食欲素B增强外膝体纤维投射至视皮层第6b层锥体细胞的突触传递
2020-04-11黄梦瑶
童 昕,黄梦瑶,蒋 斌
(中山大学中山医学院广东省脑功能与脑疾病研究重点实验室,广东广州 510080)
1998 年日本人Yanigasawa 首次在大鼠下丘脑腹外侧区(lateral hypothalamic area,LHA)发现新的神经肽。这种神经肽主要作用是维持动物的觉醒状态,敲除该神经肽基因的小鼠表现出发作性睡病[1]。后来的研究发现这种神经肽参与机体能量代谢,但作用与瘦素(leptin)相反,因此命名为食欲素(Orexin,OX)[2]。分泌食欲素(OX)的神经细胞位于下丘脑腹外侧区,它的神经纤维广泛投射至皮层和皮层下组织。OX 可分为两个亚型,Orexin A和Orexin B,都可以与Orexin 的两种受体(OX1 和OX2)结合,但亲和力有所不同[3]。视皮层也接受来自下丘脑OX神经纤维的投射。在视觉信息的传导通路中,视觉信号经眼球视网膜传送到丘脑的外膝体,信号经处理后再传至视皮层,外膝体的纤维主要传至视皮层第4 层,少量传至到第2~3 层以及第5~6 层[4],而视皮层的第6 层也将来自视皮层第2~3 层的信号反馈传递给丘脑,从而形成反馈环路[5]。作为多形细胞层,L6b 神经元又按不同的形态,分为锥体细胞、多极细胞、倒置朝向细胞、水平朝向细胞和不定向细胞[6]。尽管前人报道了视皮层第6b 层(L6b)是视皮层6 个片层中唯一对OX有反应的亚层[7],但具体是哪种类型的神经元没有明确。本研究证明了Orexin B 可以直接兴奋大多数L6b 的锥体神经元,同时还发现Orexin B 通过增强L6b 锥体细胞突触后NMDAR 电流来增强外膝体(lateral geniculate nucleus,LGN)前馈投射到L6b锥体细胞的突触传递。该研究提供了Orexin B 参与调控皮层-丘脑之间神经环路功能的新证据,同时也为动物在清醒状态下,Orexin增强视皮层对视觉信息的敏感性提供了细胞学上的依据。
1 材料与方法
1.1 实验动物
本研究所涉及动物实验方案获得中山大学中山医动物伦理(2015)第004 号批准。实验选用P25~P45 的野生型C57 小鼠(SPF 级,购于中山大学动物实验中心),性别不限。小鼠饲养条件和处理方式严格遵守中山大学动物伦理的规定,动物房的温度控制在24 ℃左右,相对湿度在55%左右。光照时间12 h,黑暗12 h,模拟昼夜变化。
1.2 脑立体定位注射
选择P25~P30 左右的C57 小鼠,称重,按剂量1 mL/20 kg 体质量腹腔注射5 g/L 戊巴比妥钠溶液使之麻醉。根据小鼠大脑立体定位图谱定位LGN的坐标,将Bregma定为零点(0,0,0),Bregma向人字点2.3 mm,中线左右2.0~2.1 mm,用颅骨钻打磨颅骨,钻出合适大小的骨窗。注射浓度为2 mg/mL的CTB555(Cholera Toxin Subunit B,Alexa FluroTM555 Conjugate,Invitrogen,USA)0.2 μL,注射速度控制在0.02 μL/min。光遗传的实验中,将CTB555 0.2 μL 和ChR2-EGFP(pAAV-CAMKⅡa-hChR2(H134R)-EGFP,滴度为(3~4)×1013V.G./mL,和元生物,上海)0.2 μL,混合成0.4 μL溶液同时注射到LGN。为了保证足够的注射的量和扩散面积,在相应位置上注射两个点。注射完毕后留针5~10 min。1~2 h后,待动物苏醒后,正常条件下饲养。
1.3 急性脑片制备
注射CTB555 1 周后或联合注射CTB555+ChR2-EGFP 3 周后的小鼠,异氟烷使之麻醉,断头,剪开颅骨后取出大脑,迅速放置到通有体积分数95%O2和5%CO2混合气、0 ℃的切片液,30 s 后转移至培养皿中修块,将脑片以横断面为底座,用胶水固定于振动切片机平台上,在通入体积分数95%O2和5%CO2混合气体,0 ℃的切片液中作视皮层的冠状切片,厚度300 μm。随后转移到通入95%O2和5%CO2混合气体、室温条件下的孵育液中,孵育1 h 后用于全细胞记录。
1.4 标记L6b 反馈投射到LGN 的神经细胞
用盛有包含biocytin(Sigma,USA)的电流钳电极液的玻璃电极,在视皮层的脑片上,选择被CTB555 标记的L6b 红色荧光细胞,钳住细胞5~10 min,载入biocytin。然后缓慢移出电极,保持细胞活性状态。每张脑片标记1~3 个神经细胞。后将脑片转移至40 g/L多聚甲醛溶液中固定,避光保存于4 ℃冰箱。24 h后,取出脑片盛放在12孔板的小孔里,用0.01 mol/L PBS 溶液漂洗3~5 次。再将脑片放入体积分数0.3% Triton-100 的PBS 中2 h。按1∶200稀释度加入Avidin 488(Streptavidin,Alexa Fluro 488 conjugate,Invitrogen corporation,USA),混匀后,用锡纸包裹,放置4 ℃冰箱过夜。后再用0.01 mol/L PBS 溶液漂洗干净,双光子显微镜扫描拍照,观察细胞的形态。
1.5 全细胞记录刺激诱发的AMPAR-EPSC 和NMDAR-EPSC
荧光显微镜下观察到由CTB555 逆行标记的L6b 的红色荧光细胞并采用电压钳电极液的方法钳住细胞。电刺激电极(同心圆金属电极,尖端150 μm,FHC,USA)放置在白质(MW)上,或用蓝光(波长470 nm,由Polygon 400E 激发,Mightex,Canada)直接刺激由ChR2-EGFP 标记的LGN 前馈投射得到VC 的神经纤维。记录由电刺激WM 引起的AMPAR-EPSC(amino-3-hydroxy-5-methylisoxazole-4-propionic acid receptor-excitatory postsynaptic current)和NMDAR-EPSC 或蓝光刺激引起的光诱发AMPA-EPSC。
1.6 数据统计与分析
实验采用AXON 公司的Clampex 软件记录电生理数据,数据在Clampfix 中打开并分析,Igor pro处理电生理trace,最后用GraphPad Prism 8 对数据进行汇总和方差分析。
2 结果
2.1 Orexin B 直接兴奋L6b 反馈投射至LGN 的神经元
为了研究L6b 中反馈投射到LGN 的神经细胞形态以及它们是否对Orexin B 有反应,我们将CTB555 注射到LGN 中(图1A)。CTB555 是常用的逆行标记物。微量注射CTB555 到LGN 后,可载入到L6b 反馈投射到LGN 的神经末梢中,经逆向运输到细胞体(图1B)。外膝体注射CTB555 小鼠7~10 d 后,红色荧光标记的细胞主要分布在视皮层的L6 中,表明标记了L6 的神经细胞(图1B2,B3)。用含有biocytin 的电极液,一共钳住了93 个L6b 中的红色荧光细胞。biocytin 标记的脑片经Avidin 488 标记后,在双光子显微镜下发现有86 个细胞为锥体神经元(图1C),仅有7 个细胞是多极神经元(图1D),没有标记到其它类型的神经细胞。这个结果表明:L6b 反馈传入外膝体的神经元主要是锥体神经元(图1E)。
随后,我们在普通灌流液(artificial cerebral spinal fluid,ACSF)中加入兴奋和抑制性突触受体的阻断剂[CNQX(6-cyano-7-nitroquinoxaline-2,3-dione)],20 μmol/L;APV[(2R)-amino-5-phosphonovaleric acid;AP5,(2R)-amino-5-phosphonopentanoate],100 μmol/L μmol/L;PTX(picrotoxin),50 μmol/L,All from Sigma,USA),阻断所有兴奋和抑制性的突触传递。在荧光显微镜下,用电流钳方法钳住由LGN逆行标记到L6b中带有红色荧光的细胞(图1、2),记录细胞的膜电位。将细胞钳制在该细胞的阈电位水平,大约在-45~-40 mV 之间,让细胞产生几个稀疏的动作电位。待细胞稳定下来后(约3 min),然后换成含有100 nmol/L Orexin B(Tocris Bioscience,UK)的ACSF(其中含有兴奋性和抑制性突触受体的阻断剂),灌流3 min 后,再用普通ACSF 洗脱来观察这些由LGN逆行标记到L6b 细胞是否被Orexin B 的激动剂所兴奋,结果发现:在一共研究的30 个L6b 细胞的红色荧光细胞有19 个细胞(占63.3%)随着Orexin B 混合灌流液的加入而逐渐爆发大量成串的动作电位,并且普通ACSF 的洗涤后,持续一段时间,最后逐渐消失(图1F)。为此,将这类对Orexin B有直接反应的细胞定义为Orexin 敏感神经元。这些结果提示,在视皮层L6b 反馈投射到LGN 的神经元是锥体神经元,而且有一半以上的细胞是Orexin 敏感神经元。
2.2 Orexin B 增强LGN 投射 纤 维到L6b 中Orexin B 敏感神经元之间的突触传递
上述结果已经证明了视皮层L6b 反馈投射到LGN的锥体神经元是Orexin敏感神经元。为了研究Orexin B 能否调控LGN 投射纤维到L6b 中Orexin敏感神经元之间的突触传递,我们用电压钳的方法钳住由CTB555 标记的L6b 红色荧光细胞(图1B3),将同心刺激电极放置于白质,激活来自LGN的前馈传入纤维,诱发L6b 中Orexin 敏感神经元的突触后电流(evoked excitatory post-synaptic currents,eEPSC)。膜电位钳制在-70 mV,待细胞稳定后,先调整刺激强度,一般采用不产生动作电位的最大EPSC 振幅的1/3 大小的刺激强度。记录基线10 min后,加入100 nmol/L Orexin B 灌流10 min,后切换正常的孵育液进行药物洗脱。实验全程刺激强度不变并维持系列电阻基本稳定(<25 MOhm)。结果显示:Orexin B 增强了LGN 投射纤维到L6b中Orexin 敏感神经元之间的突触传递(图2A)。一共记录的20 个细胞中,在Orexin B 洗脱后20~25 min 后,EPSC的振幅是基线的(128.52±5.01)%,图2B显示一个代表性的eEPSC 记录,可明显观察到加入Orexin B 后EPSC 振幅的增大。通过比较配对脉冲比率(paired-pulse ratio,PPR)的值,加入Orexin 之前PPR为1.19±0.03,加入Orexin B并洗脱后20~25 min 时PPR 为1.14±0.03,没有显示出差异性(n=20,pairedttest:t=0.28,P=0.36,图2C),说明Orexin B 增强LGN 投射纤维到L6b Orexin敏感神经元之间的突触传递作用是突触后效应。且这种增强效应被OX2R 的阻断剂TCS OX2 29(10 μmol/L)所阻断[Ctr:(132.31±7.81)%,n=10 cells from 5 mice;TCS OX2 29:(108.81±3.42)%,n=17 cells from 6 mice;unpairedttest,t=18.59,P=0.016,图2 D],说明Orexin B 对LGN 到L6b 的突触传递的增强作用是由OX2 受体所介导。

图1 Orexin B 直接兴奋L6b 反馈投射至LGN 的神经元Fig.1 Orexin B directly activated neurons feedback projection to LGN in L6b labeled by CTB555
2.3 Orexin B 通过NMDA 受体增强LGN 投射纤维到L6b 中锥体细胞的突触传递
上述结果已经证明Orexin B 增强LGN 投射纤维到L6b Orexin 敏感神经元之间的突触传递。为了区分Orexin B 的增强作用是通过直接增强AMPA 受体介导的电流还是增强NMDAR 介导的电流,我们在ACSF 中,加入AMPA 受体的阻断剂(6-cyano-7-nitroquinoxaline-2,3-dione,CNQX)20 μmol/L 和GABAA 受体的阻断剂(picrotoxin)50 μmol/L,并将膜电位钳在+40 mV,从而分离出NMDAR 电流,同心圆刺激电极置于白质,激活来自LGN 的前馈传入纤维,记录视皮层L6b 中的红色荧光细胞(图1B3)上的NMDAR-EPSC。结果显示,Orexin B 增强L6b 神红色荧光细胞上的NMDAR-EPSC,并且这种增强效应呈现浓度依赖性[加入Orexin B并洗脱后20~25 min,100 nmol/L:(110.23±2.31)%,n=9 cells from 5 mice;200 nmol/L:(125.12±4.81)%,n=12 cells from 7 mice;unpairedttest,t=18.83,P=0.026,图3A]。在TCS OX2 29(OX2R 的阻断剂)存在的情况下,Orexin B 增强NMDAR-EPSCs 的效应消失[加入Orexin B 并洗脱后20~25 min,Ctr:(125.1±3.7)%,n=7 cells from 3 mice;TCS-OX2-29:(105.14±5.11)%,n=12 cells from 5 mice;图3B;unpairedttest,t=29.79,P=0.005]。APV(NMDAR 阻断剂)100 μmol/L 存在下,记录eEPSC(方法同图2A),Orexin B 对LGN到L6b 的突触传递的增强作用消失[加入Orexin B并洗脱后20~25 min,Ctr:(128.73 ±5.74)%;n=14 cells from 6 mice,APV:(106.72 ±4.23)%,n=15 cells from 7 mice,unpairedttest,t=52.56,P=0.006,图3C]。以上实验结果说明:Orexin B 增强LGN 投射纤维到L6b Orexin 敏感神经元之间的突触传递是通过增强NMDAR 受体电流,间接增强AMPAR 电流,从而增强了突触传递。
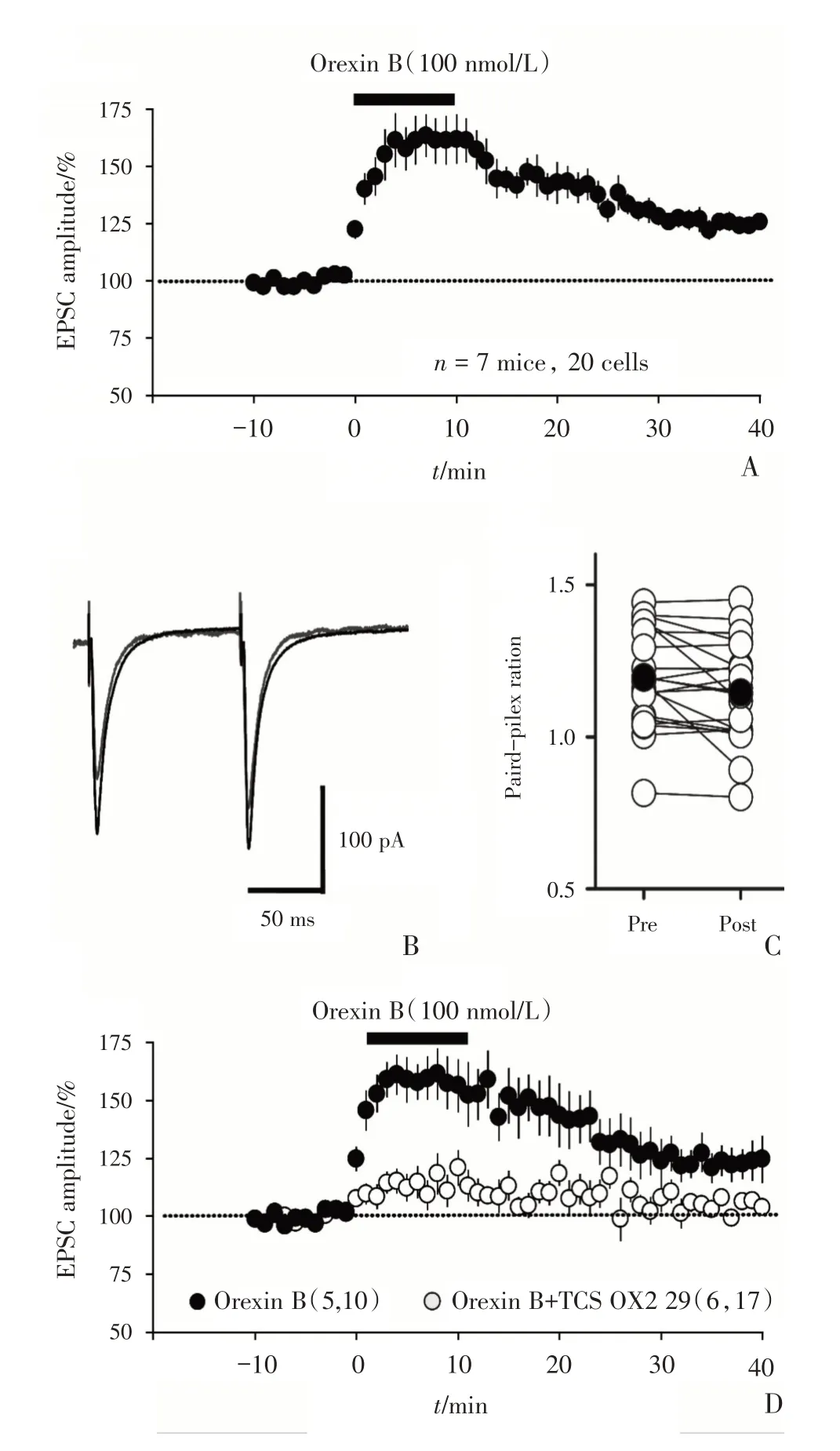
图2 Orexin B 通过OX2R 增强LGN 前馈投射到L6b Orexin 敏感神经元之间的突触传递Fig.2 Orexin B enhanced the synaptic transmission between feed-forward projection from the lateral geniculate nucleus(LGN)to orexin-sensitive neurons in L6b via OX2R
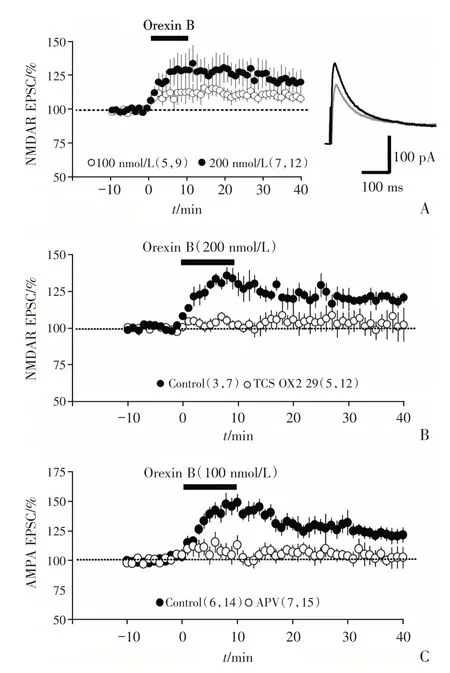
图3 Orexin B 增强LGN 前馈投射到L6b Orexin 敏感神经元上的NMDAR-EPSC 振幅Fig.3 Orexin B enhanced the amplitude of NMDAREPSC in Orexin B-sensitive neurons in L6b labeled by CTB555 evoked from LGN
2.4 Orexin B 同样增强光激活LGN 纤维投射至L6b 锥体细胞的突触传递
如前所述,电刺激视皮层白质,记录L6b 中Orexin 敏感神经元上的eEPSC,Orexin B 可以增大这种eEPSC 的振幅。电刺激视皮层白质可以激活来自LGN 投射至L6b 的纤维,但也可能激活了来自皮层内的皮层-皮层细胞(CC cells)的神经纤维以及来自另一侧皮层经胼胝体传入到另一侧皮层的神经纤维,为了直接刺激仅仅源自LGN到皮层的前馈投射纤维,我们选用P25 左右的C57BL/6J 小鼠,将CTB555 和ChR2-EGFP 混合后,微量注射到LGN(图4A),3~4 周后灌流取脑,切片。在荧光显微镜下,可以观察到由ChR2-EGFP标记的LGN 投射到VC 的绿色神经纤维(图4B1)。源自LGN 的投射纤维在视皮层上广泛分布,但主要分布在L4 和L2/3(图4B2)。L5/6 也有分布(图4B3),同时L6b 中可观察到带有红色荧光标记的细胞(图4B2、B3)。电压钳模式下钳住红色荧光标记的细胞(如前述),用蓝色LED 刺激由ChR2-EGFP 标记的LGN 投射到VC 的绿色神经纤维,记录由光刺激引起的pEPSC(photo-induced excitatory postsynaptic current)。每15 s 给予一对蓝光脉冲刺激,脉冲时间间隔也是0.1 s(图4D)。细胞钳制在-70 mV,细胞稳定后,调整光刺激强度和光刺激时长,一般采用不产生动作电位的最大pEPSC的1/3~1/2对应的刺激大小。记录10 min的基线,随后加入100 nmol/L Orexin B 混合循环液,灌流10 min后用普通ACSF 洗去,整个过程持续记录40 min。实验过程中不改变光刺激强度和光刺激时长,并维持系列电阻基本稳定(<25 MOhm)。结果显示:随着Orexin B 的加入,pEPSC 振幅逐渐增大(图4C、D),加药10 min 后,洗脱之前,pEPSC振幅达到最大值[(136.74±7.81)%,n=11 cells,6 mice]。随着洗脱,pEPSC 逐渐下降,最后趋于平缓(图4C)。说明Orexin B 增强了源自LGN 投射纤维到L6b Orexin 敏感神经元之间的突触传递功能。且这种增强效应可被OX2R 的阻断剂TCS OX2 29(10 μmol/L)所阻断[加入Orexin B 并洗脱后20~25 min,Ctr:(123.82±3.78)%,n=11 cells,from 5 mice;TCS OX2 29:(105.25±2.41)%,n=9 cells from 4 mice;unpairedttest,t=42.86,P=0.031,图4C、D],提示Orexin B 对源自LGN 到L6b的突触传递的增强作用是由OX2 受体所介导。通过对比配对脉冲比率(paired-pulse ratio,PPR)的值,Orexin B 加药之前的PPR 为0.63±0.08,加入Orexin B 并洗脱后20~25 min PPR 为0.62±0.07,加药前后PPR 无显著变化(n=11 cells,pairedttest,t=3.16,P=0.57,图4E)。这些结果显示:源自LGN 至L6b Orexin 敏感细胞的前馈突触传递中,Orexin B 可以增加L6b 中Orexin 敏感神经元的突触后电流,与电刺激的结果相对一致,即Orexin B可以增强LGN 投射到L6b 锥体神经元之间的突触传递,且这种变化为突触后效应。
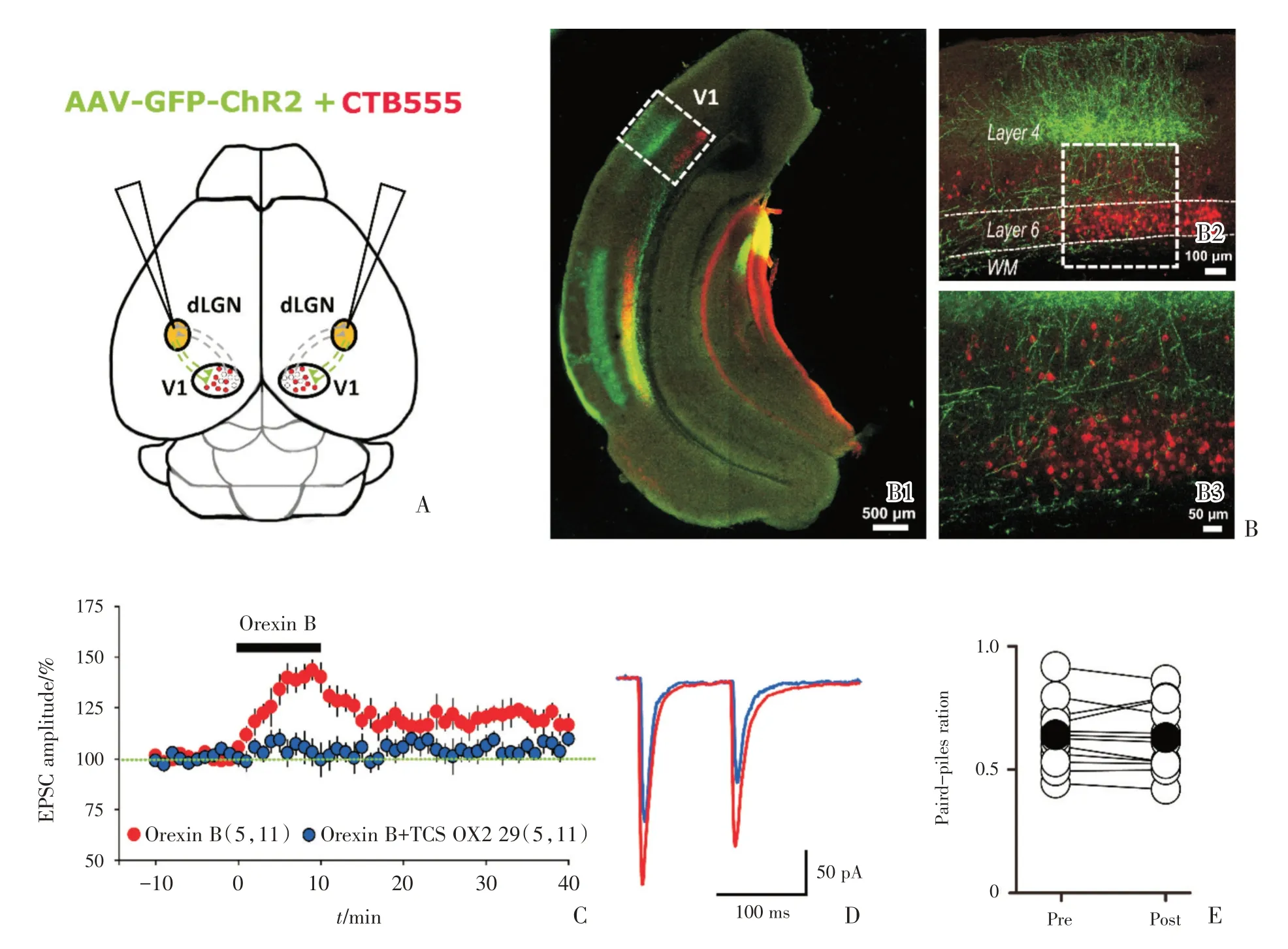
图4 Orexin B 增强光激活源自LGN 的投射纤维到L6b Orexin 敏感神经元之间的突触传递Fig.4 Orexin B enhanced the synaptic transmission between feed-forward projection from the LGN to orexin-sensitive neurons in L6b by light stimulation
3 讨论
前人报道在听皮层、体感皮层以及视皮层上的第6b 层(L6b)的某些神经元对OX 有反应[8],但没有明确对OX 有反应的具体细胞类型。由于皮层的L6 中有不同类型的神经元和不同神经连接,可分为锥体细胞、多极细胞、倒置朝向细胞、水平朝向细胞和不定向细胞[9-10]。本研究通过微量注射CTB555 逆行标记L6b 中反馈投射到LGN 的神经细胞,明确了在小鼠的视皮层L6b 中,反馈投射至LGN 的神经细胞为锥体神经元,而不是其他类型的神经元,且这些神经细胞大多数对Orexin B受体激动剂敏感,表现为Orexin 激动剂Orexin B结合OX2 受体引起这些锥体神经元直接产生成串的动作电位,因此,L6b 中的锥体细胞的细胞膜上富含有OX2 受体。之前大多数研究LGN-VC 的突触传递多采用电刺激视皮层白质,记录L6 细胞的eEPSC[11-12]。由于电刺激不可避免地激活了其它来源的神经纤维,不能刺激纯粹源自LGN 的传入纤维。为了明确刺激仅仅源自LGN 到皮层的前馈投射纤维源自LGN 的神经纤维,本研究采用ChR2-EGFP 病毒感染LGN 细胞,标记出投射到L6b 的锥体神经元源自LGN 神经纤维,避免了电刺激皮层白质的诸多问题。源自LGN 神经纤维构成视觉信息的前馈投射,将视觉信息中继到视皮层,而Orexin B 可以增强源自LGN 的神经纤维投射到L6b 的锥体神经元之间的突触传递,提高LGN 向视皮层传递视觉信息的效能,而L6 细胞的活性增强可以调控视皮层表层L4 和L2/3 细胞的兴奋性,增强L4 和L2/3 细胞的视觉刺激反应,但不改变视觉的方位选择性[13]。由于OX 的主要作用是维持动物的觉醒状态[1-2],据此推测动物在清醒的状态,Orexin 系统可以参与放大外界视觉信息传递到视皮层的过程中,可使动物在相对于睡眠的清醒、警觉的状态下更为有效的感知外界视觉信息。
已有很多研究证明,Orexin 对中枢神经系统的神经元具有兴奋作用。由于OX 的受体亚型在中枢神经中的分布各不相同。在皮质下觉醒系统中,除了蓝斑神经元以OX1R 为主,OX2R 占的比例较大[4],在大脑皮层中主要以OX2R 的分布为主。因此该项研究中,我们应用了OX2R 阻断剂TCS-OX2-29。单独使用OX2R 阻断剂就可以阻断Orexin B 对LGN 的投射纤维到L6b Orexin 敏感神经元之间的突触传递作用,进一步证明在视皮层Orexin 敏感神经元上可能主要分布OX2R。
Orexin 作为兴奋性神经肽,能对受体神经元产生一个缓慢且长久的去极化,Orexin 兴奋神经元以及促进突触传递的作用的机制如下[5,14-17]。①降低钾通道的通透性。静息状态时,细胞主要是钾离子通透,当Orexin 作用于Orexin 受体时,通过降低钾通道电导的作用,促使细胞膜对钾离子的通透性减弱,钾离子外流受阻,上抬膜电位,至阈电位时产生动作电位。②钠钙离子交换体(Na+/Ca2+exchanger,NCX)的激活。Orexin 受体是G 蛋白偶联受体。Orexin 与细胞上的受体结合后激活磷脂酶C(phosholipase C,PLC)通路,作用于内质网中的钙离子释放,胞内钙离子浓度升高。NCX 按照3∶1 的比例将钠离子转入细胞,钙离子转出细胞[15],导致细胞兴奋。③非选择性阳离子通道(non-selective cation channels,NSCC)的激活。NSCC 属于非电压依赖性通道,NSCC 可以被钙离子激活。这些受体显著影响Ca2+信号,通过经典的PLC-Ca2+释放途径,或其他多种途径介导Ca2+内流。最后导致突触可塑性的改变。该项研究中,Orexin B 直接兴奋了L6b 上的Orexin 敏感神经元,可能机制是通过上述机制中的①和②途径。Orexin 增强源自LGN 的神经纤维投射到L6b的锥体神经元之间的突触传递,其机理可能通过上述机理中的②和③,Orexin 可能与细胞上的受体结合后激活磷脂酶C(phosholipase C,PLC)通路,升高胞内钙离子浓度升高,引起L6b Orexin 敏感细胞中的AMPA 受体向突触后膜移动,增大eEPSC[15-16]。
目前尚不清楚对于NMDA 或AMPA 受体的调节作用是否是Orexin 的一种普遍功能,有报道OX增强了NMDAR 驱动的VTA 神经元胞体和树突上的多巴胺的释放[16]。本研究中,在NMDAR 阻断剂APV 存在的情况下,Orexin B 增强来自LGN 到L6b Orexin 敏感细胞之间突触传递效应消失,表明Orexin B 的增强效应是由NMDA 受体介导。Orexin 直接增强NMDAR 功能的具体机理需进一步的探讨。
综上所述,本研究证明了从L6b 反馈投射至LGN 的细胞主要为锥体神经元,并且这些神经元都是Orexin 敏感神经元。电刺激或光刺激从LGN前馈投射至L6b 的兴奋性神经纤维,Orexin 增强源自LGN 的神经纤维投射到L6b 的锥体神经元之间的突触传递,这种效应主要与L6b 中含有OX2 受体的锥体神经元上的NMDA 受体功能增强有关。
