堆型艾美耳球虫体外培养细胞的筛选及其发育研究
2020-05-18王黎霞张建军
王黎霞,黄 芳,张建军,安 健
(1.北京农业职业学院牧医系;北京 102442;2.北京农学院动物科学技术学院,北京 102206)
球虫体外培养对于研究球虫发育、球虫病防治和球虫与宿主的关系具有重大的意义和作用,首先能够更清楚研究球虫细胞内的发育状况[1];其次可分离到较纯净的抗原,为疫苗的研制打下基础;第三选育弱毒株,为制作弱毒活疫苗提供条件;第四进行新型抗球虫药或是抗球虫药物新剂型研究提供条件[2-3];第五能够更直观检测球虫与宿主细胞之间的相互作用[4]。更重要的是在科学研究中,体外培养可以减少动物试验,是动物福利实现的重要途径。
球虫体外培养中,不同细胞系对球虫的生长和发育有很大影响。牛肾传代细胞(Madin-Darby bovine kidney,MDBK)是培养E.tenella的良好细胞系[5],但不适合布氏艾美耳球虫的培养[6],小隐孢子虫在其中也无法发育[7]。幼地鼠肾细胞(baby hamster kidney,BHK)被用于培养艾美耳属球虫,且认为是优于鸡肾原代细胞和火鸡肾原代细胞[8]。Tierney认为BHK、MDBK、猪肾传代细胞(porcine kidney,PK)-13较适宜培养E.tenella[9]。MDBK和肝母细胞瘤细胞(hepatoblastoma,Hep)-2较适合培养刚地弓形虫,非洲绿猴肾传代细胞(African green monkey kidney,VERO)、BHK、PK-13对于弓形虫的体外培养不是很好[10]。人结肠癌细胞(human colon cancer cell,HCT)-8被认为最适合体外培养小隐孢子虫[11],有人比较小隐孢子虫入侵狗肾传代细胞(Madin-Darby canine kidney,MDCK)、小鼠星型胶质细胞(Mouse astrocytes,MA)-104、Hep-2和VERO,发现MDCK最易于感染,在MA-104细胞中小隐孢子虫无法进行有性生殖,VERO细胞入侵最少而且无法形成滋养体[12]。也有研究发现兔软骨细胞(Rabbit chondrocytes,VELI)体外培养小隐孢子虫得到卵囊,再次感染小鼠时的毒力更强,说明VELI细胞更适于体外培养小隐孢子虫[13]。弓形虫在宫颈癌细胞(Henrietta Lacks cervical cancer cell,Hela)中培养可以产生大量的速殖子[14],可见,细胞系对于体外培养寄生虫的影响很大,不同的胞内寄生虫对不同的细胞系的入侵和发育差别很大,所以在体外培养寄生虫时应该广泛选择细胞系。
E.acervulina体外培养过程中,有报道认为早熟减毒株裂殖子在鸡胚成纤维原代细胞(chicken embryo fibroblast,CEF)、鸡肾原代细胞(chicken kidney,CK)中不能发育,而在鸡胚小肠上皮原代细胞可以发育到卵囊阶段[15]。有研究认为E.acervulina入侵鸡胚小肠上皮48 h形成第一代裂殖子,60 h可见大小配子体,72 h可以见到合子[16]。本试验比较MDBK、VERO、PK-15、CEF、CK 5种不同细胞在37 ℃和41 ℃下,子孢子的入侵和发育的情况,以期筛选出适宜E.acervulina体外培养的细胞。
1 材料和方法
1.1 试验材料
虫株:E.acervulina孢子化卵囊,由河北农业大学秦建华馈赠,北京农学院动物科学技术学院病理实验室繁殖并保存。
细胞系:MDBK,由中国农业大学索勋馈赠;VERO、PK-15,由中国农业大学吴清民馈赠。
重要试剂: 改良伊格尔培养基(Dulbecco's modified eagle medium,DMEM)、新生牛血清为Gibico公司产品,青霉素、1∶250胰酶为Orien公司产品,硫酸链霉素为Topbio公司产品,苏木精、伊红为Amersco公司产品。
1.2 方 法
1.2.1 试验材料的纯化和培养E.acervulina子孢子的纯化参考黄芳[17],CEF培养参考陈淑亚[18]、CK培养参考姜逸[19]、传代细胞系培养参考曾华书[20]。
1.2.2 不同细胞系、不同温度下的E.acervulina子孢子入侵率的比较 用胰酶37 ℃,5%CO2消化3 min,进行细胞计数,调整细胞密度为5×105个/mL。将盖玻片加入6孔板中,每孔加入细胞悬液2 mL,在37 ℃、5%CO2和41 ℃、5%CO2分别培养2 h,显微镜下观察细胞达到融合。将纯化好的子孢子计数,每孔接入105个,再培养24 h,HE染色观察,每个样品观察3个视野,算出平均数,计算入侵率。入侵率=一个视野下被子孢子侵入细胞数/一个视野下细胞数×100%。
1.2.3 不同细胞系、不同温度对入侵时间的影响 按1.2.2方法在37 ℃,5%CO2和41 ℃,5%CO2培养,每隔0.5 h进行HE染色,共观察3 h,观察入侵时间。
1.2.4E.acervulina在MDBK中的发育 按1.2.2方法37 ℃,5%CO2培养,每隔12 h更换1次无血清培养基,每隔2 h进行高碘酸-shiff染色,观察球虫发育情况。
2 结 果
2.1 不同细胞系、不同温度对E. acervulina入侵时间的影响
温度明显可以影响子孢子的入侵时间,子孢子在41 ℃比37 ℃能够较快入侵细胞,37 ℃时子孢子入侵不同细胞系的时间一致,41 ℃时PK-15细胞入侵所需时间较长(表1)。
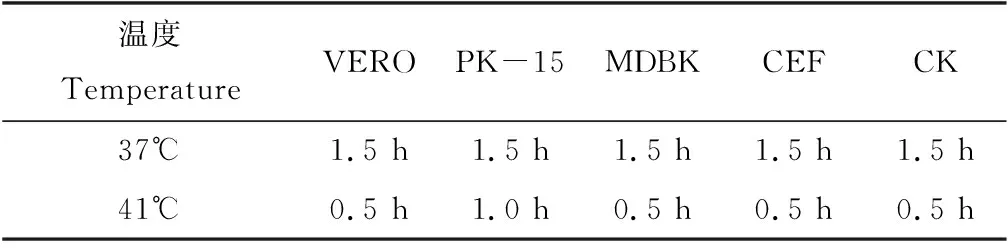
表1 不同细胞、不同温度下E. acervulina子孢子入侵时间Tab.1 The time of E. acervulina sporozoite invasion five cell at 37 ℃ and 41 ℃
2.2 不同细胞系和温度对E. acervulina入侵率的影响
在37 ℃时,MDBK和PK-15的入侵率显著高于VERO、CEF、CK(P<0.05),MDBK的入侵率高于PK-15,但二者差异不显著(P>0.05),VERO、CEF、CK的入侵率差异不显著(P>0.05),见图1。
41 ℃时,MDBK入侵率最高,但MDBK、PK-15、VERO、CEF的入侵率差异不显著(P>0.05),CK的入侵率显著低于其他4种细胞(P<0.05),见图1。
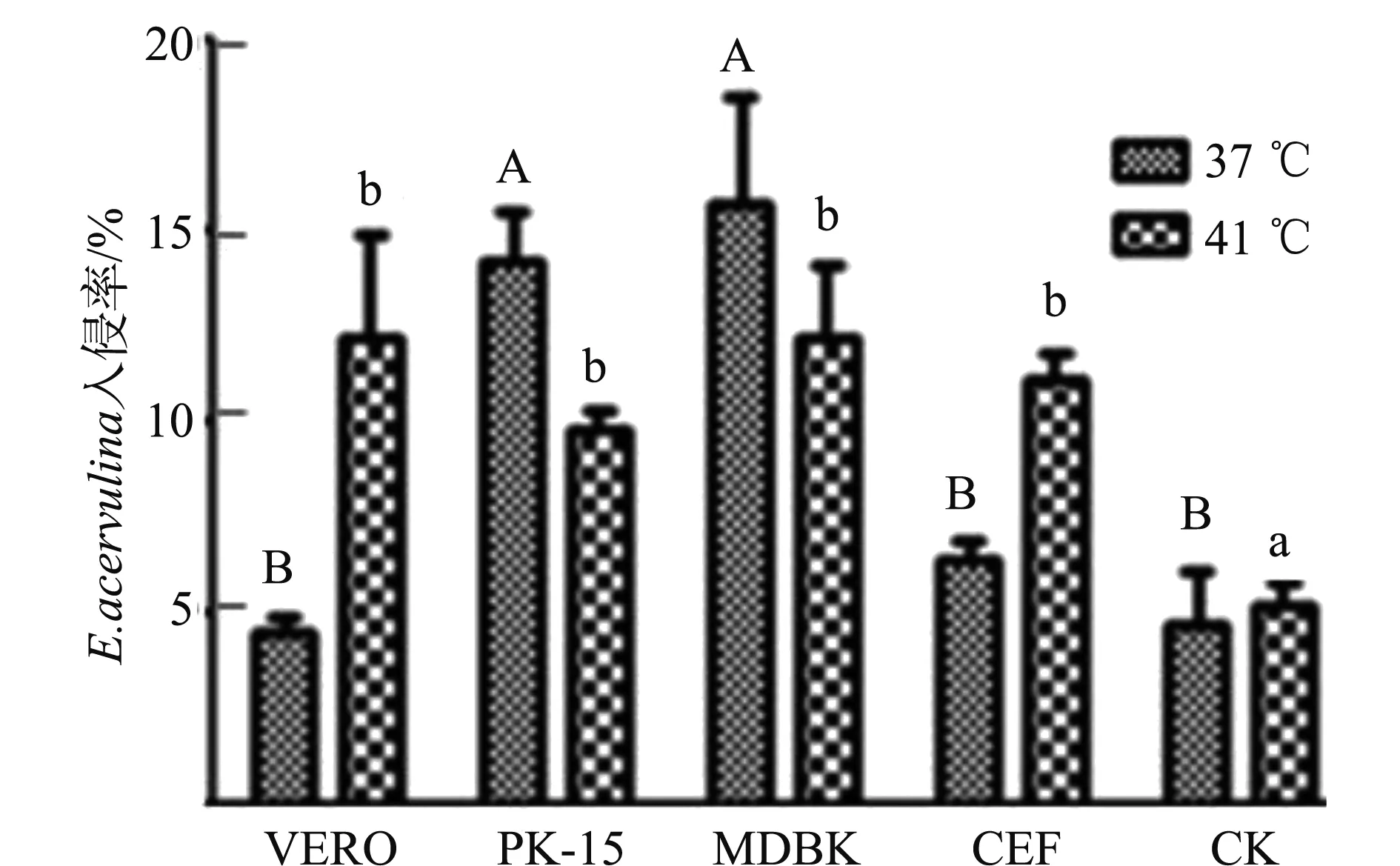
注:A、B、a、b字母不同表示差异显著(P<0.05),字母相同表示差异不显著(P>0.05)。Note: Different letter is significant difference(P<0.05),same letter is not significant difference(P>0.05).图1 不同温度、不同细胞系E. acervulina入侵率Fig.1 The invasion rate at different temperature and different cell lines
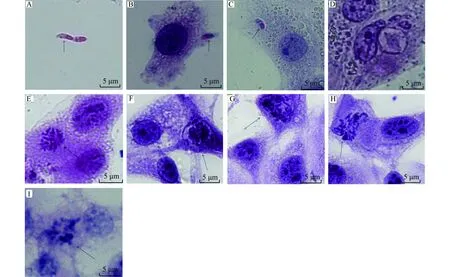
注:A,入侵前的子孢子(↑);B,共培养1 h附着于细胞表面的子孢子(↑);C,2 h后细胞入侵成功(↑);D,24 h后的子孢子长大,发育为滋养体(↑);E,52 h的第一代裂殖体(↑);F,56 h第一代裂殖体发育成熟(↑);G,H,60 h第一代裂殖子正在释放出细胞(↑);I,72 h形成第二代裂殖体(↑)。Note: A, The sporozotie before invasion(↑); B, the sporozoite adhere to the cell after co-culture 1 h(↑); C, the sporozoite entered the cell after co-culture 2 h(↑); D, the trophozoite within parasitophorous vacuole after co-culture 24 h(↑); E, the developed schizont after co-culture 52 h(↑); F,the developed schizont after co-culture 56 h(↑); G, H, the merozoite released from cells after co-culture 60 h(↑); I, the second generation schizont formed after co-culture 72 h(↑).图2 E. acervulina MDBK内发育Fig.2 Development of E. acervulina in MDBK
比较温度对入侵率的影响,PK-15和MDBK 37 ℃的入侵率显著高于41 ℃的入侵率, VERO、CEF、CK 41 ℃的入侵率显著高于37 ℃的入侵率,但总体比较MDBK的入侵率无论是在41 ℃还是在37 ℃下培养入侵率都较高,见图1。
2.3 E. acervulina 在MDBK中发育情况
染色后显微镜检查发现,共培养1 h子孢子附着于细胞表面,2 h后子孢子入侵MDBK中,形成带虫空泡。24 h后子孢子转化为早期第一代裂殖体,52 h开始形成第一代裂殖体,56 h形成成熟的第一代裂殖体,60 h第一代裂殖子释放出细胞,72 h形成完整的第二代裂殖体,证明E.acervulina利用MDBK在37 ℃培养可以进行胞内繁殖,见图2。
3 讨 论
细胞对球虫体外培养的影响很大,MDBK是来自牛的细胞系,适宜大部分艾美耳属寄生虫体外培养,但不适合布氏艾美耳球虫和肉孢子虫的培养[6]。HCT-8和MDCK适宜小隐孢子虫的培养[11],原代牛脐静脉内皮细胞适宜牛艾美耳球虫和祖氏艾美耳球虫的培养[21],因此,细胞对于体外培养寄生虫的影响十分明显,为了筛选出适宜E.acervulina体外培养的细胞,本试验选择MDBK、PK-15、VERO、CEF和CK 5种细胞进行体外培养对比试验。
41 ℃是鸡的体温,球虫生活在鸡的肠道,41 ℃比较适宜子孢子生长,对比41 ℃和37 ℃对入侵的影响。在三个传代细胞系中,41 ℃都不利于细胞生长,温度改变后细胞基本处于维持状态无法进行繁殖,不适宜体外培养E.acervulina,对于原代细胞41 ℃时只有入侵时间缩短,入侵率并没有优势。原代细胞传代中细胞发育不稳定,一般认为成纤维细胞可以传代20次左右,但是在试验过程中无法保证每次传代的生长效果完全一致,对于下一步的研究有一定的影响。
Naciri-Bontemps用3日龄鸡肾原代细胞体外培养E.acervulina时,在EHT培养基培养44 h可见第一代裂殖体。感染54 h第一代裂殖子释放出细胞。68 h,其入侵周围的细胞并发育成第二代裂殖体。72~96 h,可以观察到第三代和第四代裂殖子。第四代裂殖子可以发育成为配子体,当把收集的配子体感染给鸡45 h可以收获卵囊[22]。E.acervulina子孢子和细胞共培养2 h才侵入细胞,在42 h第一代裂殖体发育完全,56 h才从细胞中释放出来。72 h可见第二代裂殖子发育成熟。这可能与培养基中血清含量有关。有研究证明当培养基成分为90% HBSS、5%乳清蛋白水解物和5%胎牛血清比用95% M199和5%胎牛血清,E.tenella入侵更多[23]。有研究曾比较鸡血清,小牛血清,兔血清和猪血清对鸡原代细胞的生长、子孢子入侵能力以及对E.tenella的第一代和第二代裂殖体的作用效果的影响,发现,鸡血清组子孢子入侵能力最强,胎牛血清组第二代裂殖体比鸡血清多,但鸡血清组的卵囊比胎牛血清多[9]。有研究证明,在无血清培养基中,人隐孢子虫和小隐孢子虫可以发育,并能完成生活史[11]。
MDBK培养E.acervulina可以形成第二代裂殖体,而VERO、PK-15只能发育到第一代裂殖体阶段,CEF和CK中E.acervulina有第一代裂殖子的释放,但未再次入侵,并且CEF和CK不易传代,5种细胞中,MDBK最适宜E.acervulina培养;与MDBK共培养1 h子孢子附着于细胞表面,2 h子孢子入侵,子孢子入侵24 h形成第一代滋养体,52~56 h形成第一代成熟裂殖体,60 h第一代裂殖子释放出细胞,72 h形成第二代裂殖体,在VERO、MDBK、PK-15、CEF及CK这5种细胞中,MDBK是培养E.acervulina的最适宜细胞。
