、OsMKK4和OsMAPK6可能是作为一个级联调控谷粒大小[38],并且MAPK途径和BRs在谷粒生长中可能存在一定联系。
5 G蛋白信号途径
水稻的异源三聚体G蛋白复合物含有1个α亚基基因RICE G-PROTEIN ALPHA SUBUNIT(RGA1),4个GTP结合蛋白基因eXtra Large GTP-binding proteins(XLGs),1个β亚基基因RICE G-PROTEIN BETA SUBUNIT (RGB1)和5个γ亚基基因[暂定名为Rice heterotrimeric G-protein gamma subunit1(RGG1),Rice heterotrimeric G-protein gamma subunit2(RGG2),Rice heterotrimeric G-protein gamma subunit 3(RGG3)/GRAIN SIZE 3 (GS3),Rice heterotrimeric G-protein gamma subunit 4(RGG4)/DENSE AND ERECT PANICLE1 (DEP1)/DENSE PANICLE1 (DN1) 和Rice heterotrimeric G-protein gamma subunit 5(RGG5)][39]。在最近发现的一个由异源三聚体G蛋白的5个亚基组成的通路中,Gβ蛋白对植物的存活和生长至关重要,Gα为谷粒增大提供了基础,3种Gγ蛋白DEP1、GGC2和GS3拮抗地调节谷粒大小,其中DEP1和GGC2都可以在与RGB1形成复合物时增加谷粒的长度[40]。RGA1基因的功能丧失会引起水稻植株的形态异常:直立的叶子和小而圆的种子[41]。此外,水稻Gα影响BR信号级联,但Gα可能不是BRI1介导的转导中的信号分子[42]。
植物特异性G蛋白γ亚基ARABIDOPSIS G-PROTEIN GAMMA SUBUNIT 3(AGG3)通过增加拟南芥的增殖生长期来促进种子和器官生长,同时AGG3包含跨膜结构域,它位于质膜中并与功能性G蛋白α亚基G-protein α subunit(GPA1)和G蛋白β亚基Arabidopsis G-Protein β Subunit 1(AGB1)相互作用[43]。当将拟南芥G蛋白γ亚基AGG3在亚麻荠中用组成型启动子或种子特异性启动子过表达时,每棵植株的种子大小、种子质量和种子数量均增加15%~40%[44]。AGG3在水稻中的同源基因GS3和DEP1/qPE9-1已被确定为种子大小和产量的重要数量性状位点,GS3通过限制细胞增殖来影响种子和器官生长[45]。DEP1/qPE9-1蛋白的N末端含有G gamma-like(GGL)结构域,GGL负调控谷粒的长度和质量,而C末端的von Willebrand factor type C(VWFC)结构域可以抑制GGL的负调控作用。但只有1个VWFC结构域时不能改变GGL结构域对谷粒大小的抑制[46]。水稻谷粒产量QTL qLGY3编码MADS结构域转录因子OsMADS1,它是G蛋白βγ二聚体关键的下游效应蛋白;Gγ亚基GS3和DEP1直接与MADS转录因子保守的角蛋白样结构域相互作用,以增强OsMADS1的转录活性并促进共同靶基因的协同反式激活,从而调节谷粒的大小和形状[47]。Hordeum vulgare Dense anderectpanicle1(HvDep1)是AGG3型亚基的编码基因,它在大麦(Hordeum vulgare L.)中具有功能丧失性质的突变,这可以正调节大麦的茎伸长和种子大小,但是HvDep1对大麦产量的影响是受基因型和环境共同调节的[48]。
6 转录调控因子
转录因子在调控种子大小中起着重要的作用。BIG SEEDS1(BS1)基因编码植物特异性转录调节因子,通过抑制原代细胞的增殖来调控植物器官大小(包括种子、种荚和叶片)[49]。GRAIN WEIGHT8(GW8)编码SQUAMOSA promoter binding protein (SBP)家族转录因子SQUAMOSA PROMOTER BINDING PROTEIN-LIKE 16(OsSPL16),该基因的高表达促进细胞分裂和谷粒灌浆,对水稻的谷粒宽度和产量具有积极影响,此外GW8的谷粒大小和等位基因变异之间的相关性表明,在水稻育种计划中可能选择了启动子区域内的突变[50]。
在水稻中GRAIN SIZE 2(GS2)编码RICE GROWTH REGULATING FACTOR 4(OsGRF4),定位于细胞核并可作为转录激活因子,一种罕见的突变影响了GS2的microRNA(miR396c)结合位点:第3个外显子中一个2 bp的突变(1187TC→AA),导致它的表达水平升高,产生了更大的细胞并且增加了细胞数量,从而提高了谷粒质量和产量[51-52]。Growth-regulating factor interacting factor1(OsGIF1)通过调控细胞大小来影响叶片、茎和谷粒的大小,此外它也影响水稻的繁殖[53]。OsGIF1直接与OsGRF4相互作用,并且上调其表达水平从而增加谷粒大小,综上所述,miR396c-OsGRF4-OsGIF1调控机制在谷粒大小的确定中起着重要作用,并对水稻产量的提高具有重要意义[54]。
由SUPPRESSOR 7 OF DA1(SOD7)编码的B3 domain transcriptional repressor NGATHA-like protein (NGAL2),通过限制珠被和发育中种子中的细胞增殖来调节种子大小[52]。DEVELOPMENT-RELATED PcG TARGET IN THE APEX4 (DPA4/NGAL3)和SOD7與种子大小调节因子KLUH(KLU)在同一途径中起作用以调节种子生长,但独立于DA1,同时SOD7直接结合KLUH(KLU)的启动子并抑制KLU的表达[55]。
7 结论与展望
植物种子大小受到珠被、胚乳和胚的协同控制,同时这些组织的发育又被各种分子机制调节着(表1)。如图1所示,泛素-蛋白酶体途径和MAPK信号途径主要影响珠被的大小,IKU途径主要调控胚乳的发育等;并且控制种子大小的几个途径之间可能会通过一些基因或植物激素构成联系,例如生长素信号转导途径和BR信号转导途径(BZR1与ARF2),IKU途径和细胞分裂素信号途径(MINI3与CKX2),BR信号转导途径和MAPK途径(OsMKK4、OsMAPK6与BR相关基因),G蛋白途径和BR信号转导途径(Gα影响BR信号级联)等。尽管如此,调控种子大小的分子机制仍然不够清楚,各種途径之间的联系甚少,还不能阐明珠被、胚乳和胚之间的发育是如何相互协调的。
在经济作物中,探索调控种子大小的基因意义重大,可以直接有效地提高产量,然而很多已经被鉴定出的基因或QTL并不能直接用于改造已有品种,对促进分子设计育种、提高作物产量和质量的作用并不大。因此须要不断探索新的功能基因或是已知基因的新功能,同时筛选出切实可用的分子标记,将研究工作真正落实到生产中。
参考文献:
[1]Westoby M,Falster D S,Angela T M,et al. Plant ecological strategies:some leading dimensions of variation between species[J]. Annual Review of Ecology and Systematics,2002,33(1):125-159.
[2]Moles A T,Ackerly D D,Webb C O,et al. A brief history of seed size[J]. Science,2005,307(5709):576-580.
[3]刘春明,程佑发,刘永秀,等. 植物种子发育的分子机理[J]. 中国基础科学,2016,18(2):1-13.
[4]Li N,Li Y H. Signaling pathways of seed size control in plants[J]. Current Opinion in Plant Biology,2016,33:23-32.
[5]Song X J,Huang W,Shi M,et al. A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase[J]. Nature Genetics,2007,39(5):623-630.
[6]Li Y,Zheng L,Corke F,et al. Control of final seed and organ size by the DA1 gene family in Arabidopsis thaliana[J]. Genes & Development,2008,22(10):1331-1336.
[7]Xia T,Li N,Dumenil J,et al. The ubiquitin receptor DA1 interacts
with the E3 ubiquitin ligase DA2 to regulate seed and organ size in Arabidopsis[J]. The Plant Cell,2013,25(9):3347-3359.
[8]Du L,Li N,Chen L,et al. The ubiquitin receptor DA1 regulates seed and organ size by modulating the stability of the ubiquitin-specific protease UBP15/SOD2 in Arabidopsis[J]. The Plant Cell,2014,26(2):665-677.
[9]夏 天,李 娜,李云海. 植物种子和器官大小调控研究取得新进展[J]. 遗传,2013,35(10):1208.
[10]Wang J L,Tang M Q,Chen S,et al. Down-regulation of BnDA1,whose gene locus is associated with the seeds weight,improves the seeds weight and organ size in Brassica napus[J]. Plant Biotechnology Journal,2017,15(8):1024-1033.
[11]Peng Y,Chen L,Lu Y,et al. The ubiquitin receptors DA1,DAR1,and DAR2 redundantly regulate endoreduplication by modulating the stability of TCP14/15 in Arabidopsis[J]. The Plant Cell,2015,27(3):649-662.
[12]夏 天,李 杰,李胜军,等. DA1和DA2协同调控植物种子和器官大小[C]//植物分子生物学与现代农业——全国植物生物学研讨会论文摘要集,2010.
[13]Liu Y,Wang F,Zhang H,et al. Functional characterization of the Arabidopsis ubiquitin-specific protease gene family reveals specific role and redundancy of individual members in development[J]. The Plant Journal:for Cell and Molecular Biology,2008,55(5):844-856.
[14]Wang Z,Li N,Jiang S,et al. SCF(SAP) controls organ size by targeting PPD proteins for degradation in Arabidopsis thaliana[J]. Nature Communications,2016,7:11192.
[15]Li N,Liu Z,Wang Z,et al. STERILE APETALA modulates the stability of a repressor protein complex to control organ size in Arabidopsis thaliana[J]. PLoS Genetics,2018,14(2):e1007218.
[16]Wang A,Garcia D,Zhang H,et al. The VQ motif protein IKU1 regulates endosperm growth and seed size in Arabidopsis[J]. The Plant Journal:for Cell and Molecular Biology,2010,63(4):670-679.
[17]Garcia D,Saingery V,Chambrier P,et al. Arabidopsis haiku mutants reveal new controls of seed size by endosperm[J]. Plant Physiology,2003,131(4):1661-1670.
[18]Luo M,Dennis E S,Berger F,et al. MINISEED3 (MINI3),a WRKY family gene,and HAIKU2 (IKU2),a leucine-rich repeat (LRR) KINASE gene,are regulators of seed size in Arabidopsis[J]. Proceedings of the National Academy of Sciences of the United States of America,2005,102(48):17531-17536.
[19]Zhou Y,Zhang X,Kang X,et al. SHORT HYPOCOTYL UNDER BLUE1 associates with MINISEED3 and HAIKU2 promoters in vivo to regulate Arabidopsis seed development[J]. The Plant Cell,2009,21(1):106-117.
[20]Li J,Nie X,Tan J L,et al. Integration of epigenetic and genetic controls of seed size by cytokinin in Arabidopsis[J]. Proceedings of the National Academy of Sciences of the United States of America,2013,110(38):15479-15484.
[21]Jameson P E,Song J. Cytokinin:a key driver of seed yield[J]. Journal of Experimental Botany,2016,67(3):593-606.
[22]Riefler M,Novak O,Strnad M,et al. Arabidopsis cytokinin receptor mutants reveal functions in shoot growth,leaf senescence,seed size,germination,root development,and cytokinin metabolism[J]. The Plant Cell,2006,18(1):40-54.
[23]Kumar M N,Verslues P E. Stress physiology functions of the Arabidopsis histidine kinase cytokinin receptors[J]. Physiologia Plantarum,2015,154(3):369-380.
[24]Ulmasov T,Hagen G,Guilfoyle T J. ARF1,a transcription factor that binds to auxin response elements[J]. Science,1997,276(5320):1865-1868.
[25]Hagen G,Guilfoyle T. Auxin-responsive gene expression:genes,promoters and regulatory factors[J]. Plant Molecular Biology,2002,49(3/4):373-385.
[26]Ulmasov T,Hagen G,Guilfoyle T J. Activation and repression of transcription by auxin-response factors[J]. Proceedings of the National Academy of Sciences of the United States of America,1999,96(10):5844-5849.
[27]Okushima Y,Mitina I,Quach H L,et al. AUXIN RESPONSE FACTOR 2 (ARF2):a pleiotropic developmental regulator[J]. The Plant Journal,2005,43(1):29-46.
[28]Sun Y W,Wang C M,Wang N,et al. Manipulation of Auxin Response Factor 19 affects seed size in the woody perennial Jatropha curcas[J]. Scientific Reports,2017,7:40844.
[29]Ye J,Liu P,Zhu C,et al. Identification of candidate genes JcARF19 and JcIAA9 associated with seed size traits in Jatropha[J]. Functional & Integrative Genomics,2014,14(4):757-766.
[30]Jiang W B,Huang H Y,Hu Y W,et al. Brassinosteroid regulates seed size and shape in Arabidopsis[J]. Plant Physiology,2013,162(4):1965-1977.
[31]Zhu X,Liang W,Cui X,et al. Brassinosteroids promote development of rice pollen grains and seeds by triggering expression of Carbon Starved Anther,a MYB domain protein[J]. The Plant Journal,2015,82(4):570-581.
[32]Liu J F,Chen J,Zheng X M,et al. GW5 acts in the brassinosteroid signalling pathway to regulate grain width and weight in rice[J]. Nature Plants,2017,3:17043.
[33]Che R H,Tong H N,Shi B H,et al. Control of grain size and rice yield by GL2-mediated brassinosteroid responses[J]. Nature Plants,2015,2(1):15195.
[34]Xu R,Yu H,Wang J,et al. A mitogen-activated protein kinase phosphatase influences grain size and weight in rice[J]. The Plant Journal,2018,95(6):937-946.
[35]Duan P,Rao Y,Zeng D,et al. SMALL GRAIN 1,which encodes a mitogen-activated protein kinase kinase 4,influences grain size in rice[J]. The Plant Journal,2014,77(4):547-557.
[36]Liu S,Hua L,Dong S,et al. OsMAPK6,a mitogen-activated protein kinase,influences rice grain size and biomass production[J]. The Plant Journal,2015,84(4):672-681.
[37]Wang T,Zou T,He Z,et al. GRAIN LENGTH AND AWN 1 negatively regulates grain size in rice[J]. Journal of Integrative Plant Biology,2019,61(10):1036-1042.
[38]Xu R,Duan P,Yu H,et al. Control of grain size and weight by the OsMKKK10-OsMKK4-OsMAPK6 signaling pathway in rice[J]. Molecular Plant,2018,11(6):860-873.
[39]Matsuta S,Nishiyama A,Chaya G,et al. Characterization of heterotrimeric G protein γ4 subunit in rice[J]. International Journal of Molecular Sciences,2018,19(11):3596.
[40]Sun S,Wang L,Mao H,et al. A G-protein pathway determines grain size in rice[J]. Nature Communications,2018,9(1):851.
[41]Fujisawa Y,Kato T,Ohki S,et al. Suppression of the heterotrimeric G protein causes abnormal morphology,including dwarfism,in rice[J]. Proceedings of the National Academy of Sciences of the United States of America,1999,96(13):7575-7580.
[42]Oki K,Inaba N,Kitagawa K,et al. Function of the alpha subunit of rice heterotrimeric G protein in brassinosteroid signaling[J]. Plant & Cell Physiology,2009,50(1):161-172.
[43]Li S,Liu Y,Zheng L,et al. The plant-specific G protein γ subunit AGG3 influences organ size and shape in Arabidopsis thaliana[J]. The New Phytologist,2012,194(3):690-703.
[44]Roy Choudhury S,Riesselman A J,Pandey S. Constitutive or seed-specific overexpression of Arabidopsis G-protein γ subunit 3 (AGG3) results in increased seed and oil production and improved stress tolerance in Camelina sativa[J]. Plant Biotechnology Journal,2014,12(1):49-59.
[45]Li S,Liu W,Zhang X,et al. Roles of the Arabidopsis G protein γ subunit AGG3 and its rice homologs GS3 and DEP1 in seed and organ size control[J]. Plant Signaling & Behavior,2012,7(10):1357-1359.
[46]Li X,Tao Q,Miao J,et al. Evaluation of differential qPE9-1/DEP1 protein domains in rice grain length and weight variation[J]. Rice,2019,12(1):5.
[47]Liu Q,Han R,Wu K,et al. G-protein βγ subunits determine grain size through interaction with MADS-domain transcription factors in rice[J]. Nature Communications,2018,9:852.
[48]Wendt T,Holme I,Dockter C,et al. HvDep1 is a positive regulator of culm elongation and grain size in barley and impacts yield in an Environment-Dependent manner[J]. PLoS One,2016,11(12):e0168924.
[49]Ge L,Yu J,Wang H,et al. Increasing seed size and quality by manipulating BIG SEEDS1 in legume species[J]. Proceedings of the National Academy of Sciences of the United States of America,2016,113(44):12414-12419.
[50]Wang S,Wu K,Yuan Q,et al. Control of grain size,shape and quality by OsSPL16 in rice[J]. Nature Genetics,2012,44(8):950-954.
[51]Hu J,Wang Y,Fang Y,et al. A rare allele of GS2 enhances grain size and grain yield in rice[J]. Molecular Plant,2015,8(10):1455-1465.
[52]Duan P,Ni S,Wang J,et al. Regulation of OsGRF4 by OsmiR396 controls grain size and yield in rice[J]. Nature Plants,2015,2:15203.
[53]He Z S,Zeng J,Ren Y,et al. OsGIF1 positively regulates the sizes of stems,leaves,and grains in rice[J]. Frontiers in Plant Science,2017,8:1730.
[54]Li S,Gao F,Xie K,et al. The OsmiR396c-OsGRF4-OsGIF1 regulatory module determines grain size and yield in rice[J]. Plant Biotechnology Journal,2016,14(11):2134-2146.
[55]Zhang Y,Du L,Xu R,et al. Transcription factors SOD7/NGAL2 and DPA4/NGAL3 act redundantly to regulate seed size by directly repressing KLU expression in Arabidopsis thaliana[J]. The Plant Cell,2015,27(3):620-632.
[56]張雪晶,江文波,庞永珍. 植物种子大小调控机制的研究进展[J]. 植物生理学报,2016,52(7):998-1010.郑 林,祝令伟,郭学军,等. 沙门氏菌主要流行血清型耐药性的研究进展[J]. 江苏农业科学,2020,48(6):8-12.
