胰高血糖素样肽-1及其类似物对细胞氧化应激的研究进展
2020-05-12吴灵灵孙李丹
吴灵灵,孙李丹
(嘉兴学院,嘉兴314000)
胰高血糖素样肽-1(GLP-1)是一种主要由远端小肠L细胞响应营养摄入而分泌产生的肠促胰岛素[1]。GLP-1与受体结合后,可以通过刺激胰岛素分泌、抑制胰高血糖素分泌、抑制胃排空和减少食物摄入等途径影响血糖水平[2]。GLP-1对血糖具有较好的调节作用,不会诱发低血糖,因此在糖尿病临床治疗过程中得到了广泛的应用。GLP-1除了具有降血糖作用外,在肾、肺、心脏、下丘脑、内皮细胞、神经元、星形细胞、小胶质细胞等多种细胞和组织中具有抗炎、抗氧化、神经生成和血管保护作用。GLP-1和GLP-1受体(GLP-1R)激动剂也影响多种细胞通路,包括抑制炎症和细胞凋亡、保护机体免受氧化应激等。本文将着重总结GLP-1在抗氧化应激方面的保护作用。
1 GLP-1
GLP-1是一种胰高血糖素原的产物,由胰腺α细胞以及小肠下层和结肠内分泌细胞L细胞分泌产生。体内 GLP-1的主要形式为 GLP-1(7-36)-NH2(约占80%)和 GLP-1(7-37)-OH(约占20%),两者具有相同的生理功能。GLP-1具有以下作用:①通过G蛋白偶联受体,使细胞内的cAMP增加,引起胞吐作用,从而促进胰岛素分泌,也可通过正向调节胰岛素的基因表达,增加胰岛素的生物合成;②促进β细胞的增殖而增加β细胞积聚,也增加胰管上皮细胞的祖细胞分化,减少β细胞凋亡;③延缓胃排空,并有剂量依赖性[2],机制可能由迷走神经介导;④通过调整胰岛素和胰高血糖素分泌影响葡萄糖的代谢[1],可能是肠内L细胞分泌的GLP-1直接进入了门脉循环,直接调节肝脏摄取葡萄糖的结果;⑤通过作用于下丘脑的GLP-1R产生短暂的饱胀感,减退食欲、降低热量摄入[2]。
GLP-1主要通过GLP-1R而发挥各项生理作用。人体机体中GLP-1R的分布范围十分广泛,不仅分布于常见的胰腺胰岛α细胞和β细胞,在人体心、肝、肠道、胃、视网膜色素上皮细胞、下丘脑、迷走神经神经节以及中枢系统中也有一定分布。GLP-1R属于B族G蛋白偶联受体,由463个氨基酸组成。GLP-1R的N端位于细胞膜外,具有3个保守的二硫键,可与配体结合,C端位于细胞膜内,可与偶联的G蛋白作用来传导信号。目前临床上可用于治疗成年患者2型糖尿病的GLP-1R激动剂主要包括塞马鲁肽、艾塞那肽、利拉鲁肽、利西拉来、长效艾塞那肽、阿必鲁肽以及度拉鲁肽等。
1.1 GLP-1的合成与代谢
胰高血糖素原基因在小肠和大肠L细胞、中枢神经系统以及胰腺α细胞中表达,不同多肽通过胰高血糖素原特异性翻译后加工产生。在胰腺α细胞中,激素原转化酶2加工胰高血糖素原,产生胰高血糖素(主要产物)、胰高血糖素相关多肽、插入肽-1和主要的胰高血糖素原片段。在肠和脑中,激素原转化酶处理胰高血糖素原,产生GLP-1、GLP-2、胃泌酸调节素、肠高血糖素和插入肽-2,其中GLP-1是由胰高血糖素原产生的主要多肽。
GLP-1的两种活性形式GLP-1(7-37)和GLP-1(7-36)酰胺可随着体内碳水化合物、脂肪、蛋白质和膳食纤维等营养素的吸收而迅速增加。循环系统中的GLP-1被二肽酶快速降解,产生GLP-1(9-37)和 GLP-1(9-36)酰胺[3]。这些代谢物可以进一步裂解生成其他代谢片段,已有文献报道GLP-1代谢物具有许多有益的作用,包括抑制肝脏葡萄糖的产生、心脏和神经保护作用、减少血管内的氧化应激、抑制胰腺细胞凋亡和增殖作用等[3]。
1.2 GLP-1对胰腺的影响
GLP-1可促进胰腺β细胞释放胰岛素并具有葡萄糖依赖性,胰岛细胞加入1或10 nmol/L葡萄糖与GLP-1温孵12或24 h后,胰岛素释放量与细胞内胰岛素含量均明显高于对照组[4]。GLP-1诱导的胰岛素分泌主要由激活cAMP-PKA和cAMP(Epac)2信号通路的交换蛋白调节[5]。此外,GLP-1还能增强mRNA稳定性、促进生物合成和胰岛素原基因表达。胰腺胰高血糖素原基因的表达可因禁食和低血糖而上调,并受胰岛素抑制。激活蛋白激酶 C(PKC)信号通路、cAMP/蛋白激酶 A(PKA)通路和Ca2+信号通路均可增加胰岛胰高血糖素原腺胰高血糖素原基因转录。
研究表明,GLP-1可促进β细胞增殖和新生,并抑制 β细胞凋亡。PKB、PKC、PI3K、c-Src和MAPK信号通路介导GLP-1诱导的胰腺β细胞增殖。转录因子7-样2和Wnt/β-连环蛋白信号通路也参与GLP-1诱导的胰岛素分泌和β细胞增殖[6]。与野生型小鼠相比,GLP-1R缺陷型小鼠切除70%胰腺后,β细胞数量的增加明显减少,表明GLP-1在β细胞再生中的重要作用[6]。GLP-1还可作用于细胞表面的 GLP-1R,通过 PI3K/AKT/FOX01信号通路增强PDX-1的反应性,从而维持β细胞形态功能并促进非β细胞向β细胞转化。多项研究报道了GLP-1和GLP-1R激动剂对β细胞抵抗各种毒性刺激物(包括葡萄糖、脂肪酸、细胞因子和ROS)的预防作用[7]。GLP-1的抗凋亡作用是通过增加抗凋亡蛋白Bcl-2和Bcl-xL水平并激活cAMP和PI3K以及PKB和EGFR-PI3K信号通路来降低半胱天冬酶-3活性来介导产生[6]。
1.3 GLP-1的胰腺外效应
除了对胰腺的作用外,GLP-1还可降低肝脏中的葡萄糖产量并增加脂肪组织和肌肉中的葡萄糖摄取。
GLP-1的生物学效应通过与其受体结合而选择性介导,GLP-1R是特异性跨膜受体鸟嘌呤核苷酸联合蛋白(G蛋白)偶联受体(GPCR),遍布于胰岛、脑、中枢、肾和肠胃。GLP-1及其抗蛋白酶类似物可穿过血-脑脊液屏障,保护记忆形成或活动,保护突触和突触功能,减弱细胞凋亡,保护神经元免受氧化应激的影响,减少小鼠斑块形成和退行性疾病小鼠的炎症反应[如阿尔茨海默病(AD)和帕金森病(PD)等[8],表明GLP-1可能对神经退行性疾病的治疗具有一定作用。另外,GLP-1可影响肾功能并对心脏具有一定的保护作用。GLP-1通过直接或间接途径作用于胃肠道,抑制胃肠道蠕动和胃液分泌,延长胃排空时间,延缓营养物质吸收。同时,它作用于下丘脑,增加饱腹感,减少食物摄入量,减少脂肪沉积并减轻体重。此外,GLP-1还可以抑制内皮蛋白C受体(EPCR)脱落,防止血栓形成。GLP-1还通过减少炎性细胞因子的产生和免疫细胞的浸润,对胰岛和其他组织[如脂肪组织、肝脏、大脑、内皮细胞、肾脏和皮肤]具有抗炎作用[6]。因此,GLP-1具有治疗非酒精性脂肪性肝炎、神经退行性疾病、动脉粥样硬化、糖尿病肾病和银屑病等疾病的潜力。
2 GLP-1在糖尿病中的抗氧化作用
糖尿病及其并发症的产生与氧化应激增加密切相关。糖尿病经常伴随着自由基产物的增加和抗氧化防御系统的损坏。GLP-1或其受体激动剂的治疗可显著降低多种诱导氧化应激标记物(超氧化物歧化酶、谷胱甘肽还原酶、过氧化氢酶、谷胱甘肽过氧化物酶、谷胱甘肽、脂质过氧化和非酶糖化蛋白)的水平。GLP-1降低糖尿病氧化应激的机制可通过cAMP、PI3K和PKC途径介导以及Nrf-2的激活[6]。
Nrf2敲除的小鼠胰岛素瘤细胞(MIN6)或Nrf2敲除小鼠分离的胰岛细胞易受砷诱导产生细胞损伤[9],表明Nrf2信号通路对砷诱导的细胞毒性具有保护作用。激活Nrf2可改善胰腺移植后的胰岛产量和胰岛功能[10],表明Nrf2在保护胰腺β细胞免受氧化应激方面发挥重要作用。此外,激活Nrf2可提高糖尿病对胰岛素的敏感性,改善小鼠的糖尿病和肥胖,增加骨骼肌中的氧气消耗和葡萄糖摄取。以上结果表明,GLP-1诱导的Nrf2激活有利于预防和治疗糖尿病。
2.1 体外研究
多项体外研究证实了GLP-1对氧化应激具有一定保护作用。糖化血清(GS)培养HIT-T15细胞5 d后,氧化应激增加,胰岛素原含量减少,PDX-1和MafA结合 DNA的能力减弱,加入10 nmol/L GLP-1后可逆转GS诱导的细胞损伤,表明GLP-1可保护细胞免受氧化应激影响[11]。GLP-1还可降低INS-1细胞中叔丁基过氧化物诱导的氧化应激,恢复细胞内谷胱甘肽水平及其相关过氧化物酶和还原酶活性[12]。INS-1细胞与细胞因子温孵后产生高浓度ROS而增加细胞凋亡,而与GLP-1R激动剂艾塞那肽温孵后完全阻止了这种作用[13]。自发糖尿病大鼠经艾塞那肽治疗后,葡萄糖诱导的ROS产生减少,Src磷酸化水平显著降低。当用Epac特异性cAMP类似物(9CPT-2Me-cAMP)处理时,Src磷酸化和ROS产生减少,表明Epac介导的Src失活与艾塞那肽的抗氧化作用相关,与PKA或MAPK/ERK信号通路无关[6]。GLP-1(28-36)酰胺(100 nmol/L)也可抑制ROS形成并恢复细胞ATP水平[14]。Kim等[15]报道艾塞那肽可减少棕榈酸酯或H2O2诱导的ROS产生并恢复INS-1细胞中的谷胱甘肽水平和胰岛素分泌,表明PKCδ介导的Nrf2激活增加了抗氧化基因表达,改善氧化应激状态下β细胞功能。这些体外数据表明,GLP-1通过降低ROS、激活Nrf2和诱导抗氧化酶来增加抗氧化能力而发挥抗氧化作用。
2.2 体内研究
针对糖尿病患者的临床研究表明,GLP-1短期和长期治疗可降低高血糖和氧化应激。2型糖尿病患者在低血糖或高血糖症期间给予GLP-1 0.4 pmol/(kg·min),治疗 2 h可降低血浆中的 8-异前列腺素F2α(8-iso-PGF2α)和硝基酪氨酸(氧化应激标记物)水平[6]。艾塞那肽治疗 1年(10μg/d)可降低2型糖尿病患者的餐后血糖和血脂,这些作用与降低丙二醛(脂质过氧化标记)和氧化低密度脂蛋白有关[16]。然而,在另一项研究中观察到了不同的结果,利拉鲁肽治疗8个月(0.74 mg/d)不影响血清丙二醛水平[17]。这种差异可能是由于所用的利拉鲁肽剂量较低。在一项为期2个月的前瞻性试验中,2型糖尿病患者接受利拉鲁肽(1.2 mg/d)治疗后,血清脂质过氧化物、血红素加氧酶1和糖化血红蛋白降低[18]。
GLP-1在1型和2型糖尿病动物模型中的部分作用可归因于其抗氧化活性。重组人GLP-124 nmol/(kg·d)可降低糖尿病小鼠链脲佐菌素(STZ)诱导的高血糖,增强胰腺超氧化物歧化酶和谷胱甘肽过氧化物酶活性[19]。STZ诱导的糖尿病大鼠连续10周给予艾塞那肽1μg/(kg·d)后,血糖水平降低和胰腺胰岛素释放增加。此外,胰腺中过氧化氢酶和谷胱甘肽还原酶阳性细胞数量增加[6]。Shimoda等[20]发现在糖尿病 db/db小鼠接受 GLP-1类似物利拉鲁肽400μg/(kg·d)治疗2周有利于保护胰腺β细胞,过氧化氢酶、谷胱甘肽过氧化物酶等抗氧化应激相关基因在胰腺β细胞中的表达显著增加。
肥胖ob/ob小鼠使用艾塞那肽10或20μg/(kg·d),治疗60 d后,血糖和肝脂肪变性显著降低,胰岛素敏感性提高,肝脏脂肪变性明显改善,肝组织中作为氧化应激标志物的硫代巴比妥反应物水平显著降低[21]。与单用艾塞那肽或奥美拉唑相比,STZ诱导的糖尿病小鼠使用奥美拉唑30 mg/(kg·d)和艾塞那肽8μg/(kg·d)联合治疗4周后,血糖水平明显改善,这可能与肝脏脂质过氧化减少和 Nrf-2表达增加有关[22]。
3 GLP-1在糖尿病并发症中的抗氧化作用
大量体外和体内研究表明,GLP-1对糖尿病并发症具有一定的保护作用。在2型糖尿病患者中,低GLP-1水平和高硝基酪氨酸(ROS-依赖性氧化应激标记物)水平与心脏重塑和心血管疾病的发展相关。在HL-1心肌细胞中,GLP-1(100和200 nmol/L)可降低棕榈酸诱导的细胞质和线粒体氧化应激,同时增加ATP合酶表达[6]。据报道,GLP-1及其类似物通过调节c-Jun N末端蛋白激酶信号通路、磷酸肌醇3-激酶(PI3K)通路和 ERK1/2通路来抑制心肌细胞凋亡。此外,GLP-1可恢复线粒体膜通透性和细胞色素C氧化酶活性,从而抑制线粒体中的氧化损伤。使用艾塞那肽作用于新生大鼠心肌细胞,H2O2诱导的ROS生成下降,过氧化氢酶、谷胱甘肽过氧化物酶-1和锰超氧化物歧化酶等抗氧化酶的合成增加,该作用主要依赖于GLP-1R介导的Epac途径[23]。艾塞那肽(1和200 nmol/L)可以改善H2O2和缺氧/复氧导致的心肌细胞(H9C2)细胞凋亡、线粒体功能障碍和ROS生成,该作用与PI3K/AKT信号通路的活化有关[6]。多项证据表明AKT磷酸化可激活PI3K/AKT信号通路,保护心肌免于MI/R后的细胞凋亡。
艾塞那肽24 nmol/(kg·d),治疗 40 d还可减轻高脂饮食诱导的糖尿病性心肌病,如心肌纤维化和脂肪变性。此外,艾塞那肽通过抑制ROS产生酶-烟酰胺腺嘌呤二核苷酸磷酸(NADPH)氧化酶4,升高抗氧化酶(超氧化物歧化酶-1和谷胱甘肽过氧化物酶)而改善心肌氧化应激[20]。此外,艾塞那肽(10μg/kg)治疗还可逆转STZ和高脂饮食诱导糖尿病大鼠的主动脉氧化损伤[24]。
糖尿病慢性肾病倾向性KK/Ta-Akita小鼠的蛋白尿和肾小球系膜扩张增加,肾小球超氧化物和肾NADPH氧化酶上调,使用西格列汀(一种DPP-4抑制剂,10 mg/d)连续治疗20周后,自发糖尿病大鼠肾中抗氧化应答基因(过氧化物还原酶和谷胱甘肽 S-转移酶)的表达降低且 miR-200a下调[25]。研究证明糖尿病小鼠 miR-200a通过醛糖还原酶依赖性机制来调节肾细胞中的Nrf2[26]。利拉鲁肽200μg/(kg·d)治疗4周可抑制肾病进展,并且该效应由 cAMP和 PKA水平升高介导[27]。Hendarto等[28]也报道用利拉鲁肽 0.6 mg/(kg·d)治疗STZ诱导的糖尿病大鼠,治疗4周可减少肾组织中的氧化应激标记物(尿:8-羟基-2-脱氧鸟苷,肾:二氢乙锭染色)并而改善糖尿病肾病。
4 GLP-1在衰老中的抗氧化作用
Denham Harman提出的自由基衰老理论假定衰老是自由基引起的有害影响累积导致。ROS诱导的细胞损伤对生物体寿命具有重要的影响。与该理论一致,衰老组织的线粒体ROS产生增加,mtDNA的8-氧代-dG含量也增加,氧化性DNA损伤的逐渐积累是导致衰老的重要促成因素。多项研究表明,GLP-1对细胞衰老具有保护作用。GLP-1(10 nmol/L)治疗可抑制 H2O2诱导的人脐静脉内皮细胞DNA损伤和细胞衰老,并使用维达列汀抑制 DPP-43 mg/(kg·d)可防止 Zucker糖尿病肥胖大鼠的血管衰老[6]。研究还发现cAMP/PKA依赖性途径可介导GLP-1对血管的抗衰老作用。
5 GLP-1在神经疾病中的抗氧化作用
GLP-1R激动剂可穿过血-脑脊液屏障,目前已有多项研究对GLP-1神经炎症、线粒体功能、神经元保护和中枢神经系统内细胞增殖的影响进行探究。脑缺血再灌注损伤模型小鼠经过艾塞那肽(10μg/kg)治疗后,细胞内cAMP水平增加并且可观察到GLP-1的神经保护作用[29]。在脑缺血的大鼠模型中,长效艾塞那肽负载的微球(NF)-κBp65和 p-AKT/内皮型一氧化氮合酶(p-AKT/p-eNOS)通路介导了神经保护作用[30]。与缺血对照组相比,利拉鲁肽(50μg/kg)预治疗14 d后,大鼠大脑中动脉闭塞诱导的脑缺血后梗死体积更小,神经功能缺损减少,脑组织中脂质过氧化标记物水平降低,包括谷胱甘肽和超氧化物歧化酶在内的抗氧化分子活性提高[6]。利拉鲁肽还可穿过血-脑脊液屏障,提高脑内cAMP水平,有利于神经退行性疾病的治疗。利拉鲁肽100μg/(kg·d)可减少缺血大鼠的梗死体积并改善运动和体感功能,表明利拉鲁肽可通过激活PI3K/AKT和MAPK通路发挥其对缺血诱导神经细胞凋亡的保护作用[31]。利拉鲁肽(500 nmol/L)还可通过降低原代神经元ROS、增加AKT和ERK磷酸化及降低p-38和c-Jun N端激酶的磷酸化来抑制凋亡[31]。利拉鲁肽(50 nmol/L)可通过cAMP-反应元件结合蛋白磷酸化来促进人体神经母细胞瘤细胞的增殖。此外,利拉鲁肽预治疗可降低H2O2或谷氨酸诱导的细胞凋亡并改善创伤性脑损伤小鼠行为。这些结果表明GLP-1可通过氧化应激相关保护作用来治疗神经疾病。
6 结 论
ROS和氧化应激促进了包括糖尿病、糖尿病并发症、神经疾病和肿瘤等多种慢性疾病的致病过程。体外和体内研究结果表明GLP-1和GLP-1R激动剂可降低ROS水平,提高抗氧化酶的表达并激活Nrf2。尽管上述作用属于GLP-1的葡萄糖非依赖性效应,但GLP-1在糖尿病及其并发症中的抗氧化作用也可能部分归因于其降低葡萄糖的作用。本文大部分结果来自体外和体内研究(图1),GLP-1抗氧化作用的详细机制需要进一步研究。
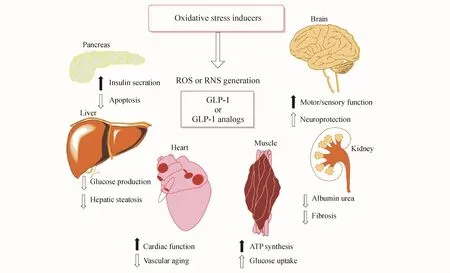
Figure 1 Antioxidant mechanisms of GLP-1 on various organs.GLP-1 receptor-mediated(GLT-1R)signaling blocks the production of reactive oxygen species(ROS)induced by various oxidative stressors and regulates physiological functions of various organs(black solid arrow,increase;black hollow,reduce)
